


Sirtuin 1 Expression in Canine Mammary Tumors: A Pilot Study
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Canine Mammary Tissue Collection for Histology
2.2. Immunohistochemistry
2.3. Cell Lines and Cultures
2.4. Western Blot Analyses
2.5. Immunofluorescence
2.6. Statistical Analysis
3. Results
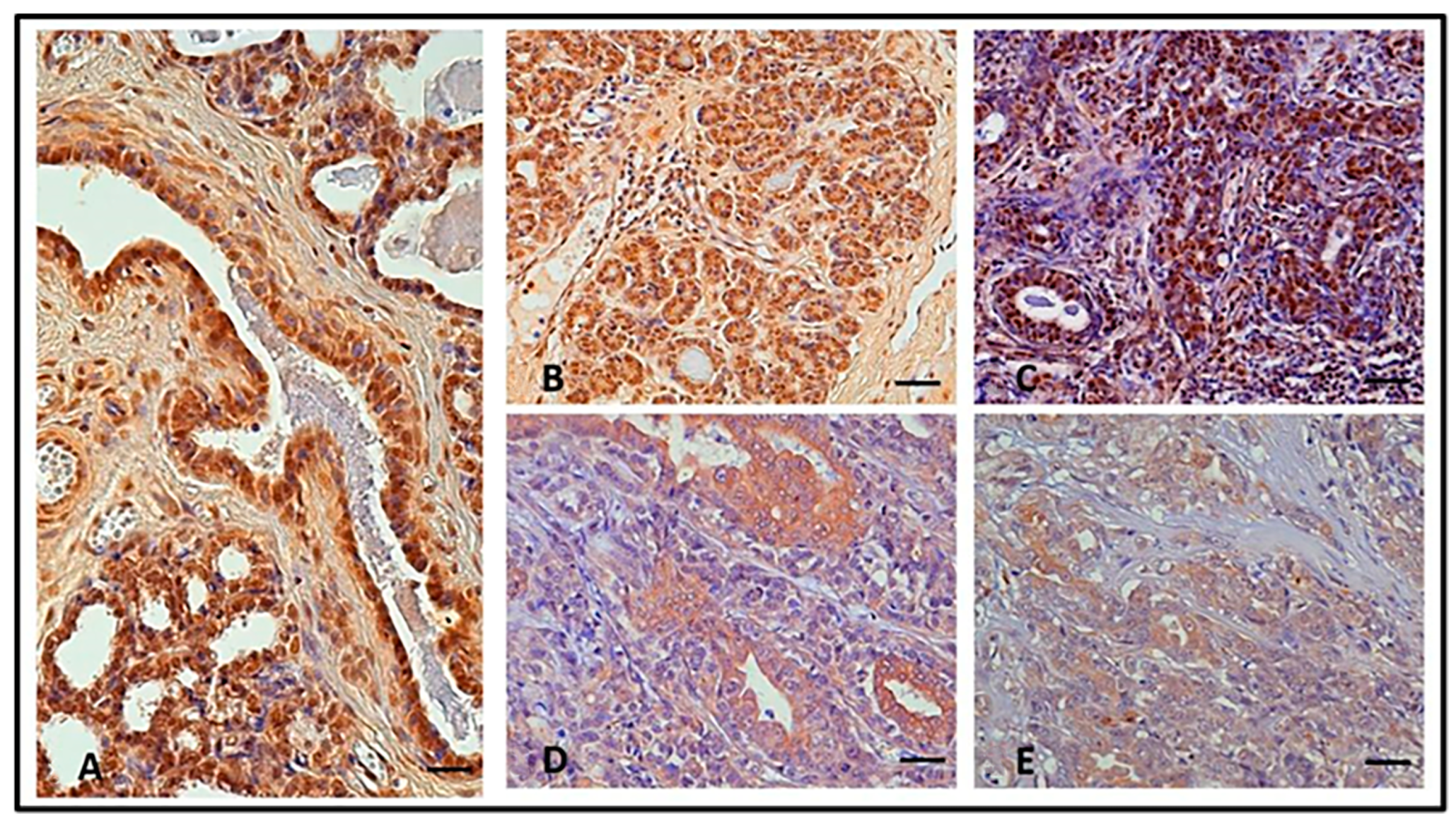
3.1. Immunohistochemistry
3.2. Western Blot
3.3. Immunofluorescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haigis, M.C.; Sinclair, D.A. Mammalian sirtuins: Biological insights and disease relevance. Annu. Rev. Pathol. 2010, 5, 253–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.C.; Guarente, L. SIRT1 and other sirtuins in metabolism. Trends. Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef]
- Levine, D.C.; Kuo, H.S.; Hong, H.-K.; Cedernaes, J. NADH inhibition of SIRT1 links energy state to transcription during time-restricted feeding. Nat. Metab. 2021, 12, 1621–1632. [Google Scholar] [CrossRef] [PubMed]
- Kida, Y.; Goligorsky, M.S.M. Sirtuins, Cell Senescence, and Vascular Aging. Can. J. Cardiol. 2016, 32, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, S.; Cristopher, M.A.; Kaeberlein, M.; Guarente, L. Transcriptional silencing, and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature 2000, 403, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, D.A.; Guarente, L. Unlocking the secrets of longevity genes. Sci. Am. 2006, 294, 48–57. [Google Scholar] [CrossRef]
- Bai, W.; Zhang, X. Nucleus or cytoplasm? The mysterious case of SIRT1′s subcellular localization. Cell Cycle News Views 2016, 24, 3337–3338. [Google Scholar] [CrossRef] [Green Version]
- Alves-Fernandes, D.K.; Jasiulionis, M.G. The Role of SIRT1 on DNA Damage Response and Epigenetic Alterations in Cancer. Int. J. Mol. Sci. 2019, 20, 31153–31165. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Peterson, L.M.; Xiaoling, L.I. Trending topics of SIRT1 in tumorigenicity. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2021, 865, 129952. [Google Scholar] [CrossRef]
- Marfe, G.; De Martino, L.; Tafani, M.; Pasolini, M.P.; Navas, L.; Papparella, S.; Gambacurta, A.; Paciello, O. A multicancer-like syndrome in a dog characterized by p53 and cell cycle-checkpoint kinase 2 (CHK2) mutations and sirtuin gene (SIRT1) down-regulation. Res. Vet. Sci. 2010, 93, 240–245. [Google Scholar] [CrossRef]
- Cacciola, N.A.; Sgadari, M.; Petillo, O.; Margarucci, S.; Martano, M.; Cocchia, N.; Maiolino, P.; Restucci, B. Carnitine palmitoyltransferase 1 A expression profile in canine mammary tumors. Vet. J. 2020, 257, 105453. [Google Scholar] [CrossRef] [PubMed]
- Cacciola, N.A.; Sgadari, M.; Petillo, O.; Margarucci, S.; Martano, M.; Cocchia, N.; Maiolino, P.; Restucci, B. Metabolic Flexibility in Canine Mammary Tumors: Implications of the Carnitine System. Animals 2021, 11, 2969. [Google Scholar] [CrossRef]
- Visan, S.; Balacescu, O.; Berindan-Neagoe, I.; Catoi, C. In vitro comparative models for canine and human breast cancers. Clujul. Med. 2016, 89, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goebel, K.; Merner, N.D. A monograph proposing the use of canine mammary tumours as a model for the study of hereditary breast cancer susceptibility genes in humans. Vet. Med. Sci. 2017, 3, 51–62. [Google Scholar] [CrossRef]
- Abdelmegeed, S.M.; Mohammed, S.M. Canine mammary tumors as a model for human disease. A review. Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahabi, K.; Gayathri, T.S.; Rasedee, A.; Cheah, Y.K.; Tan, G.C. (2018). Comparative aspects of microRNA expression in canine and human cancers. J. Vet. Sci. 2018, 19, 162–171. [Google Scholar] [CrossRef]
- Liu, D.; Xiong, H.; Ellis, A.E.; Northrup, N.C.; Rodriguez Jr, C.O.; O’Regan, R.M.; Dalton, S.; Zhao, S. Molecular homology and difference between spontaneous canine mammary cancer and human breast cancer. Cancer Res. 2014, 74, 5045–5056. [Google Scholar] [CrossRef] [Green Version]
- Zappulli, V.; Peña, L.; Rasotto, R.; Goldschmidt, M.; Gama, A.; Scruggs, J. Volume 2: Mammary Tumors. In Kiupel, Surgical Pathology of Tumors of Domestic Animals; Davis-Thompson DVM Foundation: Washington, DC, USA, 2019. [Google Scholar]
- Pena, L.; De Andres, P.J.; De Andres, M.; Cuesta, P.; Perez-Alenza, M.D. Prognostic value of histological grading in noninflammatory canine mammary carcinomas in a prospective study with two-year follow-up: Relationship with clinical and histological characteristics. Vet. Pathol. 2013, 50, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Burrai, G.P.; Tanca, A.; Cubeddu, T.; Abbondio, M.; Polinas, M.; Addis, M.F.; Antuofermo, E. A first immunohistochemistry study of transketolase and transketolase-like 1 expression in canine hyperplastic and neoplastic mammary lesions. BMC Vet. Res. 2017, 13, 38. [Google Scholar] [CrossRef] [Green Version]
- Fedchenko, N.; Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue—A review. Diagn. Pathol. 2014, 9, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Altamura, G.; Uberti, B.D.; Galiero, G.; Martano, M.; Pirro, A.; Russo, M.; Borzacchiello, G. Expression and activation of platelet-derived growth factor beta receptor, mitogen-activated protein/extracellular signal-regulated kinase kinase (MEK) and extracellular signal-regulated kinase (ERK) in canine mammary tumours. Res. Vet. Sci. 2017, 110, 29–33. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawey, A.E. Calculate the Corrected Total Cell Fluorescence (CTCF). Res. Gate 2016. [Google Scholar] [CrossRef]
- Rivera, P.; von Euler, H. Molecular biological aspects on canine and human mammary tumors. Vet. Pathol. 2011, 48, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine mammary tumours as a model to study human breast cancer: Most recent findings. Vivo 2011, 25, 455–465. [Google Scholar]
- Hiraike, H.; Wada-Hiraike, O.; Nakagawa, S. Expression of DBC1 is associated with nuclear grade and HER2 expression in breast cancer. Exp. Ther. Med. 2011, 2, 1105–1109. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Wei, W.; Xiao, X. Expression of SIRT1 is associated with lymph node metastasis and poor prognosis in both operable triple-negative and non-triple-negative breast cancer. Med. Oncol. 2012, 29, 3240–3249. [Google Scholar] [CrossRef]
- Lee, H.; Kim, K.R.; Noh, S.J. Expression of DBC1 and SIRT1 is associated with poor prognosis for breast carcinoma. Hum. Pathol. 2011, 42, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Kimberly, R.; Holloway, K.R.; Barbieri, A.; Malyarchuk, S.; Saxena, M. SIRT1 Positively Regulates Breast Cancer Associated Human Aromatase (CYP19A1) Expression. Mol. Endocrinol. 2013, 27, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, B.; Wong, N. Sirtuin 1 is upregulated in a subset of hepatocellular carcinomas where it is essential for telomere maintenance and tumor cell growth. Cancer Res. 2011, 71, 4138–4149. [Google Scholar] [CrossRef] [Green Version]
- Jang, K.Y.; Noh, S.J.; Lehwald, N. SIRT1 and c-Myc promote liver tumor cell survival and predict poor survival of human hepatocellular carcinomas. PLoS ONE 2012, 7, 45119. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.N.; Bae, J.S.; Jamiyandorj, U. Expression and role of SIRT1 in hepatocellular carcinoma. Oncol. Rep. 2011, 26, 503–510. [Google Scholar] [CrossRef]
- Wang, C.; Yang, W.; Dong, F. The prognostic role of Sirt1 expression in solid malignancies: A meta-analysis. Oncotarget 2017, 8, 66343–66351. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Chu, Z.; Ma, P.; Meng, Y.; Yang, Y. SIRT1 promotes the proliferation and metastasis of human pancreatic cancer cells. Tumor Biol. 2017, 39, 101042831769118. [Google Scholar] [CrossRef] [Green Version]
- Firestein, R.; Blander, G.; Michan, S. The SIRT1 deacetylase suppresses intestinal tumorigenesis and colon cancer growth. PLoS ONE 2008, 3, e2020. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.-H.; Min, K.-W.; Paik, S.S.; Jang, K.-S. Loss of SIRT1 histone deacetylase expression associates with tumour progression in colorectal adenocarcinoma. J. Clin. Pathol. 2012, 735–739. [Google Scholar] [CrossRef]
- Kabra, N.; Li, Z.; Chen, L.; Li, B.; Zhang, X.; Wang, C.; Yeatman, T.; Coppola, D.; Chen, J. SirT1 is an inhibitor of proliferation and tumor formation in colon cancer. J. Biol. Chem. 2009, 284, 18210–18217. [Google Scholar] [CrossRef] [Green Version]
- Jang, K.Y.; Hwang, S.H.; Kwon, K.S. SIRT1 expression is associated with poor prognosis of diffuse large B-cell lymphoma. Am. J. Surg. Pathol. 2008, 32, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, B.; Gao, W. SIRT1 is downregulated in gastric cancer and leads to G1-phase arrest via NF kappaB/cyclin D1 signaling. Mol. Cancer Res. 2013, 11, 1497–1507. [Google Scholar] [CrossRef] [Green Version]
- Jang, K.Y.; Kim, K.S.; Hwang, S.H. Expression and prognostic significance of SIRT1 in ovarian epithelial tumours. Pathology 2009, 41, 366–371. [Google Scholar] [CrossRef]
- Huffman, D.M.; Grizzle, W.E.; Bamman, M.M. SIRT1 is significantly elevated in mouse and human prostate cancer. Cancer Res. 2007, 67, 6612–6618. [Google Scholar] [CrossRef] [Green Version]
- Herranz, D.; Maraver, A.; Canamero, M. SIRT1 promotes thyroid carcinogenesis driven by PTEN deficiency. Oncogene 2013, 32, 4052–4056. [Google Scholar] [CrossRef]
- Wang, R.-H.; Sengupta, K.; Li, C. Impaired DNA damage response, genome instability, and tumorigenesis in SIRT1 mutant mice. Cancer Cell 2008, 14, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; QIN, Q.; Chen, R.; Wei, C. SIRT1 promotes proliferation, migration, and invasion of breast cancer cell line MCF-7 by upregulating DNA polymerase delta1 (POLD1). Biochem. Biophys. Res. Commun. 2018, 502, 351–357. [Google Scholar] [CrossRef]
- Kratz, E.M.; Solkiewicz, K.; Kubis-Kubiak, A.; Piwowar, A.K.-K. Sirtuins as Important Factors in Pathological States and the Role of Their Molecular Activity Modulators. Int. J. Mol. Sci. 2021, 22, 630. [Google Scholar] [CrossRef] [PubMed]
- Aventaggiato, M.; Vernucci, E.; Barreca, F.; Matteo, A. Sirtuins’ control of autophagy and mitophagy in cancer. Pharmacol Ther. 2021, 221, 107748. [Google Scholar] [CrossRef] [PubMed]
- van de Ven, R.A.H.; Santos, D.; Haigis, M.C. Mitochondrial Sirtuins and Molecular Mechanisms of Aging. Trends Mol. Med. 2017, 23, 320–331. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Wang, J.; Yang, Y.; Li, J. Mitochondrial Sirtuins in Parkinson’s Disease. Neurochem. Res. 2022, 47, 1491–1502. [Google Scholar] [CrossRef]
- Fang, Y.; Nicholl, M.B. Sirtuin 1 in malignant transformation: Friend or foe? Cancer Lett. 2010, 306, 10–14. [Google Scholar] [CrossRef]
- Carafa, V.; Altucci, l.; Nebbioso, A. Dual Tumor Suppressor and Tumor Promoter Action of Sirtuins in Determining Malignant Phenotype. Front. Pharmacol. 2019, 10, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Song, N.Y.; Surh, Y.J. Janus-faced role of SIRT1 in tumorigenesis. Ann. N. Y. Acad. Sci. 2012, 1271, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, R.D.; Harmer, D.H.; Coleman, R.A.; Clark, B.J. GAPDH mRNA expression in a panel of 72 human tissues. Physiol. Genom. 2005, 21, 389–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Shein, Y. An old method facing a new challenge: Re-visiting housekeeping protein as an internal reference control for neuroscience research. Life Sci. 2013, 92, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Greer, S.; Honeywell, R.; Geletu, M.; Arulanandam, R.; Raptis, L. Housekeeping genes; expression levels may change with density of cultured cells. J. Immunolol. Methods 2010, 355, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Harbison, C.E.; Taheri, F.; Knight, H.; Miller, A.D. Immunohistochemical Characterization of Large Intestinal Adenocarcinoma in the Rhesus Macaque (Macaca mulatta). Vet. Pathol. 2015, 52, 732–740. [Google Scholar] [CrossRef]
- Deng, C.X. SIRT1, is it a tumor promoter or tumor suppressor? Int. J. Biol. Sci. 2009, 5, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byles, V.; Chmilewski, L.K.; Wang, J.; Zhu, L. Aberrant Cytoplasm Localization and Protein Stability of SIRT1 is Regulated by PI3K/IGF-1R Signaling in Human Cancer Cells. Int. J. Biol. Sci. 2010, 6, 599–612. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.Y.; Sun, F.L.; Zhang, Y.; Wang, Z. SIRT1 acts as a potential tumor suppressor in oral squamous cell carcinoma. J. Chin. Med. Assoc. 2018, 81, 416–422. [Google Scholar] [CrossRef]
- Mvunta, D.H.; Miyamoto, T.; Asaka, R.; Yamada, Y. Overexpression of SIRT1 is Associated With Poor Outcomes in Patients With Ovarian Carcinoma. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 415–421. [Google Scholar] [CrossRef]
- Kuo, S.-J.; Lin, H.-Y.; Chien, S.-Y.; Chien, D.R. SIRT1 suppresses breast cancer growth through downregulation of the Bcl-2 protein. Oncol. Rep. 2013, 30, 125–130. [Google Scholar] [CrossRef] [Green Version]
- D’Onofrio, N.; Mele, L.; Martino, E.; Salzano, A.; Restucci, B. Synergistic Effect of Dietary Betaines on SIRT1-Mediated Apoptosis in Human Oral Squamous Cell Carcinoma. Cancers 2020, 12, 246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, Q.; Zeng, Z.; Wu, J. Sirt1 Inhibits Oxidative Stress in Vascular Endothelial Cells. Oxid. Med. Cell. Longev. 2017, 2017, 7543973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Wei, Y.; Xu, F.; Zhao, M. SIRT1 promotes formation of breast cancer through modulating Akt activity. Cancer 2018, 9, 2012–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Mammary Gland | ||||||
| No. Sample | Breed | Age (Years) | Histologic Classification | % Positive Cells | Intensity | IRS |
| 1. | Poodle | 6 | - | 78.4 | 3 | 12 |
| 2. | Mixed breed | 9 | - | 69.5 | 3 | 12 |
| 3. | Mixed breed | 5 | - | 81.2 | 3 | 12 |
| 4. | Mixed breed | 6 | - | 27.8 | 2 | 4 |
| 5. | Jack russel terrier | 8 | - | 32.4 | 3 | 9 |
| Benign Tumor | ||||||
| No. Sample | Breed | Age (Years) | Histologic Classification | % Positive Cells | Intensity | IRS |
| 1. | Yorkshire terrier | 11 | Mixed adenoma | 58.1 | 3 | 9 |
| 2. | Poodle | 12 | Benign mixed tumor | 10.8 | 1 | 2 |
| 3. | Mixed breed | 10 | Papillary cystic adenoma | 39.1 | 3 | 9 |
| 4. | Maltese | 7 | Ductal adenoma | 38.3 | 3 | 9 |
| 5. | Mixed breed | 8 | Tubular adenoma | 24.5 | 3 | 6 |
| 6. | Mixed breed | 6 | Adenoma Simple | 71.4 | 3 | 12 |
| 7. | Mixed breed | 11 | Complex Adenoma | 66.4 | 3 | 12 |
| 8. | Mixed breed | 6 | Adenoma Simple | 70.8 | 2 | 8 |
| 9. | Mixed breed | 9 | Benign Mixed Tumor | 51.3 | 2 | 6 |
| 10. | Yorkshire Terrier | 8 | Complex Adenoma | 62.6 | 3 | 12 |
| Grade 1 | ||||||
| No. Sample | Breed | Age (Years) | Histologic Classification | % Positive Cells | Intensity | IRS |
| 1. | English setter | 8 | Mixed type carcinoma | 11.3 | 2 | 4 |
| 2. | Mixed breed | 11 | Complex type carcinoma | 22.5 | 1 | 2 |
| 3. | Mixed breed | 10 | Complex type carcinoma | 12.3 | 3 | 6 |
| 4. | Mixed breed | 14 | Complex type carcinoma | 39.8 | 2 | 9 |
| 5. | Mixed breed | 6 | Cystic papillary carcinoma | 36.1 | 3 | 9 |
| 6. | Mixed breed | 9 | Adenocarcinoma tubular | 10.1 | 1 | 2 |
| 7. | Mixed breed | 12 | Carcinoma arising in benign mixed tumor | 24.8 | 2 | 4 |
| 8. | Mixed breed | 10 | Simple tubular carcinoma | 65.8 | 3 | 12 |
| 9. | Mixed breed | 10 | Complex type carcinoma | 56.3 | 3 | 9 |
| 10. | Mixed breed | 11 | Simple tubular carcinoma | 34.3 | 1 | 3 |
| 11. | Yorkshire Terrier | 10 | Complex Adenoma | 71.6 | 2 | 8 |
| 12. | Mixed Breed | 9 | Tubular Carcinoma | 27.5 | 3 | 6 |
| Grade 2 | ||||||
| No. Sample | Breed | Age (Years) | Histologic Classification | % Positive Cells | Intensity | IRS |
| 1. | Mixed breed | 9 | Tubulopapillary carcinoma | 15.3 | 1 | 2 |
| 2. | Poodle | 9 | Complex type carcinoma | 45.1 | 3 | 9 |
| 3. | Maltese | 10 | Mixed type carcinoma | 18.7 | 2 | 4 |
| 4. | Mixed breed | 11 | Solid carcinoma | 20 | 2 | 4 |
| 5. | Mixed breed | 10 | Tubulopapillary carcinoma | 19.2 | 3 | 6 |
| 6. | Labrador retriever | 12 | Tubulopapillary carcinoma | 31.2 | 2 | 6 |
| 7. | Mixed breed | 9 | Complex type carcinoma | 37.1 | 2 | 6 |
| 8. | Shih-tzu | 11 | Simple tubular carcinoma | 9.2 | 2 | 2 |
| 9. | Mixed Breed | 11 | Tubular Carcinoma | 65.8 | 2 | 8 |
| 10. | Cocker Spaniel | 10 | Tubular Carcinoma | 56.3 | 3 | 9 |
| 11. | Epagneul Breton | 10 | Tubulopapillary Carcinoma | 33.8 | 2 | 6 |
| 12. | Mixed breed | 6 | Mixed Type Carcinoma | 30.4 | 2 | 4 |
| Grade 3 | ||||||
| No. sample | Breed | Age (Years) | Histologic Classification | % Positive Cells | Intensity | IRS |
| 1. | Doberman pinscher | 13 | Simple tubular carcinoma | 40.7 | 1 | 3 |
| 2. | Maltese | 11 | Solid Carcinoma | 29.2 | 1 | 2 |
| 3. | Jack Russel Terrier | 13 | Simple Tubular Carcinoma | 27.3 | 3 | 6 |
| 4. | Greyhound | 9 | Simple Tubular Carcinoma | 6.7 | 1 | 1 |
| 5. | Siberian Husky | 11 | Comedocarcinoma | 0 | 0 | 0 |
| 6. | Cocker Spaniel | 6 | Complex Type Carcinoma | 3.4 | 1 | 1 |
| 7. | Mixed Breed | 9 | Carcinoma And Malignant Myoepithelioma | 13.4 | 2 | 4 |
| 8. | Mixed Breed | 11 | Complex Type Carcinoma | 43.1 | 2 | 6 |
| 9. | Shih Tzu | 9 | Complex Type Carcinoma | 0 | 0 | 0 |
| 10. | Mixed Breed | 12 | Mixed Type Carcinoma | 21.6 | 3 | 6 |
| 11. | Greyhound | 9 | Spindle Cell Carcinoma | 36.9 | 2 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sgadari, M.; Cacciola, N.A.; Power, K.; Martano, M.; Restucci, B. Sirtuin 1 Expression in Canine Mammary Tumors: A Pilot Study. Animals 2023, 13, 2609. https://doi.org/10.3390/ani13162609
Sgadari M, Cacciola NA, Power K, Martano M, Restucci B. Sirtuin 1 Expression in Canine Mammary Tumors: A Pilot Study. Animals. 2023; 13(16):2609. https://doi.org/10.3390/ani13162609
Chicago/Turabian StyleSgadari, Mariafrancesca, Nunzio Antonio Cacciola, Karen Power, Manuela Martano, and Brunella Restucci. 2023. "Sirtuin 1 Expression in Canine Mammary Tumors: A Pilot Study" Animals 13, no. 16: 2609. https://doi.org/10.3390/ani13162609
APA StyleSgadari, M., Cacciola, N. A., Power, K., Martano, M., & Restucci, B. (2023). Sirtuin 1 Expression in Canine Mammary Tumors: A Pilot Study. Animals, 13(16), 2609. https://doi.org/10.3390/ani13162609