
Genetic Diversity and Maternal Phylogenetic Relationships among Populations and Strains of Arabian Show Horses
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Population Samples
2.2. DNA Extraction and Whole D-Loop Sequencing
2.3. Data Analyses
3. Results
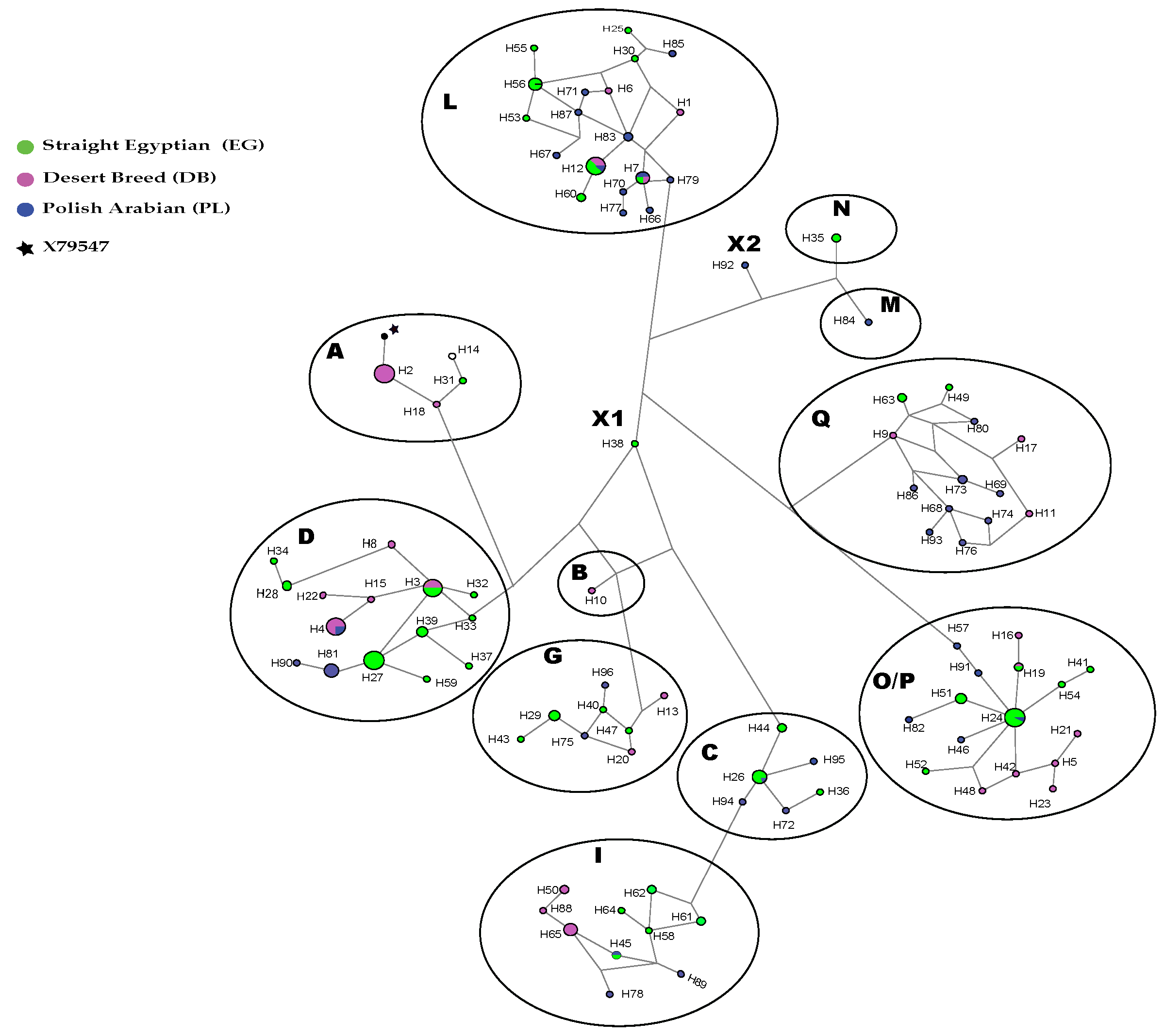
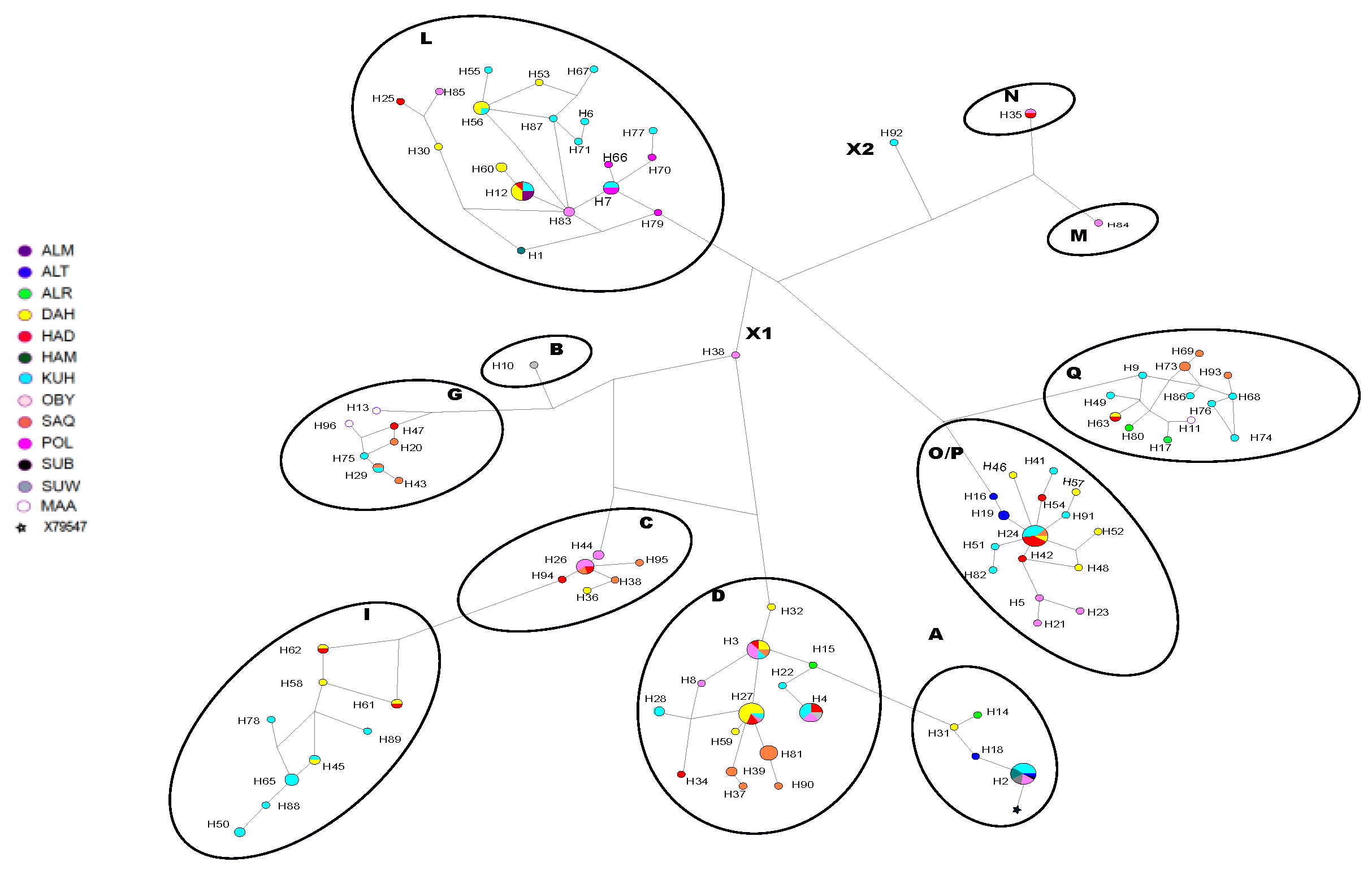
3.1. Haplotypes and Haplogroups
3.2. DNA Sequence Polymorphism and Tests of Neutrality
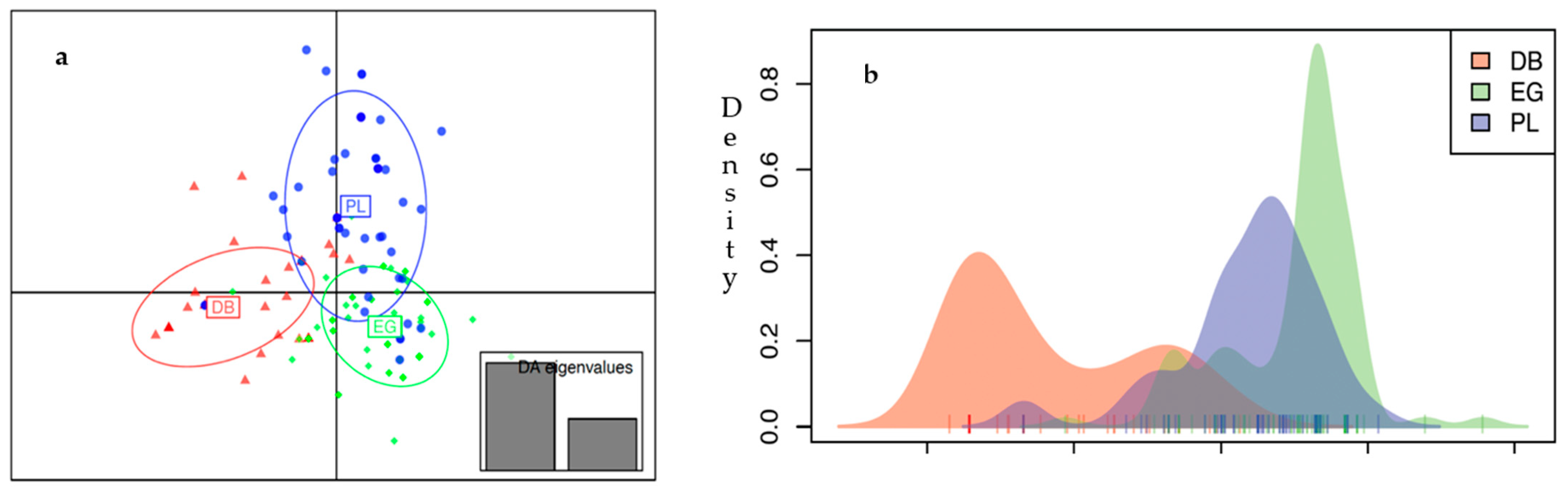
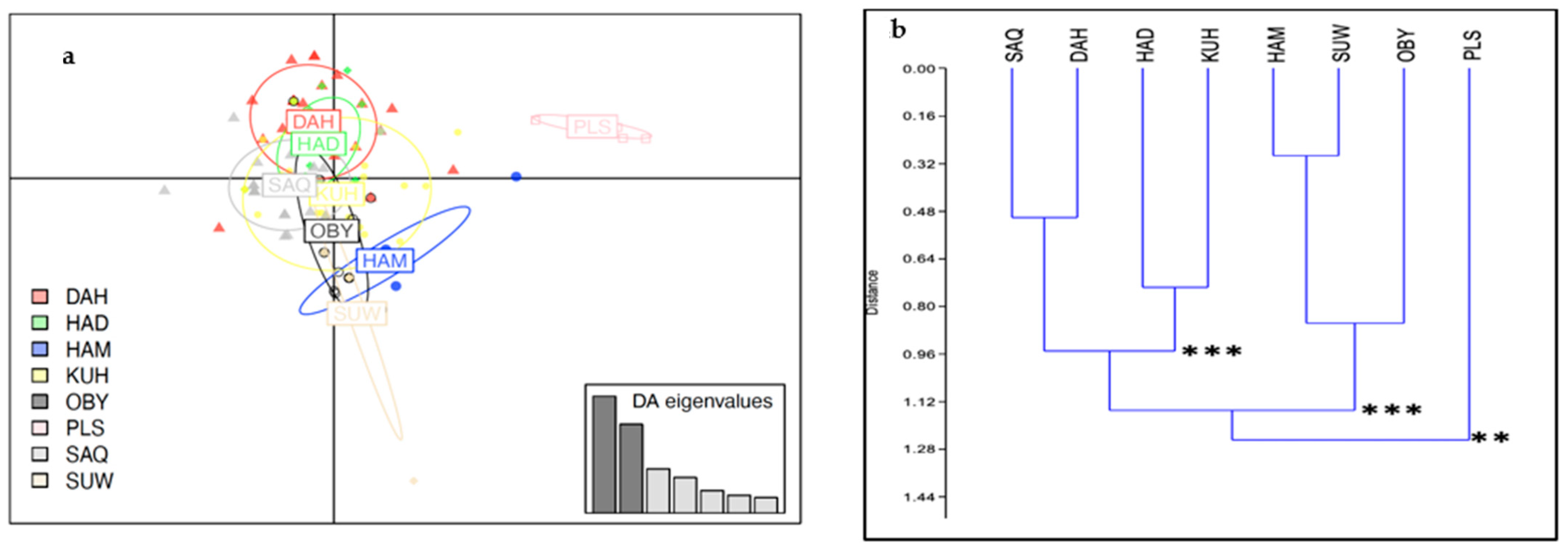
3.3. Genetic Differentiation between Populations and between Strains
3.3.1. Population-Level
3.3.2. Strain Level
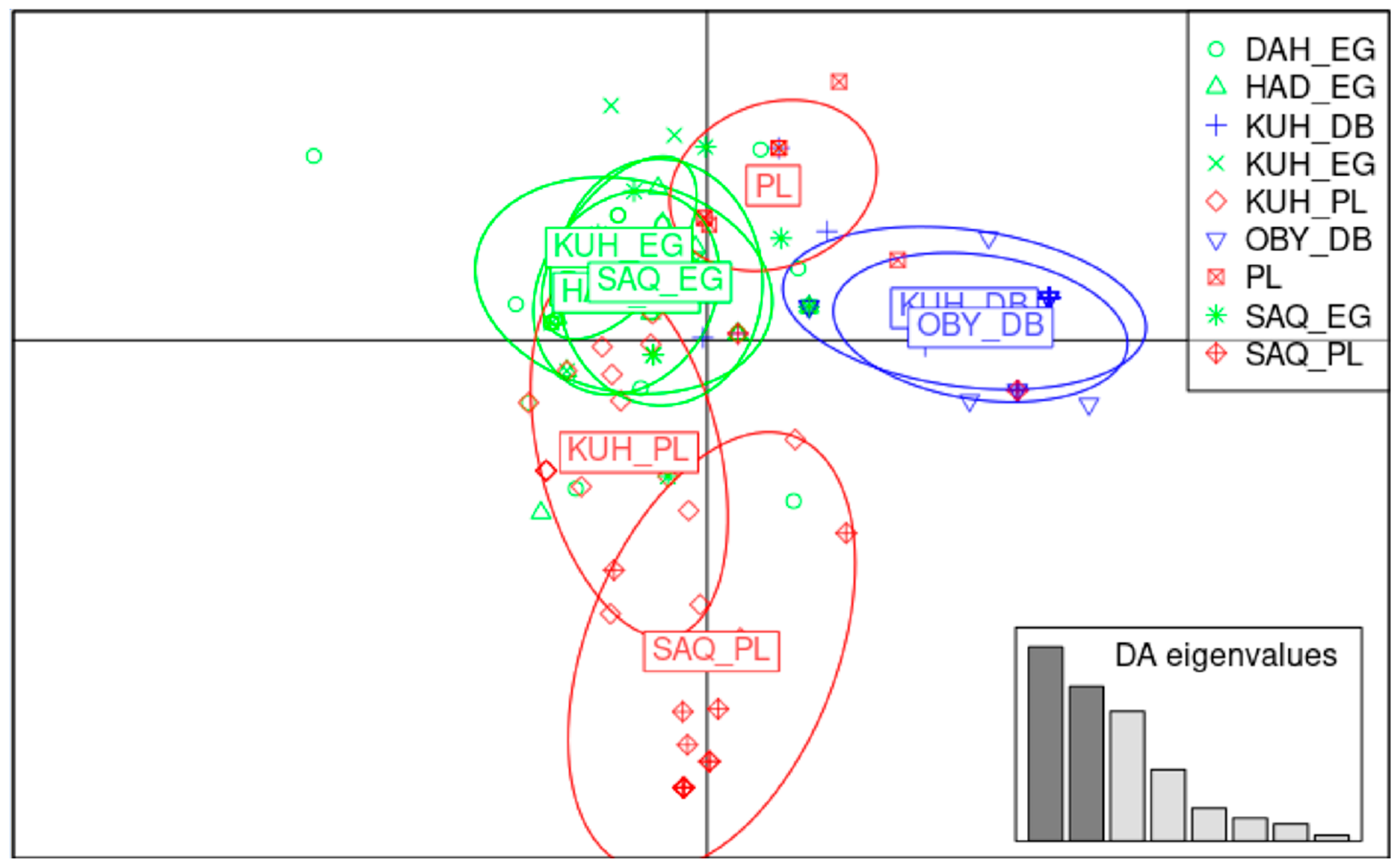
3.3.3. Population and Strain Level Combination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagel, H. The Arabian Horse: Nature’s Creation and the Art of Breeding, 1st ed.; Nawal Media: Spoleto, Italy, 2013. [Google Scholar]
- Qurachi, A. Arab Horse Traits Law: An Original Study between Abu Ubaidah and Al-Asma’i; Shabanat Press: Riyadh, Saudi Arabia, 2012. [Google Scholar]
- Forbis, J.; Gulsan, S. The Abbas Pasha Manuscript and Horsemen of Arabian and Egypt during the Time of Abbas Pasha 1800–1860; Ansata Publication: Mena, AR, USA, 1993. [Google Scholar]
- Al-Dahdah, E. The State of Arabian Breeding in its Area of Origin; Al Khaima, the Institute’s Magazine: New York, NY, USA, 2005; Volume 2. [Google Scholar]
- Forbis, J. The Classic Arabian Horse; WW Norton & Company: New York, NY, USA, 1976. [Google Scholar]
- Machmoum, M.; Mezrour, L.; Boujenane, I.; Piro, M. Morphologie comparative des chevaux arabes de lignées égyptienne, polonaise et Desert Breed. Rev. Marocaine Sci. Agron. Vét. 2019, 7, 58–166. [Google Scholar]
- Cosgrove, E.J.; Sadeghi, R.; Schlamp, F.; Holl, H.M.; Moradi-Shahrbabak, M.; Miraei-Ashtiani, S.R.; Abdalla, S.; Shykind, B.; Troedsson, M.; Stefaniuk-Szmukier, M.; et al. Genome Diversity and the Origin of the Arabian Horse. Sci. Rep. 2020, 10, 9702. [Google Scholar] [CrossRef] [PubMed]
- Skorkowski, E. Polish Genealogical Tables of Pure-Bred Arab Horse; Państwowe Wydawnictwo Rolnicze i Leśne: Krakow, Poland, 1960. [Google Scholar]
- Głazewska, I.; Jezierski, T. Pedigree analysis of Polish Arabian horses based on founder contribution. Livest. Prod. Sci. 2004, 90, 293–298. [Google Scholar] [CrossRef]
- Machmoum, M.; Boujenane, I.; Azelhak, R.; Badaoui, B.; Petit, D.; Piro, M. Genetic Diversity and Population Structure of Arabian Horse Populations Using Microsatellite Markers. J. Equine Vet. Sci. 2020, 93, 103200. [Google Scholar] [CrossRef]
- Khanshour, A.; Conant, E.; Juras, R.; Cothran, E.G. Microsatellite analysis of genetic diversity and population structure of Arabian horse populations. J. Hered. 2013, 104, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Khanshour, A.M.; Cothran, E.G. Maternal phylogenetic relationships and genetic variation among Arabian horse populations using whole mitochondrial DNA D-loop sequencing. BMC Genet. 2013, 14, 83. [Google Scholar] [CrossRef] [Green Version]
- Almarzook, S.; Reissmann, M.; Arends, D.; Brockmann, G.A. Genetic diversity of Syrian Arabian horses. Anim. Genet. 2017, 48, 486–489. [Google Scholar] [CrossRef]
- Sadeghi, R.; Moradi-Shahrbabak, M.; Miraei Ashtiani, S.R.; Schlamp, F.; Cosgrove, E.J.; Antczak, D.F. Genetic diversity of Persian arabian horses and their relationship to other native iranian horse breeds. J. Hered. 2019, 110, 173–182. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Xu, X.; Gullberg, A.; Arnason, U. The complete mitochondrial DNA (mtDNA) of the donkey and mtDNA comparisons among four closely related mammalian species-pairs. J. Mol. Evol. 1996, 43, 438–446. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.-X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genet. Soc. Am. Stat. 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.L.; Clark, A.G.; Clark, A.G. Principles of Population Genetics, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 1997. [Google Scholar]
- Wright, S. Evolution and the Genetics of Populations. Variability within and among Natural Populations; Chicago University Press: Chicago, IL, USA, 1978. [Google Scholar]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Kashani, B.H.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef] [Green Version]
- Vilà, C.; Leonard, J.A.; Gotherstrom, A.; Marklund, S.; Sandberg, K.; Lidén, K.; Wayne, R.K.; Ellegren, H. Widespread origins of domestic horse lineages. Science 2001, 291, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: PAleontological STatistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. Genetics 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Hudson, W. Whole-loop mitochondrial DNA D-loop sequence variability in Egyptian Arabian equine matrilines. PLoS ONE 2017, 12, e0184309. [Google Scholar] [CrossRef] [Green Version]
- Almarzook, S.; Reissmann, M.; Brockmann, G.A. Diversity of mitochondrial DNA in three Arabian horse strains. J. Appl. Genet. 2017, 58, 273–276. [Google Scholar] [CrossRef]
- Głazewska, I.; Wysocka, A.; Gralak, B.; Prus, R.; Sell, J. A new view on dam lines in Polish Arabian horses based on mtDNA analysis. Genet. Sel. Evol. 2007, 39, 609–619. [Google Scholar] [CrossRef]
- Guastella, A.M.; Zuccaro, A.; Criscione, A.; Marletta, D.; Bordonaro, S. Genetic analysis of Sicilian autochthonous horse breeds using nuclear and mitochondrial DNA markers. J. Hered. 2011, 102, 753–758. [Google Scholar] [CrossRef] [Green Version]
- Bowling, A.T.; Del Valle, A.; Bowling, M. A pedigree-based study of mitochondrial D-loop DNA sequence variation among Arabian horses. Anim. Genet. 2000, 31, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jansen, T.; Forster, P.; Levine, M.A.; Oelke, H.; Hurles, M.; Renfrew, C.; Weber, J.; Olek, K. Mitochondrial DNA and the origins of the domestic horse. Proc. Natl. Acad. Sci. USA 2002, 99, 10905–10910. [Google Scholar] [CrossRef] [Green Version]
- Aberle, K.S.; Hamann, H.; Drögemüller, C.; Distl, O. Phylogenetic relationships of German heavy draught horse breeds inferred from mitochondrial DNA D-loop variation. J. Anim. Breed. Genet. 2007, 24, 94–100. [Google Scholar] [CrossRef]
- Cieslak, M.; Pruvost, M.; Benecke, N.; Hofreiter, M.; Morales, A.; Reissmann, M.; Ludwig, A. Origin and history of mitochondrial DNA lineages in domestic horses. PLoS ONE 2010, 5, e15311. [Google Scholar] [CrossRef] [Green Version]
- Zaher, A. Arabian Horse Breeding and the Arabians of America, 2nd ed.; Cairo University Press: Cairo, Egypt, 1961. [Google Scholar]
- Lizet, B. Le cheval arabe du Nejd et le système des races orientales dans le manuscrit de Wenceslas Severyn Rzewuski. Anthropozoologica 2004, 39, 79–97. [Google Scholar]
- Remer, V.; Bozlak, E.; Felkel, S.; Radovic, L.; Rigler, D.; Grilz-Seger, G.; Stefaniuk-Szmukier, M.; Bugno-Poniewierska, M.; Brooks, S.; Miller, D.C.; et al. Y-Chromosomal Insights into Breeding History and Sire Line Genealogies of Arabian Horses. Genes 2022, 13, 229. [Google Scholar] [CrossRef]
- Raswan, C.R.; Wenzler, G.; Seydel, H. Der Araber und Sein Pferd; Konrad Wittwer Verlages: Stuttgart, Germany, 1981. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop. | N | NHap | NPs | HD (SD) | Pi (SD) | K | Fu Li’s D * | Fu Li’s F * | Fu’s Fs | Th |
|---|---|---|---|---|---|---|---|---|---|---|
| DB | 54 | 23 | 44 | 0.822 (0.050) | 0.009 (0.010) | 8.372 | 0.167 | −0.071 | −3.011 ** | −0.450 |
| EG | 106 | 45 | 55 | 0.925 (0.018) | 0.009 (0.011) | 11.553 | 0.758 | 0.688 | −10.580 *** | 0.309 |
| PL | 51 | 39 | 52 | 0.985 (0.009) | 0.011 (0.013) | 12.940 | 0.641 | 0.665 | −16.969 *** | 0.412 |
| All | 211 | 96 a | 69 | 0.964 (0.006) | 0.013 (0.013) | 11.780 | 0.150 | 0.118 | −61.499 *** | −0.171 |
| Strains | N | NHap | NPS | HD (SD) | Pi (SD) | K |
|---|---|---|---|---|---|---|
| Dahman (DAH) | 38 | 23 | 38 | 0.936 (0.029) | 0.012 (0.002) | 11.030 |
| Hadban (HAD) | 24 | 14 | 44 | 0.833 (0.077) | 0.011 (0.002) | 10.395 |
| Hamdani (HAM) | 9 | 5 | 29 | 0.806 (0.120) | 0.011 (0.018) | 9.778 |
| Kuhailan (KUH) | 59 | 36 | 50 | 0.948 (0.018) | 0.013 (0.002) | 12.252 |
| Obeyan (OBY) | 19 | 11 | 22 | 0.924 (0.037) | 0.007 (0.001) | 7.053 |
| Saqlawi (SAQ) | 29 | 21 | 51 | 0.963 (0.023) | 0.013 (0.013) | 12.986 |
| Suwaity (SUW) | 5 | 3 | 14 | 0.700 (0.218) | 0.007 (0.002) | 6.200 |
| All | 183 | 86 * | 67 | 0.961 (0.007) | 0.013 (0.013) | 11.820 |
| DB | EG | PL | |
|---|---|---|---|
| DB | - | - | - |
| EG | 0.013 | - | - |
| PL | 0.015 | 0.005 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machmoum, M.; Badaoui, B.; Petit, D.; Germot, A.; El Alaoui, M.A.; Boujenane, I.; Piro, M. Genetic Diversity and Maternal Phylogenetic Relationships among Populations and Strains of Arabian Show Horses. Animals 2023, 13, 2021. https://doi.org/10.3390/ani13122021
Machmoum M, Badaoui B, Petit D, Germot A, El Alaoui MA, Boujenane I, Piro M. Genetic Diversity and Maternal Phylogenetic Relationships among Populations and Strains of Arabian Show Horses. Animals. 2023; 13(12):2021. https://doi.org/10.3390/ani13122021
Chicago/Turabian StyleMachmoum, Mohamed, Bouabid Badaoui, Daniel Petit, Agnès Germot, Moulay Abdelaziz El Alaoui, Ismaïl Boujenane, and Mohammed Piro. 2023. "Genetic Diversity and Maternal Phylogenetic Relationships among Populations and Strains of Arabian Show Horses" Animals 13, no. 12: 2021. https://doi.org/10.3390/ani13122021
APA StyleMachmoum, M., Badaoui, B., Petit, D., Germot, A., El Alaoui, M. A., Boujenane, I., & Piro, M. (2023). Genetic Diversity and Maternal Phylogenetic Relationships among Populations and Strains of Arabian Show Horses. Animals, 13(12), 2021. https://doi.org/10.3390/ani13122021