
Stability of Bacillus and Enterococcus faecium 669 Probiotic Strains When Added to Different Feed Matrices Used in Dairy Production

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiments
2.1.1. Exp. 1—Pellet Trial with Bacillus spp.
2.1.2. Exp. 2—Mineral–Vitamin Premix Trial with Bacillus spp.
2.1.3. Exp. 3—Milk Replacer Trial with Bacillus spp.
2.1.4. Exp. 4—Mineral–Vitamin Premix Trial with E. faecium 669
2.1.5. Exp. 5—Milk Powder Trial with E. faecium 669
2.1.6. Exp. 6—Milk Replacer Trial with E. faecium 669
2.2. Counts Methodologies
2.2.1. Bacillus Spore
2.2.2. Enterococcus faecium 669
2.3. Statistical Analysis
3. Results
3.1. Exp. 1
3.2. Exp. 2
3.3. Exp. 3
3.4. Exp. 4
3.5. Exp. 5
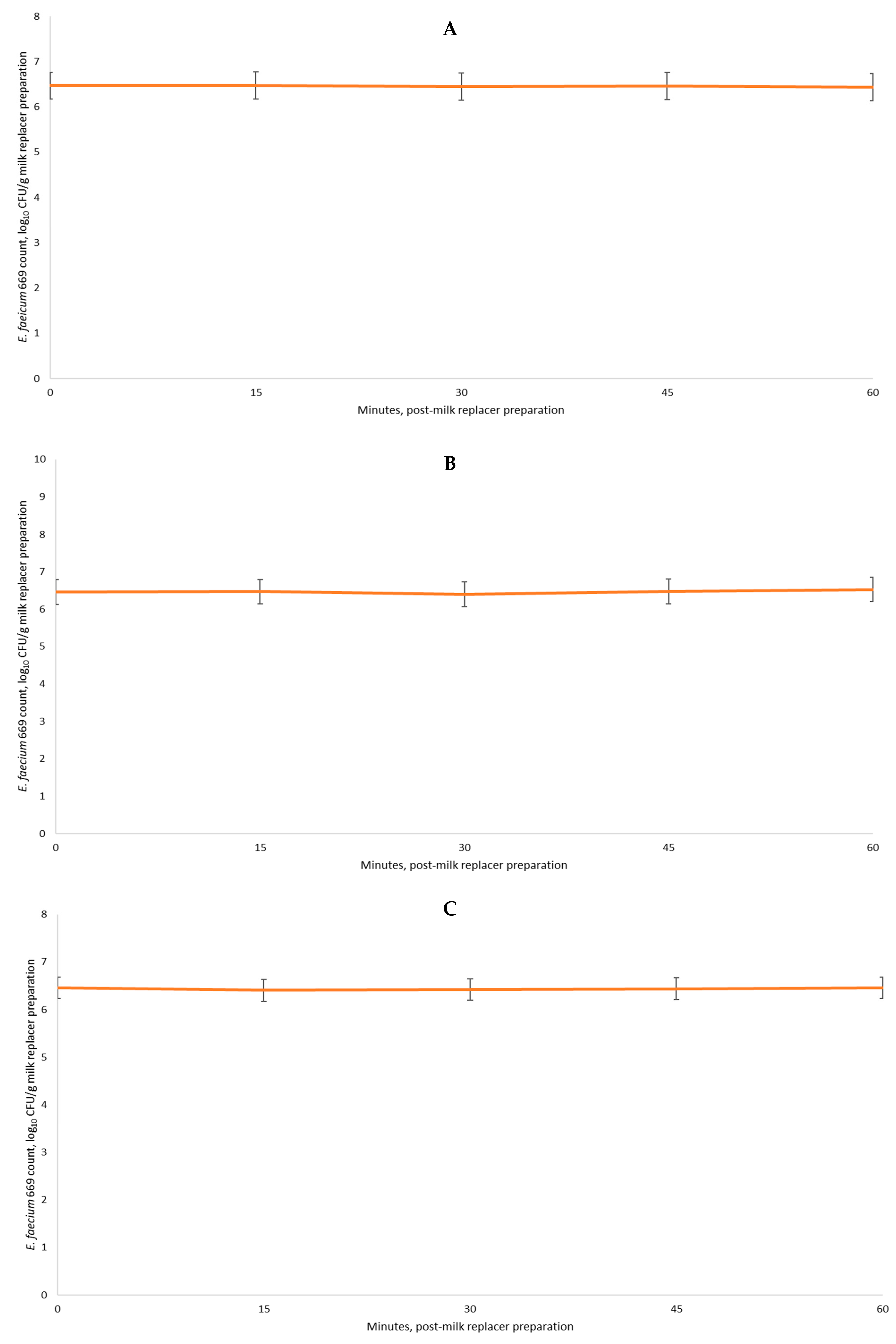
3.6. Exp. 6
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agricultural Organization of the United Nations and World Health Organization. Probiotics in Food: Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic acid Bacteria. 2001. Available online: https://www.fao.org/3/a0512e/a0512e.pdf (accessed on 4 April 2023).
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Cull, C.; Singu, V.K.; Cull, B.J.; Lechtenberg, K.F.; Amachawadi, R.G.; Schutz, J.S.; Bryan, K.A. Efficacy of Lactobacillus animalis and Propionibacterium freudenreichii-based feed additives in reducing Salmonella-associated health and performance effects in commercial beef calves. Antibiotics 2022, 11, 1328. [Google Scholar] [CrossRef] [PubMed]
- Cull, C.; Singu, V.K.; Cull, B.J.; Lechtenberg, K.F.; Amachawadi, R.G.; Schutz, J.S.; Bryan, K.A. Efficacy of two probiotic products fed daily to reduce Clostridium perfringens-based adverse health and performance effects in dairy calves. Antibiotics 2022, 11, 1513. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Beauchemin, K.A.; Alazzeh, A.Y.; Baah, J.; Teather, R.M.; Stanford, K. Review: The use of direct fed microbials to mitigate pathogens and enhance production in cattle. Can. J. Anim. Sci. 2011, 91, 193–211. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Harper, K.; Queiroz, O.; Copani, G.; Cappellozza, B.I. Effects of a Bacillus-based direct-fed microbial on in vitro nutrient digestibility of forage and high-starch concentrate substrates. Transl. Anim. Sci. 2022, 6, txac067. [Google Scholar] [CrossRef]
- Dias, B.G.C.; Santos, F.A.P.; Meschiatti, M.; Brixner, B.M.; Almeida, A.A.; Queiroz, O.; Cappellozza, B.I. Effects of feeding different probiotic types on metabolic, performance, and carcass responses of Bos indicus feedlot cattle offered a high-concentrate diet. J. Anim. Sci. 2022, 100, skac289. [Google Scholar] [CrossRef]
- Cappellozza, B.I.; Copani, G.; Boll, E.J.; Queiroz, O. Supplementation of the direct-fed microbial Enterococcus faecium 669 impacts performance of pre-weaning dairy calves. JDS Commun. 2023, 4, 284–287. [Google Scholar] [CrossRef]
- Hong, H.A.; Le, H.D.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Bernardeau, M.; Lehtinen, M.J.; Forssten, S.D.; Nurminen, P. Importance of the gastrointestinal life cycle of Bacillus for probiotic functionality. J. Food Sci. Technol. 2017, 54, 2570–2584. [Google Scholar] [CrossRef]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. probiotic strains as a potential tool for limiting the use of antibiotics and improving the growth and health of pigs and chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on Lactiferm® (Enterococcus faecium) as a feed additive for weaned piglets and calves. EFSA J. 2012, 10, 2574. [Google Scholar] [CrossRef]
- CSN EN-15784; Animal Feeding Stuffs: Methods of Sampling and Analysis—Detection and Enumeration of Bacillus spp. Used as Feed Additive. European Standards s.r.o.: Pilsen, Czech Republic, 2021.
- CSN EN-15788; Animal Feeding Stuffs: Methods of Sampling and Analysis—Detection and Enumeration of Enterococcus (E. faecium) spp. Used as Feed Additive. European Standards s.r.o.: Pilsen, Czech Republic, 2021.
- Copani, G.C.; Queiroz, O.C.M.; Boll, E.J. Lactobacillus animalis LA51 and Bacillus sp. probiotics confer protection from the damaging effects of pathogenic Clostridium perfringens and Escherichia coli on the intestinal barrier. J. Dairy Sci. 2020, 103 (Suppl. S1), 103. [Google Scholar]
- Guimarães, O.; Cappellozza, B.I.; Capern, L.C.; Schutz, J.; Cull, C.A.; Queiroz, O.; Copani, G. A novel direct-fed microbial mixture for beef cattle has a supportive effect against Clostridium perfringens in vitro and in vivo. Ruminants 2023. submitted. [Google Scholar]
- La Fata, G.; Weber, P.; Mohajeri, M. Probiotics and the gut immune system: Indirect regulation. Probiotics Antimicro. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- NASEM. Nutrient Requirements of Beef Cattle Model, 8th ed.; National Academic Press: Washington, DC, USA, 2016. [Google Scholar]
- Fritzen-Freire, C.B.; Prudêncio, E.S.; Amboni, R.D.M.C.; Pinto, S.S.; Negrão-Murakami, A.N.; Murakami, F.S. Microencapsulation of bifidobacteria by spray drying in the presence of prebiotics. Food Res. Int. 2012, 45, 306–312. [Google Scholar] [CrossRef]
- Mathews, S. Microencapsulation of probiotics by calcium alginate and gelatin and evaluation of its survival in simulated human gastro-intestinal condition. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 2080–2087. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Sharma, D.; Chauhan, R.; Goel, G. Skimmed milk-based encapsulation for enhanced stability and viability of Lactobacillus gastricus BTM 7 under simulated gastrointestinal conditions. Probiotics Antimicrob. Proteins 2019, 11, 850–856. [Google Scholar] [CrossRef]
- Shridhar, P.B.; Amachawadi, R.G.; Tokach, M.; Patel, I.; Gangiredla, J.; Mammnel, M.; Nagaraja, T.G. Whole genome sequence analyses-based assessment of virulence potential and antimicrobial susceptibilities and resistance of Enterococcus faecium strains isolated from commercial swine and cattle probiotic products. J. Anim. Sci. 2022, 100, skac030. [Google Scholar] [CrossRef]
- Biricik, H.; Brav, F.C.; Çetin, E.; Aydin, L.; Fantinati, P.; Cappellozza, B.I. Effects of supplementing a direct-fed microbial containing Enterococcus faecium 669 on performance, health, and metabolic responses of pre-weaning Holstein dairy calves. J. Dairy Sci. 2023; in press. [Google Scholar]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Schallmey, M.; Sigh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cappellozza, B.I.; Joergensen, J.N.; Copani, G.; Bryan, K.A.; Fantinati, P.; Bodin, J.-C.; Malek Khahi, M.; NinoDeGuzman, C.; Arriola, K.G.; Lima, L.O.; et al. Evaluation of a Bacillus-based direct-fed microbial probiotic on in vitro rumen gas production and nutrient digestibility of different feedstuffs and total mixed rations. Transl. Anim. Sci. 2023, 7, txad044. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.A.; Cooke, R.F.; Brandão, A.P.; Wiegand, J.B.; Schubach, K.M.; Sowers, C.A.; Duff, G.C.; Block, E.; Gouvêa, V. Performance, health, and physiological responses of newly received feedlot cattle supplemented with pre- and probiotic ingredients. Animal 2021, 15, 100214. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, Z.M.; Górka, P.; Schlagheck, A.; Jagusiak, W.; Micek, P.; Strzetelski, J. Performance of Holstein calves fed milk-replacer and starter mixture supplemented with probiotic feed additive. J. Anim. Feed Sci. 2009, 18, 399–411. [Google Scholar] [CrossRef]
- Sun, P.; Wang, J.Q.; Zhang, H.T. Effects of supplementation of Bacillus subtilis natto Na and N1 strains on rumen development in dairy calves. Anim. Feed Sci. Technol. 2011, 164, 154–160. [Google Scholar] [CrossRef]
- Lucey, P.M.; Lean, I.J.; Aly, S.S.; Golder, H.M.; Block, E.; Thompson, J.S.; Rossow, H.A. Effects of mannan-oligosaccharide and Bacillus subtilis supplementation to preweaning Holstein dairy heifers on body weight gain, diarrhea, and shedding of fecal pathogens. J. Dairy Sci. 2021, 104, 4290–4302. [Google Scholar] [CrossRef]
- Sun, P.; Wang, J.Q.; Deng, L.F. Effects of Bacillus subtilis natto on milk production, rumen fermentation and ruminal microbiome of dairy cows. Animal 2013, 7, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Souza, V.L.; Lopes, N.M.; Zacaroni, O.F.; Silveira, V.A.; Pereira, R.A.N.; Freitas, J.A.; Almeida, R.; Salvati, G.G.S.; Pereira, M.N. Lactation performance and diet digestibility of dairy cows in response to the supplementation of Bacillus subtilis spores. Livest. Sci. 2017, 200, 35–39. [Google Scholar] [CrossRef]
- Oyebade, A.; Lee, S.; Arriola, K.; Sultana, H.; Duvalsaint, E.; Amaro, F.; Marenchino, I.F.; Nino De Guzman, C.; Pacheco, L.M.; Mu, L.; et al. Effects of probiotics supplementation on production performance of early-lactation dairy cows. J. Dairy Sci. 2021, 104 (Suppl. S1), 99. [Google Scholar]
- Amerah, A.; Quiles, A.; Medel, P.; Sanchez, J.; Lehtinen, M.; Gracia, M. Effect of pelleting temperature and probiotic supplementation on growth performance and immune function of broilers fed maize/soy-based diets. Anim. Feed Sci. Technol. 2013, 180, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Welboren, A.C.; Van Baal, J.; Ghaffari, M.H.; Sauerwein, H.; Renaud, J.B.; Martín-Tereso, J.; Steele, M.A.; Leal, L.N. Gastrointestinal permeability and inflammatory status of preweaning dairy calves in response to decreasing the ratio of n-6 to n-3 fatty acid in milk replacer. J. Dairy Sci. 2023, 106, 22568. [Google Scholar] [CrossRef] [PubMed]
- Cobos, M.A.; Valencia, L.H.; Pinos-Rodríguez, J.M.; González-Muñoz, S.S.; Ramírez, J.; Hernández, D. Influence of yeast products in rumen microorganisms, in vitro degradation and fermentation of a diet for steers. J. Appl. Anim. Res. 2010, 37, 129–133. [Google Scholar] [CrossRef]
- Teixeira, D.A.A.; Cappellozza, B.I.; Fernandes, J.R.; Nascimento, K.S.; Bonfim, L.E.L.M.; Lopes, C.N.; Ehrhardt, J.A.C.; Peres, J.R.; Harris, S.A.; Simas, J.M.C.; et al. Effects of monensin on in vitro rumen fermentation characteristics and performance of Bos indicus beef bulls offered a high-concentrate diet. Transl. Anim. Sci. 2020, 4, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Sitta, C. Additives (Ionophore and No Ionophore), Corn Grain Processing and Level of Neutral Detergent Fiber from Corn Silage on Finishing Cattle Diets. Ph.D. Thesis, University of São Paulo, São Paulo, Brazil, 2016; p. 121. [Google Scholar]


{kind=link}
| Temperature, °C | Expected, log10 CFU/gram of Pellet | Observed, log10 CFU/gram of Pellet |
|---|---|---|
| 75 | 5.58 | 5.58 ± 0.025 |
| 85 | 5.58 | 5.58 ± 0.025 |
| 95 | 5.58 | 5.64 ± 0.025 |
| Time, Months | Observed, log10 CFU/gram of Premix | p = (Versus T0) |
|---|---|---|
| T0 | 7.60 ± 0.029 | -- |
| T1 | 7.56 ± 0.029 | 0.44 |
| T3 | 7.66 ± 0.029 | 0.22 |
| T6 | 7.60 ± 0.029 | 0.98 |
| T12 | 7.51 ± 0.041 | 0.17 |
| Item | Observed, log10 CFU/gram of Milk Replacer |
|---|---|
| Temperature, °C | |
| 37 | 6.01 ± 0.021 |
| 50 | 6.00 ± 0.021 |
| Time, minutes | |
| T0 | 6.02 ± 0.025 |
| T30 | 6.02 ± 0.025 |
| T60 | 5.99 ± 0.025 |
| Time, Months | Observed, log10 CFU/gram of Premix | p = (Versus T0) |
|---|---|---|
| T0 | 7.61 ± 0.054 | -- |
| T1 | 7.64 ± 0.054 | 0.64 |
| T3 | 7.75 ± 0.054 | 0.13 |
| T6 | 7.63 ± 0.054 | 0.74 |
| T12 | 7.54 ± 0.054 | 0.42 |
| Item | Observed, log10 CFU/gram of Milk Replacer Preparation |
|---|---|
| Condition 2 | |
| 1 | 6.46 ± 0.018 |
| 2 | 6.47 ± 0.018 |
| 3 | 6.44 ± 0.018 |
| Time, Months | Observed, log10 CFU/gram of Milk Replacer | p = (Versus T0) |
|---|---|---|
| T0 | 6.95 ± 0.027 | -- |
| T1 | 7.00 ± 0.027 | 0.27 |
| T3 | 7.00 ± 0.027 | 0.27 |
| T6 | 6.95 ± 0.027 | 1.00 |
| T12 | 6.90 ± 0.027 | 0.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappellozza, B.I.; Segura, A.; Milora, N.; Galschioet, C.; Schjelde, M.; Copani, G. Stability of Bacillus and Enterococcus faecium 669 Probiotic Strains When Added to Different Feed Matrices Used in Dairy Production. Animals 2023, 13, 2350. https://doi.org/10.3390/ani13142350
Cappellozza BI, Segura A, Milora N, Galschioet C, Schjelde M, Copani G. Stability of Bacillus and Enterococcus faecium 669 Probiotic Strains When Added to Different Feed Matrices Used in Dairy Production. Animals. 2023; 13(14):2350. https://doi.org/10.3390/ani13142350
Chicago/Turabian StyleCappellozza, Bruno I., Audrey Segura, Nina Milora, Christel Galschioet, Mette Schjelde, and Giuseppe Copani. 2023. "Stability of Bacillus and Enterococcus faecium 669 Probiotic Strains When Added to Different Feed Matrices Used in Dairy Production" Animals 13, no. 14: 2350. https://doi.org/10.3390/ani13142350
APA StyleCappellozza, B. I., Segura, A., Milora, N., Galschioet, C., Schjelde, M., & Copani, G. (2023). Stability of Bacillus and Enterococcus faecium 669 Probiotic Strains When Added to Different Feed Matrices Used in Dairy Production. Animals, 13(14), 2350. https://doi.org/10.3390/ani13142350