
Non-Invasive Quantification of Faecal and Urine Reproductive Hormone Metabolites in the Naked Mole-Rat (Heterocephalus glaber)

 ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals and Housing
2.2. Sampling Schedule and Administration
2.3. Sample Storage and Steroid Extraction
2.4. Enzyme Immunoassays
2.5. Data Analysis
3. Results
3.1. EIA Validation
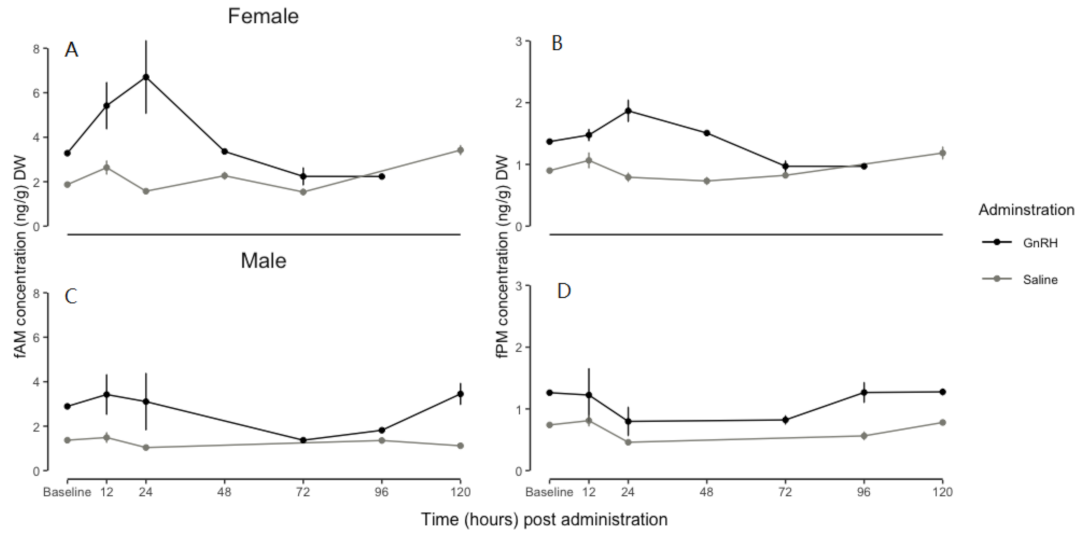
3.2. GnRH and Saline Administration: A Comparison of fAM Concentrations
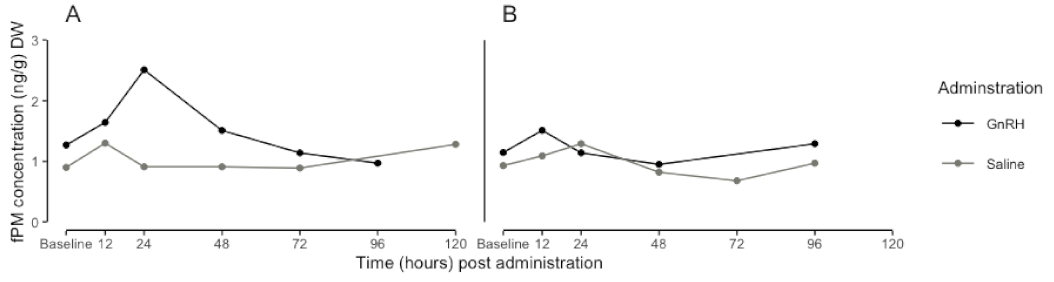
3.3. GnRH and Saline Administration: A Comparison of fPM Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tibbetts, E.A.; Pardo-Sanchez, J.; Weise, C. The Establishment and Maintenance of Dominance Hierarchies. Philos. Trans. R. Soc. B 2022, 377, 20200450. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.; Balshine, S. The Evolution of Cooperative Breeding in the African Cichlid Fish, Neolamprologus Pulcher. Biol. Rev. 2011, 86, 511–530. [Google Scholar] [CrossRef] [PubMed]
- Lukas, D.; Clutton-Brock, T. Cooperative Breeding and Monogamy in Mammalian Societies. Proc. R. Soc. B Biol. Sci. 2012, 279, 2151–2156. [Google Scholar] [CrossRef] [PubMed]
- Creel, S.; Creel, N.M. The African Wild Dog: Behavior, Ecology, and Conservation; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Nelson-Flower, M.J.; Hockey, P.A.R.; O’Ryan, C.; Raihani, N.J.; Du Plessis, M.A.; Ridley, A.R. Monogamous Dominant Pairs Monopolize Reproduction in the Cooperatively Breeding Pied Babbler. Behav. Ecol. 2011, 22, 559–565. [Google Scholar] [CrossRef]
- Greenwood, P.J. Mating Systems, Philopatry and Dispersal in Birds and Mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Bennett, N.C.; Faulkes, C.G.; Jarvis, J.U.M. Socially-Induced Infertility Incest Avoidance and the Monopoly of Reproduction in Cooperatively Breeding African Mole-Rats. Adv. Study Behav. 1999, 28, 75–113. [Google Scholar]
- Bell, M.B.V.; Cant, M.A.; Borgeaud, C.; Thavarajah, N.; Samson, J.; Clutton-Brock, T.H. Suppressing Subordinate Reproduction Provides Benefits to Dominants in Cooperative Societies of Meerkats. Nat. Commun. 2014, 5, 4499. [Google Scholar] [CrossRef]
- Van den Berghe, F.; Paris, M.C.J.; Sarnyai, Z.; Briggs, M.B.; Millar, R.P.; Ganswindt, A.; Paris, D.B.B.P. Social Rank does not Affect Sperm Quality in Male African Wild Dogs (Lycaon Pictus). Reprod. Fertil. Dev. 2019, 31, 875–887. [Google Scholar] [CrossRef]
- Kriegsfeld, L.J.; Mei, D.F.; Bentley, G.E.; Ubuka, T.; Mason, A.O.; Inoue, K.; Ukena, K.; Tsutsui, K.; Silver, R. Identification and Characterization of a Gonadotropin-Inhibitory System in the Brains of Mammals. Proc. Natl. Acad. Sci. USA 2006, 103, 2410–2415. [Google Scholar] [CrossRef]
- Clarke, I.J. Two Decades of Measuring GnRH Secretion. Reprod. Camb. Engl. Suppl. 2002, 59, 1–13. [Google Scholar]
- Jin, J.-M.; Yang, W.-X. Molecular Regulation of Hypothalamus-Pituitary-Gonads Axis in Males. Gene 2014, 551, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Wintermantel, T.M.; Campbell, R.E.; Porteous, R.; Bock, D.; Gröne, H.-J.; Todman, M.G.; Korach, K.S.; Greiner, E.; Pérez, C.A.; Schütz, G.; et al. Definition of Estrogen Receptor Pathway Critical for Estrogen Positive Feedback to Gonadotropin-Releasing Hormone Neurons and Fertility. Neuron 2006, 52, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Sower, S.A.; Freamat, M.; Kavanaugh, S.I. The Origins of the Vertebrate Hypothalamic-Pituitary-Gonadal (HPG) and Hypothalamic-Pituitary-Thyroid (HPT) Endocrine Systems: New Insights from Lampreys. Gen. Comp. Endocrinol. 2009, 161, 20–29. [Google Scholar] [CrossRef]
- Touma, C.; Sachser, N.; Möstl, E.; Palme, R. Effects of Sex and Time of Day on Metabolism and Excretion of Corticosterone in Urine and Feces of Mice. Gen. Comp. Endocrinol. 2003, 130, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Palme, R. Non-Invasive Measurement of Glucocorticoids: Advances and Problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Bennett, N.C.; Faulkes, C.G. African Mole-Rats: Ecology and Eusociality; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Jarvis, J.U. Eusociality in a Mammal: Cooperative Breeding in Naked Mole-Rat Colonies. Science 1981, 212, 571–573. [Google Scholar] [CrossRef]
- Wilson, E. The Insect Societies; Belknap Press; Harvard University Press: Cambridge, MA, USA, 1975; Volume 10, p. 548. [Google Scholar]
- Clutton-Brock, T. Social Evolution in Mammals. Science 2021, 373, eabc9699. [Google Scholar] [CrossRef]
- Reeve, H.; Sherman, P. Intracolonial Aggression and Nepotism by the Breeding Female Naked Mole-Rat. In The Biology of the Naked Mole-Rat; Sherman, P.W., Jarvis, J.U.M., Alexander, R.D., Eds.; Princeton University Press: Princeton, NJ, USA, 1991; pp. 337–357. [Google Scholar]
- O’Riain, M.J.; Jarvis, J.U.M.; Faulkes, C.G. A Dispersive Morph in the Naked Mole-Rat. Nature 1996, 380, 619–621. [Google Scholar] [CrossRef]
- Toor, I.; Edwards, P.D.; Kaka, N.; Whitney, R.; Ziolkowski, J.; Monks, D.A.; Holmes, M.M. Aggression and Motivation to Disperse in Eusocial Naked Mole-Rats, Heterocephalus Glaber. Anim. Behav. 2020, 168, 45–58. [Google Scholar] [CrossRef]
- Kayanja, F.I.B.; Jarvis, J.U.M. Histological Observations on the Ovary, Oviduct and Uterus of the Naked Mole-Rat. Z. Für Säugetierkunde 1971, 36, 114–121. [Google Scholar]
- Faulkes, C.G.; Abbott, D.H.; Jarvis, J.U.M. Social Suppression of Ovarian Cyclicity in Captive and Wild Colonies of Naked Mole-Rats, Heterocephalus Glaber. Reproduction 1990, 88, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Faulkes, C.G.; Abbott, D.H.; Jarvis, J.U.M.; Sherriff, F.E. LH Responses of Female Naked Mole-Rats, Heterocephalus Glaber, to Single and Multiple Doses of Exogenous GnRH. Reproduction 1990, 89, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Faykoo-Martinez, M.; Kalinowski, L.M.; Holmes, M.M. Neuroendocrine Regulation of Pubertal Suppression in the Naked Mole-Rat: What We Know and What Comes Next. Mol. Cell. Endocrinol. 2021, 534, 111360. [Google Scholar] [CrossRef]
- Bennett, N.C.; Jarvis, J.U.M. Coefficients of Digestibility and Nutritional Values of Geophytes and Tubers Eaten by Southern African Mole-Rats (Rodentia: Bathyergidae). J. Zool. 1995, 236, 189–198. [Google Scholar] [CrossRef]
- Fraňková, M.; Palme, R.; Frynta, D. Family Affairs and Experimental Male Replacement Affect Fecal Glucocorticoid Metabolites Levels in the Egyptian Spiny Mouse Acomys Cahirinus. Zool. Stud. 2012, 51, 277–287. [Google Scholar]
- Palme, R.; Touma, C.; Arias, N.; Dominchin, M.F.; Lepschy, M. Steroid Extraction: Get the Best out of Faecal Samples. Wien. Tierärztliche Monatsschrift 2013, 100, 238–246. [Google Scholar]
- Millspaugh, J.J.; Washburn, B.E. Use of Fecal Glucocorticoid Metabolite Measures in Conservation Biology Research: Considerations for Application and Interpretation. Gen. Comp. Endocrinol. 2004, 138, 189–199. [Google Scholar] [CrossRef]
- Dehnhard, M.; Fanson, K.; Frank, A.; Naidenko, S.V.; Vargas, A.; Jewgenow, K. Comparative Metabolism of Gestagens and Estrogens in the Four Lynx Species, the Eurasian (Lynx lynx), the Iberian (L. pardinus), the Canada Lynx (L. canadensis) and the Bobcat (L. rufus). Gen. Comp. Endocrinol. 2010, 167, 287–296. [Google Scholar] [CrossRef]
- Schwarzenberger, F.; Möstl, E.; Palme, R.; Bamberg, E. Faecal Steroid Analysis for Non-Invasive Monitor& of Reproductive Status in Farm, Wild and Zoo Animals. Anim. Reprod. Sci. 1996, 42, 15–526. [Google Scholar]
- Palme, R.; Mostl, E. Biotin–Streptavidin Enzyme Immunoassay for the Determination of Oestrogen and Androgens in Boar Faeces. In Advances of Steroid Analysis; Görög, E.S., Ed.; Akadémiai Kiadó: Budapest, Hungary, 1993; pp. 111–117. [Google Scholar]
- Ganswindt, A.; Heistermann, M.; Borragan, S.; Hodges, J.K. Assessment of Testicular Endocrine Function in Captive African Elephants by Measurement of Urinary and Fecal Androgens. Zoo Biol. 2002, 21, 27–36. [Google Scholar] [CrossRef]
- Folin, O.; Morris, J.L. On the Determination of Creatinine and Creatine in Urine. J. Biol. Chem. 1914, 17, 469–473. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bates, D.M.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Stoops, M.A.; Pairan, R.D.; Roth, T.L. Follicular, Endocrine and Behavioural Dynamics of the Indian Rhinoceros (Rhinoceros unicornis) Oestrous Cycle. Reproduction 2004, 128, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Williams-Kelly, K.S.; Robert, K.A.; Palme, R.; Fanson, K. V Validation of an Enzyme Immunoassay for the Non-Invasive Measurement of Faecal Androgen Metabolites in Spinifex Hopping Mice (Notomys Alexis). Aust. Mammal. 2022, 45, 192–198. [Google Scholar] [CrossRef]
- Auer, K.E.; Kußmaul, M.; Möstl, E.; Hohlbaum, K.; Rülicke, T.; Palme, R. Measurement of Fecal Testosterone Metabolites in mice: Replacement of Invasive Techniques. Animals 2020, 10, 165. [Google Scholar] [CrossRef]
- Wallace, K.M.E.; Hart, D.W.; Hagenah, N.; Ganswindt, A.; Bennett, N.C. A Comprehensive Profile of Reproductive Hormones in Eusocial Damaraland Mole-Rats (Fukomys damarensis). Gen. Comp. Endocrinol. 2023, 333, 114194. [Google Scholar] [CrossRef]
- Clarke, F.M.; Faulkes, C.G. Dominance and Queen Succession in Captive Colonies of the Eusocial Naked Mole-Rat, Heterocephalus Glaber. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1997, 264, 993–1000. [Google Scholar] [CrossRef]
- Clarke, F.M.; Faulkes, C.G. Hormonal and Behavioural Correlates of Male Dominance and Reproductive Status in Captive Colonies of the Naked Mole-Rat, Heterocephalus Glaber. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 1391–1399. [Google Scholar] [CrossRef]
- Faulkes, C.G.; Abbott, D.H.; Jarvis, J.U.M. Social Suppression of Reproduction in Male Naked Mole-Rats, Heterocephalus Glaber. Reproduction 1991, 91, 593–604. [Google Scholar] [CrossRef]
- Boomsma, J.J.; Gawne, R. Superorganismality and Caste Differentiation as Points of no Return: How the Major Evolutionary Transitions Were Lost in Translation. Biol. Rev. 2018, 93, 28–54. [Google Scholar] [CrossRef]
- Van der Walt, L.; Bennett, N.C.; Schoeman, S. Reproductive Suppression and Pituitary Sensitivity to Exogenous GnRH in the Highveld Mole-Rat (Cryptomys hottentotus pretoriae). J. Zool. 2001, 254, 177–184. [Google Scholar] [CrossRef]
- Spinks, A.C.; Bennett, N.C.; Faulkes, C.G.; Jarvis, J.U.M. Circulating LH Levels and the Response to Exogenous GnRH in the Common Mole-Rat: Implications for Reproductive Regulation in this Social, Seasonal Breeding Species. Horm. Behav. 2000, 37, 221–228. [Google Scholar] [CrossRef]
- Bennett, N.C.; Faulkes, C.G.; Spinks, A.C. LH Responses to Single Doses of Exogenous GnRH by Social Mashona Mole–Rats: A Continuum of Socially Induced Infertility in the Family Bathyergidae. Proc. R. Soc. London. Ser. B Biol. Sci. 1997, 264, 1001–1006. [Google Scholar] [CrossRef]
- Molteno, A.J.; Kallo, I.; Bennett, N.C.; King, J.A.; Coen, C.W. A Neuroanatomical and Neuroendocrinological Study into the Relationship between Social Status and the GnRH System in Cooperatively Breeding Female Damaraland Mole-Rats, Cryptomys Damarensis. Reproduction 2004, 127, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Buffenstein, R. Negligible Senescence in the Longest Living Rodent, the Naked Mole-Rat: Insights from a Successfully Aging Species. J. Comp. Physiol. B 2008, 178, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Blecher, A.S.; Bennett, N.C.; Medger, K.; Hagenah, N.; Ganswindt, A.; Oosthuizen, M.K. Effect of Colony Disruption and Social Isolation on Naked Mole-Rat Endocrine Correlates. Gen. Comp. Endocrinol. 2020, 113520. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ID | Testosterone EIA | Epiandrosterone EIA | Progesterone EIA | 5α Progesterone EIA |
|---|---|---|---|---|
| F1 | * 23 | * −24 | * 5 | * 12 |
| F2 | ^ −14 | ^ 28 | ^ 16 | ^ −0 |
| F3 | * −65 | * −67 | * 70 | * 72 |
| F4 | ^ 34 | ^ 70 | ^ 96 | ^ 327 |
| F5 | ^ 29 | ^ 55 | ^ 35 | ^ 110 |
| F6 | ^ −23 | ^ 6 | ^ 23 | ^ 15 |
| M1 | −89 | −84 | −87 | −89 |
| M2 | −15 | −22 | −16 | −6 |
| M4 | * 232 | * 80 | ‘ 136 | ‘ 8.5 |
| M5 | ^ 10 | ^ −17 | ^ 75 | ^ 78 |
| M6 | 23 | 35 | ‘ 1 | 31 |
| ID | Testosterone EIA | Epiandrosterone EIA | Progesterone EIA | 5α Progesterone EIA |
|---|---|---|---|---|
| F4 | * −2 | ‘ 41 | ‘ 16 | ‘ 34 |
| F5 | ‘ 641 | ‘ 298 | ‘ 153 | ‘ 109 |
| F6 | ^ 69 | ^ 123 | ^ 67 | ^ 98 |
| M4 | * 221 | 206 | ‘ 11 | * 41 |
| M5 | ^ 41 | ^ 163 | ^ 182 | ^ 73 |
| M6 | ‘ 39 | ‘ 42 | ‘ 23 | ‘ 13 |
| fAM | GnRH | Saline | ||||||
| Baseline (ng/g DW) | Time | Peak (ng/g DW) | % Change | Baseline (ng/g DW) | Time | Peak (ng/g DW) | % Change | |
| F1 | 0.45 | 25 | 1.09 | 142 | 1.31 | 12 | 1.91 | 46 |
| F2 | 1.33 | 10 | 2.77 | 108 | 1.57 | 16 | 1.68 | 7 |
| F3 | 0.95 | 11 | 1.43 | 51 | 1.11 | 12 | 1.16 | 5 |
| F4 | 2.25 | 25 | 3.17 | 41 | 1.85 | 14 | 1.44 | −22 |
| F5 | 2.93 | 27 | 11 | 275 | 1.89 | 5 | 3.34 | 77 |
| F6 | 0.11 | 4 | 0.24 | 123 | 0.21 | 2 | 0.27 | 29 |
| M1 | 2.79 | 9 | 4.35 | 56 | 3.01 | 6 | 5.23 | 74 |
| M2 | 3.04 | 15 | 4.45 | 46 | 3.19 | 17 | 5.6 | 76 |
| M3 | 1.43 | 14 | 3.7 | 159 | 1.25 | 14 | 1.36 | 9 |
| M4 | 1.82 | 20 | 5.59 | 208 | 1.26 | 10 | 2.08 | 65 |
| M5 | 0.16 | 4 | 0.4 | 150 | 0.26 | 17 | 1.18 | 363 |
| M6 | 1.64 | 19 | 2.32 | 42 | 1.45 | 27 | 2.98 | 106 |
| fPM | GnRH | Saline | ||||||
| Baseline (ng/g DW) | Time | Peak (ng/g DW) | % Change | Baseline (ng/g DW) | Time | Peak (ng/g DW) | % Change | |
| F1 | 0.86 | 25 | 0.93 | 8 | 1.18 | 18 | 1.43 | 21 |
| F2 | 1.81 | 22 | 1.56 | −14 | 1.05 | 16 | 1.81 | 72 |
| F3 | 1.15 | 1 | 1.51 | 32 | 0.93 | 20 | 1.29 | 39 |
| F4 | 1.81 | 28 | 2.42 | 34 | 1.57 | 14 | 1.65 | 5 |
| F5 | 1.27 | 26 | 2.51 | 98 | 0.9 | 5 | 1.3 | 44 |
| F6 | 0.26 | 4 | 0.51 | 98 | 0.26 | 0 | 0.44 | 69 |
| M1 | 1.35 | 9 | 1.37 | 1 | 1.35 | 10 | 1.83 | 36 |
| M2 | 1.9 | 9 | 1.68 | −12 | 1.83 | 17 | 2.42 | 32 |
| M3 | 0.74 | 14 | 1.01 | 36 | 0.68 | 7 | 0.46 | −32 |
| M4 | 1.17 | 10 | 1.74 | 48 | 0.66 | 10 | 0.98 | 48 |
| M5 | 0.14 | 4 | 0.23 | 70 | 0.32 | 21 | 0.55 | 75 |
| M6 | 1.42 | 13 | 1.6 | 13 | 1.14 | 6 | 1.66 | 46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majelantle, T.L.; Ganswindt, A.; Ganswindt, S.B.; Hagenah, N.; Hart, D.W.; Bennett, N.C. Non-Invasive Quantification of Faecal and Urine Reproductive Hormone Metabolites in the Naked Mole-Rat (Heterocephalus glaber). Animals 2023, 13, 3039. https://doi.org/10.3390/ani13193039
Majelantle TL, Ganswindt A, Ganswindt SB, Hagenah N, Hart DW, Bennett NC. Non-Invasive Quantification of Faecal and Urine Reproductive Hormone Metabolites in the Naked Mole-Rat (Heterocephalus glaber). Animals. 2023; 13(19):3039. https://doi.org/10.3390/ani13193039
Chicago/Turabian StyleMajelantle, Tshepiso Lesedi, Andre Ganswindt, Stefanie Birgit Ganswindt, Nicole Hagenah, Daniel William Hart, and Nigel Charles Bennett. 2023. "Non-Invasive Quantification of Faecal and Urine Reproductive Hormone Metabolites in the Naked Mole-Rat (Heterocephalus glaber)" Animals 13, no. 19: 3039. https://doi.org/10.3390/ani13193039
APA StyleMajelantle, T. L., Ganswindt, A., Ganswindt, S. B., Hagenah, N., Hart, D. W., & Bennett, N. C. (2023). Non-Invasive Quantification of Faecal and Urine Reproductive Hormone Metabolites in the Naked Mole-Rat (Heterocephalus glaber). Animals, 13(19), 3039. https://doi.org/10.3390/ani13193039