

A Cadaveric Study Using Computed Tomography for Measuring the Ocular Bulb and Scleral Skeleton of the Atlantic Puffin (Aves, Alcidae, Fratercula arctica)
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. CT Technique
2.3. Measurements
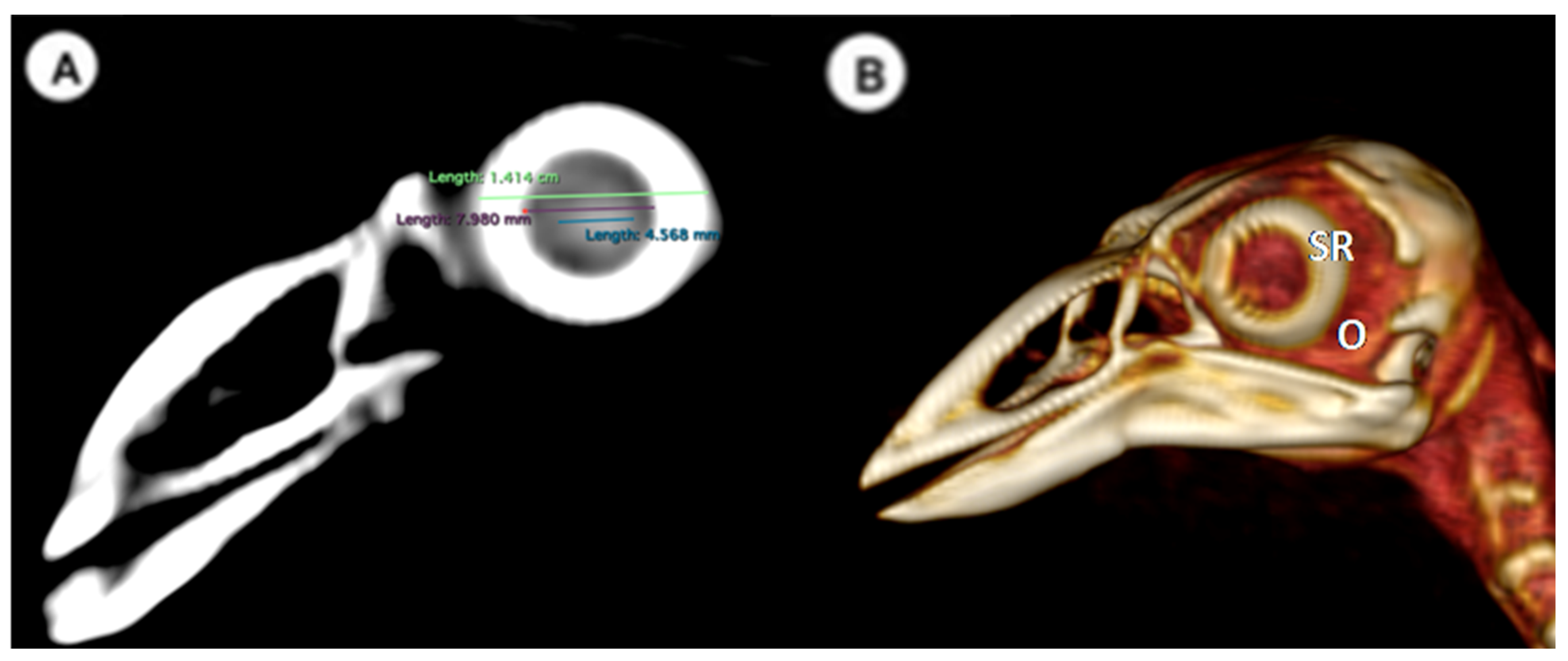
- (A)
- Measurements in the transverse plane relative to the ocular bulb:
- -
- Lens diameter: This parameter refers to the maximum distance between the lateral and medial edges of the lens, also known as the equatorial diameter (Figure 1A);
- -
- Internal diameter of the sclerotic ring, which represents the maximum distance between the inner lateromedial edges of the ring close to the cornea (Figure 1A);
- -
- External diameter of the sclerotic ring, which corresponds to the maximum distance between the outer lateromedial edges of the ring close to the sclera (Figure 1A);
- -
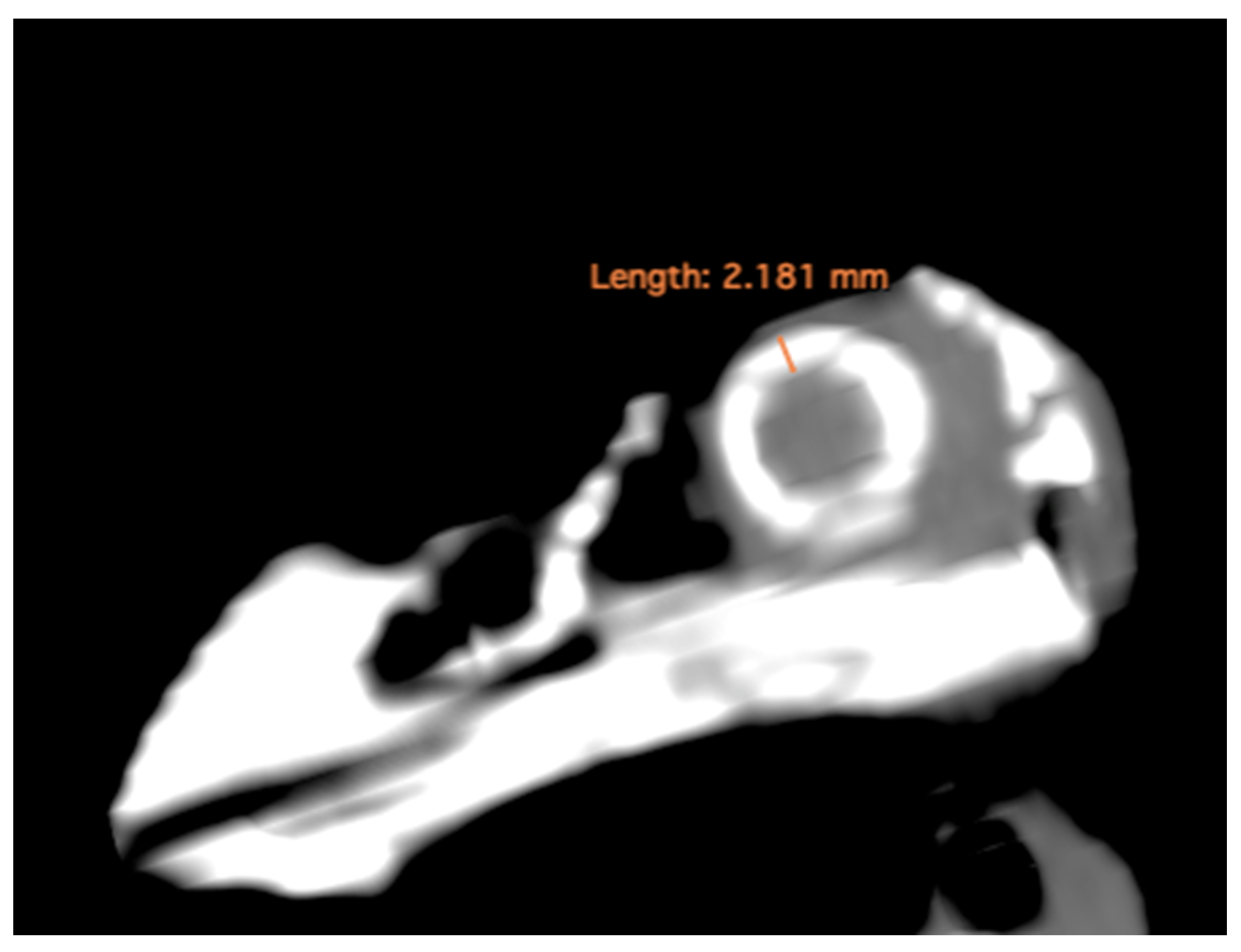
- Thickness of the sclerotic ring, defined as the distance between the internal diameter of the sclerotic ring and the external diameter, measured in the dorsal arch (Figure 2).
- (B)
- Transverse plane concerning the puffin’s body
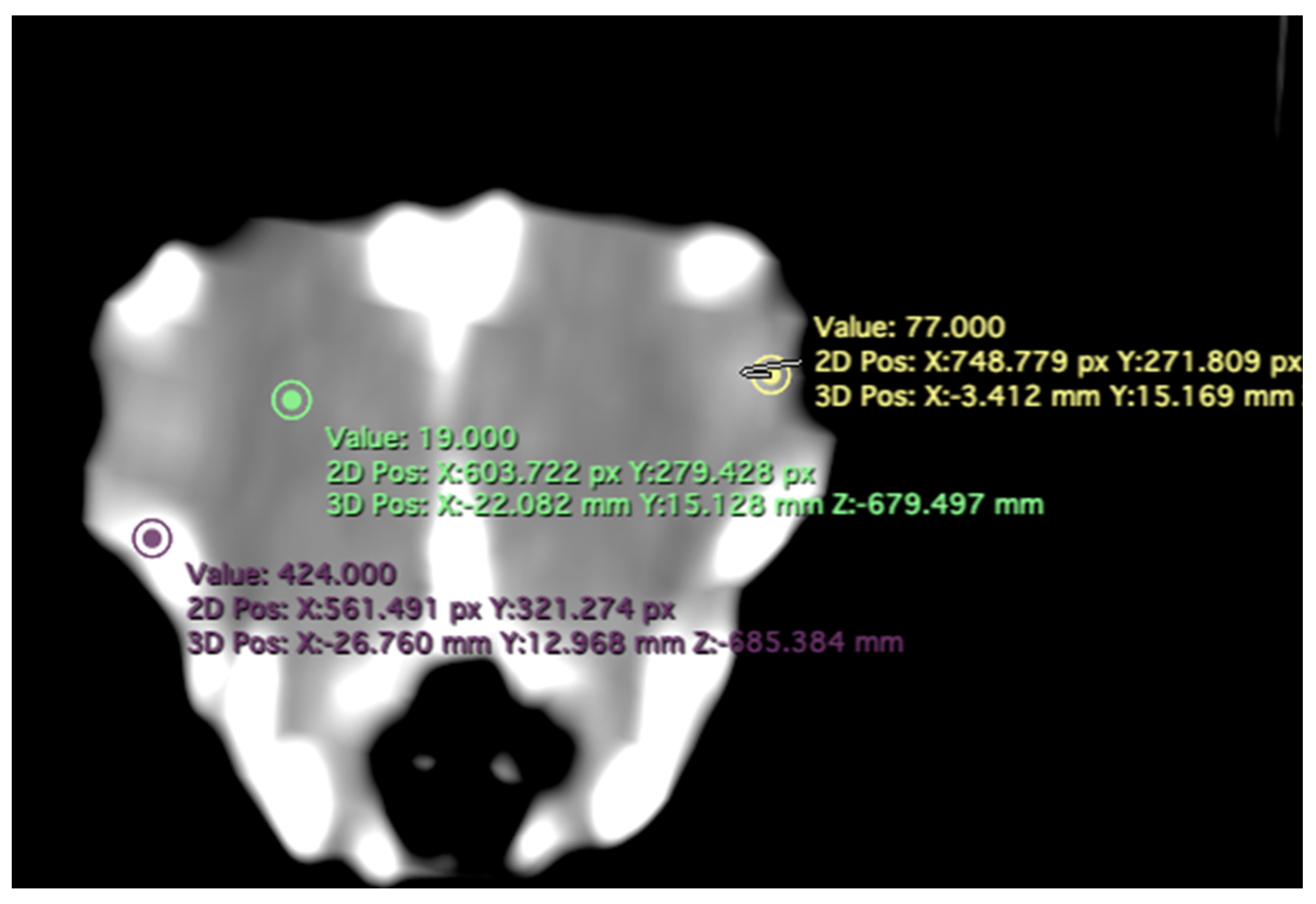
- -
- Attenuation of the sclerotic ring and lens: This measurement was taken in the dorsal area and expressed in Hounsfield Units, providing information about the radiodensity of the sclerotic ring, lens, and vitreous humor (Figure 4).
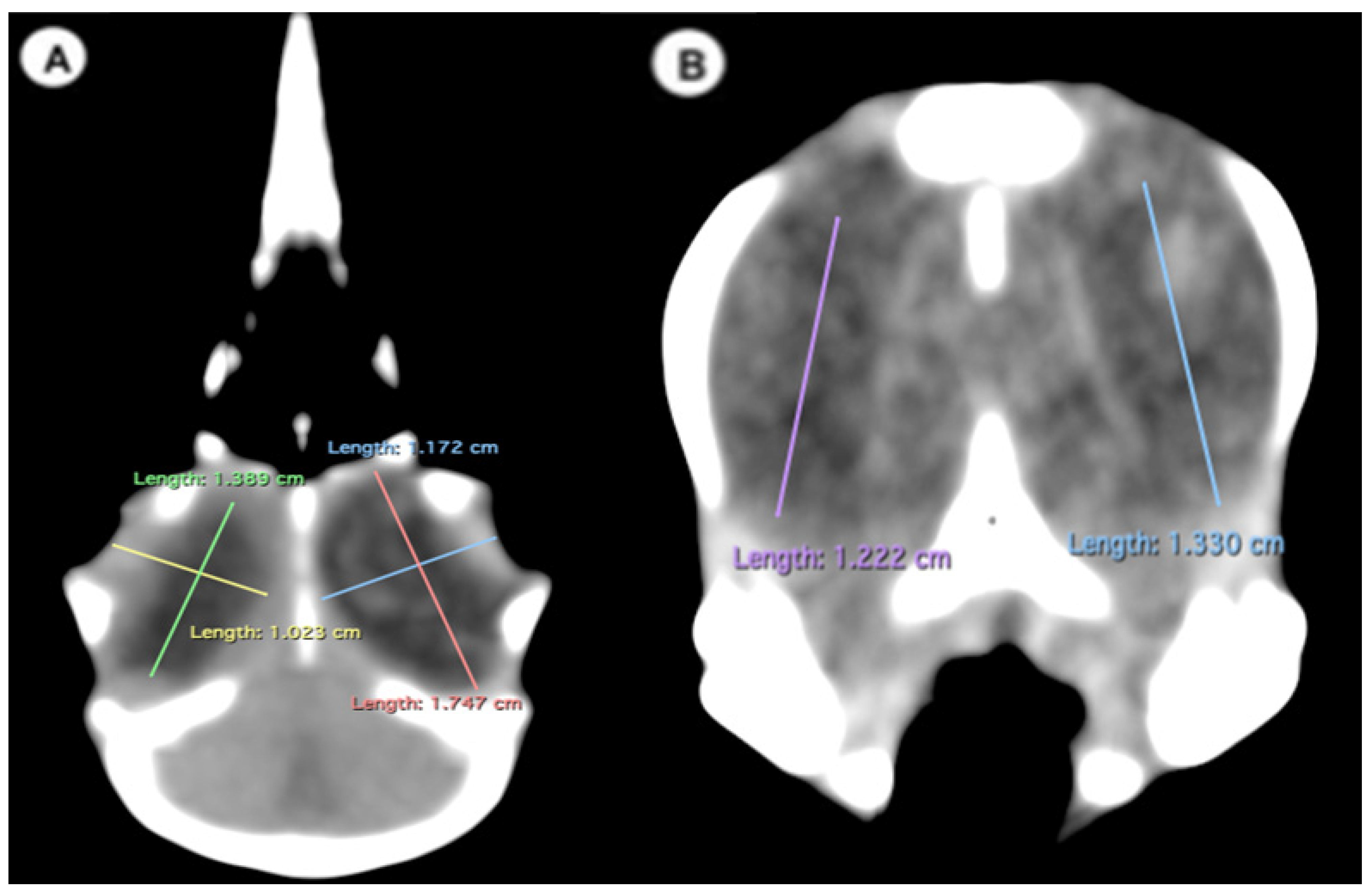
- (C)
- Dorsal plane
- -
- Width of the ocular bulb: This measurement represents the lateromedial length of the ocular bulb, extending from the os lacrimale to the inner face of the os frontale (Figure 3A);
- -
- Length of the ocular bulb: This measurement refers to the distance between the most rostral and caudal portions of the ocular bulb, spanning from the septum interorbitale to the processus antorbitalis (Figure 3A).
- (D)
- Three-dimensional reconstruction
- -
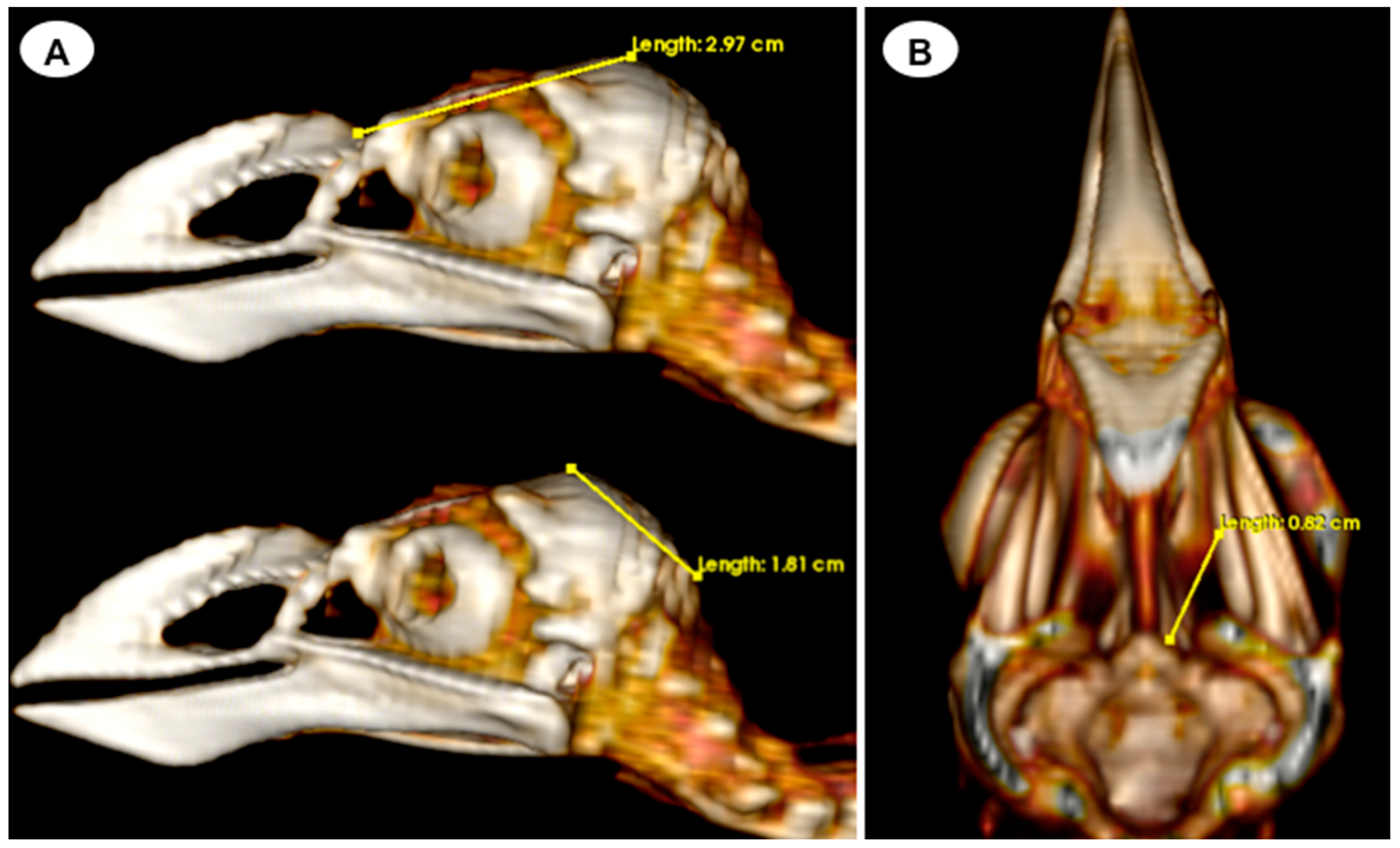
- Head length: This measurement is obtained from the centre point of the junction between the beak joint to the prominentia cerebellaris, providing the length of the head (Figure 5A);
- -
- Depth of the orbit: This measurement represents the distance between the midpoint of the orbital diameter and the foramen opticum, indicating the depth of the orbit (Figure 5B).
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- BirdLife International. Fratercula arctica. In The IUCN Red List of Threatened Species; 2018; p. e.T22694927A132581443. Available online: https://www.iucnredlist.org/especies/22694927/132581443 (accessed on 12 July 2023).
- Myers, P.; Espinosa, R.; Parr, C.S.; Jones, T.; Hammond, G.S.; Dewey, T.A. The Animal Diversity Web (Online). 2023. Available online: https://animaldiversity.org (accessed on 25 June 2023).
- Breton, A.; Diamond, A.; Kress, S. Encounter, survival and movement probabilities from an Atlantic puffin (Fratercula arctica) metapopulation. Ecol. Monogr. 2006, 76, 133–149. [Google Scholar] [CrossRef] [Green Version]
- Durant, J.; Tycho, A.; Nills, C. Ocean climate prior to breeding affects the duration of the nestling period in the Atlantic puffin. Biol. Lett. 2006, 2, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Lowther, P.E.; Diamond, A.W.; Kress, S.W.; Robertson, G.J.; Russell, K. The Birds of North America; The Birds of North America, Inc.: Philadelphia, PA, USA, 2002; pp. 1–23. [Google Scholar]
- Rodway, M. Relationship between wing length and body mass in atlantic puffin chicks. J. Field Ornithol. 1997, 14, 338–347. [Google Scholar]
- Burnham, K.K.; Burnham, J.L.; Johnson, J.A.; Huffman, A. Migratory movements of Atlantic puffins Fratercula arctica naumanni from high Arctic Greenland. PLoS ONE 2021, 16, e0252055. [Google Scholar] [CrossRef]
- Harris, M.; Daunt, F.; Newell, M.; Phillips, R.; Wanless, S. Wintering areas of adult Atlantic Puffins Fratercula arctica from a North Sea colony as revealed by geolocation technology. Mar. Biol. 2010, 157, 827–836. [Google Scholar] [CrossRef]
- Guilford, T.; Freeman, R.; Boyle, D.; Dean, B.; Kirk, H.; Phillips, R.; Perrins, C. A Dispersive Migration in the Atlantic Puffin and Its Implications for Migratory Navigation. PLoS ONE 2011, 6, e21336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramp, S. The Birds of the Western Palearctic, Full ed.; Oxford University Press: Oxford, UK, 1985. [Google Scholar]
- Boag, D.; Alexander, M. The Puffin; Blandford Press: London, UK, 1995. [Google Scholar]
- Creelman, E.; Storey, A.E. Sex differences in reproductive behavior of Atlantic Puffins. Condor 1991, 93, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Anker-Nilssen, T. The breeding performance of Puffins Fratercula arctica on Røst, northern Norway in 1979–1985. Fauna Norv. 1987, 10, 21–38. [Google Scholar]
- Harris, M.P.; Hislop, J.R.G. The food of young Puffins Fratercula arctica. J. Zool. 1978, 185, 213–236. [Google Scholar] [CrossRef]
- Falk, K.; Jensen, J.K.; Kampp, K. Winter diet of Atlantic puffins (Fratercula arctica) in the northeast Atlantic. Col. Waterbirds 1992, 15, 230–235. [Google Scholar] [CrossRef]
- Durant, J.; Anker-Nilssen, T.; Stenseth, N.C. Trophic interactions under climate fluctuations: The Atlantic puffin as an example. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, H.; Erikstad, K.E.; Barrett, R.T.; Yoccoz, N.G. The effect of climate on adult survival in five species of North Atlantic seabirds. J. Anim. Ecol. 2005, 74, 817–831. [Google Scholar] [CrossRef]
- Melillo, J.M.; Richmond, T.T.C.; Yohe, G.W. Extreme Weather. In Highlights of Climate Change Impacts in the United States: The Third National Climate Assessment; U.S. Global Change Research Program; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Wilhelm, S.I.; Mailhiot, J.; Arany, J.; Chardine, J.W.; Robertson, G.J.; Ryan, P.C. Update and trends of three important seabird populations in the western North Atlantic using a geographic information system approach. Mar. Ornithol. 2015, 43, 211–222. [Google Scholar]
- Thorup, S.H.; Jensen, J.K.; Petersen, K.T.; Kasper, D.B. Færøsk Trækfugleatlas: The Faroese Bird Migration Atlas; Faroe University Press: Tórshavn, Denmark, 2014. [Google Scholar]
- Harris, M.P.; Wanless, S. The Puffin; Poyser Monographs; Bloomsbury Publishing: London, UK, 2011. [Google Scholar]
- Martin, G.R.; Wanless, S. The visual fields of Common Guillemots Uria aalge and Atlantic Puffins Fratercula arctica: Foraging, vigilance and collision vulnerability. Ibis 2015, 157, 798–807. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.R.; White, C.R.; Butler, P.J. Vision and the foraging technique of Great Cormorants Phalacrocorax carbo: Pursuit or flush-foraging? Ibis 2008, 150, 39–48. [Google Scholar] [CrossRef]
- Regular, P.M.; Hedd, A.; Montevecchi, W.A. Fishing in the dark: A pursuit-diving seabird modifies foraging behaviour in response to nocturnal light levels. PLoS ONE 2011, 6, e26763. [Google Scholar] [CrossRef]
- Franz-Odendaal, T.A. Skeletons of the Eye: An Evolutionary and Developmental Perspective. Anat. Rec. 2020, 303, 100–109. [Google Scholar] [CrossRef]
- Bartol, S.M.; Musick, J.A.; Ochs, A.L. Visual acuity thresholds of juvenile loggerhead sea turtles (Caretta caretta): An electrophysiological approach. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2002, 187, 953–960. [Google Scholar] [CrossRef]
- Waldvogel, J.A. The bird’s eye view. Am. Sci. 1990, 78, 342–353. [Google Scholar]
- Lauridsen, H.; Hansen, K.; Wang, T.; Agger, P.; Andersen, J.L.; Knudsen, P.S.; Rasmussen, A.S.; Uhrenholt, L.; Pedersen, M. Inside Out: Modern Imaging Techniques to Reveal Animal Anatomy. PLoS ONE 2011, 6, e17879. [Google Scholar] [CrossRef] [Green Version]
- Webb, E.; Yuan, M.; Lemoine, N.; Wang, Y. Imaging in animal models. Integr. Cancer Sci. Ther. 2016, 3, 428–431. [Google Scholar] [CrossRef]
- Schambach, S.J.; Bag, S.; Schilling, L.; Groden, C.; Brockmann, M.A. Application of micro-CT in small animal imaging. Methods 2010, 50, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Farrag, M.; Pukale, D.D.; Leipzig, N.D. Micro-computed tomography utility for estimation of intraparenchymal spinal cord cystic lesions in small animals. Neural Regen. Res. 2021, 16, 2293–2298. [Google Scholar] [PubMed]
- Barrett, R.T.; Fieler, R.; Anker-Nilssen, T.; Rikardsen, F. Measurements and weight changes of norwegian adult puffins Fratercula arctica and kittiwakes Rissa tridactyla during the breeding season. Ring. Migr. 1985, 6, 102–112. [Google Scholar] [CrossRef]
- Westmoreland, L.S.; Hadfield, C.A.; Clayton, L.A.; Atkins, R.M.; Hyman, J.A.; Rossi, B.H. Ocular examinations and investigation of intraocular pressure, tear production, central corneal thickness, and corneal touch threshold in a captive flock of Atlantic puffins (Fratercula arctica). J. Avian Med. Surg. 2020, 34, 123–131. [Google Scholar] [CrossRef]
- Fernández, M.S.; Archuby, F.; Talevi, M.; Ebner, R. Ichthyosaurian eyes: Paleobiological information content in the sclerotic ring of Caypullisaurus (Ichthyosauria, Ophthalmosauria). J. Vertebr. Paleontol. 2005, 25, 330–337. [Google Scholar] [CrossRef]
- Hall, M.I. The anatomical relationships between the avian eye, orbit and sclerotic ring: Implications for inferring activity patterns in extinct birds. J. Anat. 2008, 212, 781–794. [Google Scholar] [CrossRef]
- Salgüero, R.; Johnson, V.; Williams, D.; Hartley, C.; Holmes, M.; Dennis, R.; Herrtage, M. CT dimensions, volumes and densities of normal canine eyes. Vet. Rec. 2015, 176, 386. [Google Scholar] [CrossRef]
- Boroffka, S.A.E.B.; Voorhout, G. Direct and reconstructed multiplanar computed tomography of the orbits of healthy dogs. Am. J. Vet. Res. 1999, 60, 1500–1507. [Google Scholar]
- Sreenivasan, S.; Linden, A.; Owen, M.; Pemberton, S.; Pinard, C.; Matsuyama, A.; Poirier, V.J. Computed tomography measurements of intraocular structures of the feline eye. Vet. Rec. 2019, 184, 651–656. [Google Scholar]
- Fumero-Hernández, M.; Encinoso Quintana, M.; Ramírez, A.S.; Morales Fariña, I.; Calabuig, P.; Jaber, J.R. Morphometric Study of the Eyeball of the Loggerhead Turtle (Caretta caretta) Using Computed Tomography (CT). Animals 2023, 13, 1016. [Google Scholar] [CrossRef] [PubMed]
- Andrade, S.B.D.; Araujo, N.L.L.C.D.; Raposo, A.C.S.; Muramoto, C.; Oriá, A.P. Morphometric descriptive report of scleral ossicle rings, by ultrasound and computed tomography, in three Testudines specimens. Ciência Rural. 2023, 53, e20210423. [Google Scholar] [CrossRef]
- Kuszyk, B.S.; Heath, D.G.; Bliss, D.F.; Fishman, E.K. Skeletal 3-D CT: Advantages of Volume Rendering over Surface Rendering. Skelet. Radiol. 1996, 25, 207–214. [Google Scholar] [CrossRef]
- Pérez, S.; Encinoso, M.; Morales, M.; Arencibia, A.; Suárez-Bonnet, A.; González-Rodríguez, E.; Jaber, J.R. Comparative evaluation of the Komodo dragon (Varanus komodoensis) and the Green iguana (Iguana iguana) skull by three dimensional computed tomographic reconstruction. Slov. Vet. Res. 2021, 58, 111–116. [Google Scholar] [CrossRef]
- Schultz, A.H. The size of the orbit and of the eye in primates. Am. J. Phys. Anthropol. 1940, 26, 389–408. [Google Scholar] [CrossRef]
- Kay, R.F.; Cartmill, M. Cranial morphology and adaptations of Palaechthon nacimienti and other paromomyidae (plesiadapoidea,? Primates), with a description of a new genus and species. J. Hum. Evol. 1977, 6, 19–35. [Google Scholar] [CrossRef]
- Kay, R.F.; Kirk, E.C. Osteological evidence for the evolution of activity pattern and visual acuity in primates. Am. J. Phys. Anthropol. 2000, 113, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Heesy, C.P.; Ross, C.F. Evolution of activity patterns and chromatic vision in primates: Morphometrics, genetics and cladistics. J. Hum. Evol. 2001, 40, 111–149. [Google Scholar] [CrossRef] [Green Version]
- Metscher, B.D. MicroCT for comparative morphology: Simple staining methods allow high-contrast 3D imaging of diverse non-mineralized animal tissues. BMC Physiol. 2009, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Right Eye | Left Eye | Both Eyes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Median | Range | SD | Mean | Median | Range | SD | Mean | Median | Range | SD | |
| Lens diameter (mm) | 3.34 | 3.3 | 2.7–4.2 | 0.36 | 3.30 | 3.2 | 2.8–3.9 | 0.29 | 3.32 | 3.3 | 2.7–4.2 | 0.32 |
| Internal diameter of the sclerotic ring (cm) | 0.64 | 0.63 | 0.59–0.69 | 0.03 | 0.63 | 0.63 | 0.59–0.7 | 0.03 | 0.63 | 0.63 | 0.59–0.7 | 0.03 |
| External diameter of the sclerotic ring (cm) | 1.44 | 1.44 | 1.38–1.48 | 0.03 | 1.43 | 1.43 | 1.38–1.49 | 0.03 | 1.44 | 1.44 | 1.38–1.49 | 0.03 |
| Sclerotic ring thickness (mm) | 2.15 | 2.14 | 2.04–2.28 | 0.07 | 2.18 | 2.195 | 2.08–2.29 | 0.06 | 2.16 | 2.16 | 2.04–2.29 | 0.06 |
| Ocular bulb height (cm) | 1.54 | 1.52 | 1.47–1.61 | 0.04 | 1.52 | 1.52 | 1.41–1.61 | 0.05 | 1.53 | 1.52 | 1.41–1.61 | 0.05 |
| Ocular bulb width (cm) | 1.29 | 1.29 | 1.1–1.42 | 0.09 | 1.27 | 1.28 | 1.1–1.39 | 0.08 | 1.28 | 1.29 | 1.1–1.42 | 0.08 |
| Ocular bulb length (cm) | 1.62 | 1.64 | 1.46–1.7 | 0.07 | 1.60 | 1.6 | 1.48–1.71 | 0.07 | 1.61 | 1.62 | 1.46–1.71 | 0.07 |
| Orbit depth (mm) | 8.16 | 8.2 | 7.9–8.4 | 0.16 | 8.23 | 8.3 | 7.9–8.5 | 0.18 | 8.19 | 8.2 | 7.9–8.5 | 0.18 |
| Lens attenuation (HU) | 70.14 | 71 | 64–79 | 2.95 | 68.93 | 69 | 64–74 | 2.72 | 69.53 | 69.5 | 64–79 | 2.88 |
| Vitreous humor attenuation (HU) | 35.17 | 35 | 31–41 | 3.06 | 35.59 | 36 | 31–45 | 3.76 | 35.38 | 35.5 | 31–45 | 3.40 |
| Sclerotic ring attenuation (dorsal arch) (HU) | 721.21 | 725 | 687–752 | 18.39 | 719.79 | 723 | 687–745 | 17.64 | 720.50 | 723 | 687–752 | 17.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fumero-Hernández, M.; Encinoso, M.; Ramírez, A.S.; Morales, I.; Suárez Pérez, A.; Jaber, J.R. A Cadaveric Study Using Computed Tomography for Measuring the Ocular Bulb and Scleral Skeleton of the Atlantic Puffin (Aves, Alcidae, Fratercula arctica). Animals 2023, 13, 2418. https://doi.org/10.3390/ani13152418
Fumero-Hernández M, Encinoso M, Ramírez AS, Morales I, Suárez Pérez A, Jaber JR. A Cadaveric Study Using Computed Tomography for Measuring the Ocular Bulb and Scleral Skeleton of the Atlantic Puffin (Aves, Alcidae, Fratercula arctica). Animals. 2023; 13(15):2418. https://doi.org/10.3390/ani13152418
Chicago/Turabian StyleFumero-Hernández, Marcos, Mario Encinoso, Ana Sofia Ramírez, Inmaculada Morales, Alejandro Suárez Pérez, and José Raduan Jaber. 2023. "A Cadaveric Study Using Computed Tomography for Measuring the Ocular Bulb and Scleral Skeleton of the Atlantic Puffin (Aves, Alcidae, Fratercula arctica)" Animals 13, no. 15: 2418. https://doi.org/10.3390/ani13152418
APA StyleFumero-Hernández, M., Encinoso, M., Ramírez, A. S., Morales, I., Suárez Pérez, A., & Jaber, J. R. (2023). A Cadaveric Study Using Computed Tomography for Measuring the Ocular Bulb and Scleral Skeleton of the Atlantic Puffin (Aves, Alcidae, Fratercula arctica). Animals, 13(15), 2418. https://doi.org/10.3390/ani13152418