
The Effect of Oat Hay, Alfalfa Hay, and Their Combined Diets on the Morphology and Function of the Pancreas in Preweaning Yak Calves

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Animals
2.2. Feeding and Management
2.3. Sample Collection
2.4. Index Determination and Method
2.4.1. Growth and Development of the Pancreas
2.4.2. Morphological Development of the Pancreas
2.4.3. Functional Growth of the Pancreas
2.4.4. Metabolomics of the Pancreas
2.5. Data Processing and Statistic Analysis
3. Results
3.1. Effects of Different Forage Source Diets on the Growth and Development of the Pancreas in Preweaning Yak Calves
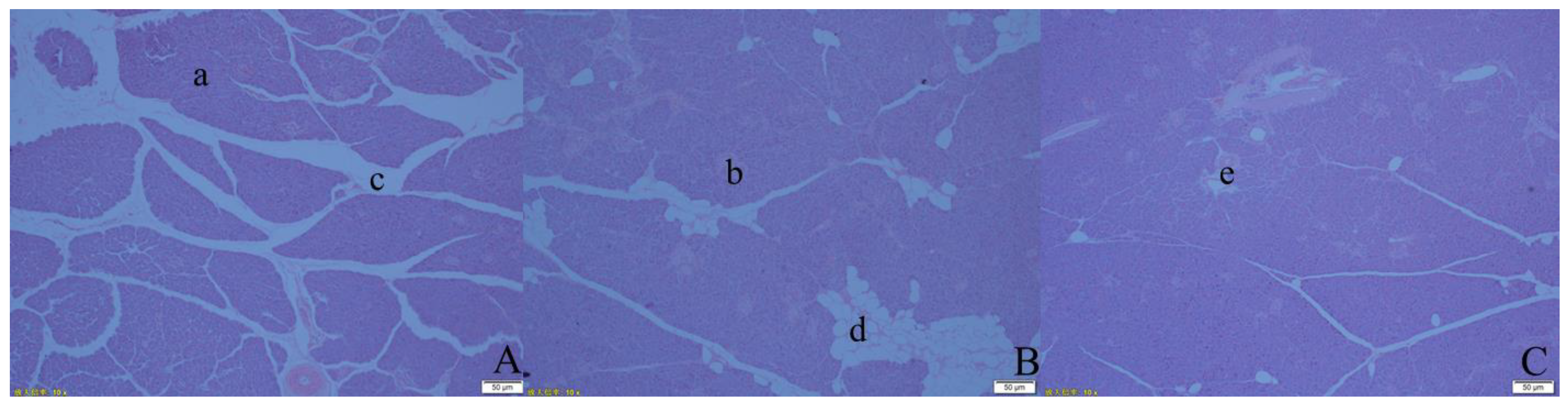
3.2. Effects of Different Forage Source Diets on the Morphological Development of the Pancreas in Preweaning Yak Calves
3.3. Effects of Different Forage Source Diets on the Development of Pancreatic Function in Preweaning Yak Calves
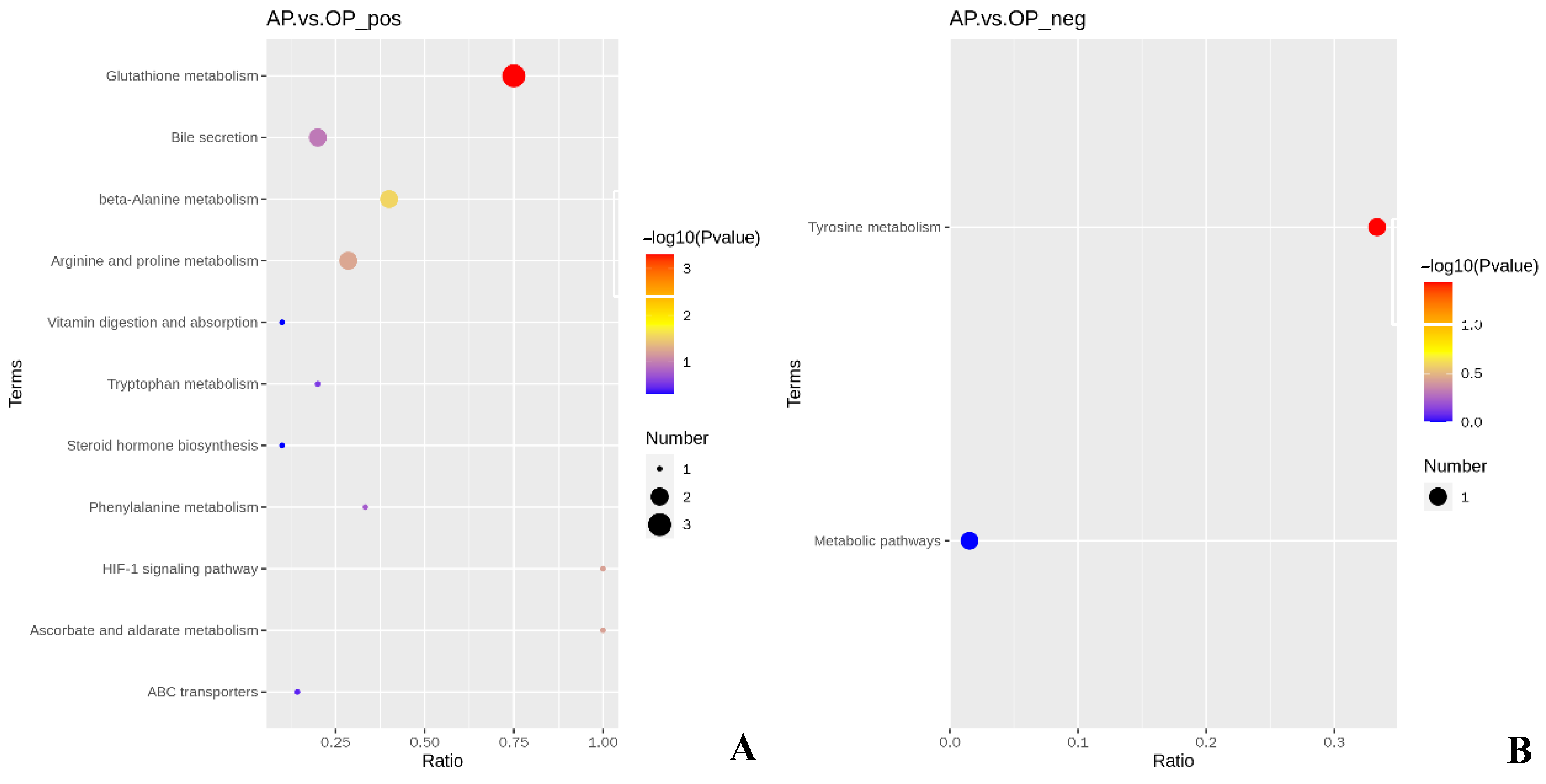
3.4. Effects of Different Forage Source Diets on the Metabolomics of the Pancreas of Preweaning Yak Calves
4. Discussion
4.1. Effects of Different Forage Source Diets on Growth Performance of Preweaning Yak Calves
4.2. Effects of Different Forage Source Diets on the Morphological Development of the Pancreas in Preweaning Yak Calves
4.3. Effects of Different Forage Source Diets on the Development of Pancreatic Function in Preweaning Yak Calves
4.4. Effects of Different Forage Source Diets on the Metabolomics of the Pancreas of Preweaning Yak Calves
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, K.; Shahzad, M.; Zhang, H.; Jiang, X.; Mehmood, K.; Zhao, X.D.; Li, J.K. Socio-economic burden of parasitic infections in yaks from 1984 to 2017 on Qinghai Tibetan Plateau of China-A review. Acta Trop. 2018, 183, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Y.M.; Shabir, M.; Kubrevi, S.S.; Dar, R.A.; Angmo, K.; Kanwar, M.S.; Bhat, R.A.; Muheet, M.; Parray, O.R. A study on biological rhythms of Himalayan yaks. Biol. Rhythm. Res. 2020, 51, 1087–1094. [Google Scholar] [CrossRef]
- Zhang, X.F.; Guan, J.Q.; Tu, Y.Q.; An, T.W.; Zhao, H.W.; Xiong, X.L.; Yang, P.G.; Wei, J.; LI, H.; Bai, Q.; et al. Progress of research on early weaning of yaks and its effect on calves. J. Grassl. Forage Sci. 2022, 2, 65–70. [Google Scholar]
- Kuan, J.Q.; Zhang, X.F.; Li, H.D.; Tan, X.; An, T.W.; Zhou, J.X.; Xie, R.Q.; Wu, W.S.; Zhao, H.W.; Luo, X.L. Early weaning and low altitude housing overwintering combined to promote rapid development of yak calves. J. Grassl. Forage Sci. 2020, 6, 71–74. [Google Scholar]
- Chai, S.M.; Yun, D.C.R.; Zhang, L.X.; Ge, S.L.M.; Zhao, M.D.; Yi, H.L.; Li, J.B.; Gong, J.C.R.; Ci, R.D.B.J.; Deng, B. Research on supplemental feeding and early weaning of calves in Gila golden yaks. China Dairy 2021, 12, 63–66. [Google Scholar]
- Li, Y.; Dai, D.W.; Yang, Y.K.; Wang, X.; Liu, S.J.; Hong, W.; Chai, S.T.; Wang, S.X. Influence of different concentrate to coarse rations on growth performance, apparent digestibility of nutrients, serum biochemical indexes and rumen fermentation parameters in late fattening yaks. Chin. J. Anim. Nutr. 2022, 34, 3056–3065. [Google Scholar]
- Contreras, G.F.E.; Muck, R.E.; Mertens, D.R.; Weimer, P.J. Microbial inoculant effects on silage and in vitro ruminal fermentation, and microbial biomass estimation for alfalfa, bmr corn, and corn silages. Anim. Feed. Sci. Technol. 2011, 163, 2–10. [Google Scholar] [CrossRef]
- Yang, C.; Hou, F.; Sun, Y.; Yuan, H.; Liu, Y.; Zhang, Y.; Chang, S. Oats hay supplementation to yak grazing alpine meadow improves carbon return to the soil of grassland ecosystem on the Qinghai-Tibet Plateau, China. Glob. Ecol. Conserv. 2020, 23, e01158. [Google Scholar] [CrossRef]
- Cui, Z.H.; Wu, S.R.; Li, J.L.; Yang, Q.E.; Chai, S.T.; Wang, L.; Wang, X.; Zhang, X.W.; Liu, S.J.; Yao, J.H. Effect of alfalfa hay and starter feeding intervention on gastrointestinal microbial community, growth and immune performance of yak calves. Front. Microbiol. 2020, 11, 994. [Google Scholar] [CrossRef]
- Hao, W.J.; Xia, H.Z.; Jiao, Y.; Cui, Z.H.; Liu, S.J.; Zhou, L.; Zhou, Y.Q. Effects of different GI and CBI combination diets on rumen and rectal bacterial compartments of yaks. China Anim. Husb. Vet. Med. 2022, 49, 1715–1726. [Google Scholar]
- Xia, H.Z.; Huang, W.Z.; Zhang, L.L.; Zhang, X.H.; Cui, Z.H.; Liu, S.J. Effect of different graded index oat hay-alfalfa hay combinations on rumen fermentation in vitro in yaks. J. Agric. Sci. Technol. 2021, 23, 191–201. [Google Scholar]
- Wu, D. Research on Pancreas Segmentation Method Based on Level Set; University of Electronic Science and Technology of China: Chengdu, China, 2022. [Google Scholar]
- Bharmal, S.H.; Pendharkar, S.A.; Singh, R.G.; Goodarzi, M.O.; Pandol, S.J.; Petrov, M.S. Relationship between circulating levels of pancreatic proteolytic enzymes and pancreatic hormones. Pancreatology 2017, 17, 876–883. [Google Scholar] [CrossRef]
- Cade, J.E.; Hanison, J. The pancreas. Anaesth. Intensive Care Med. 2017, 18, 527–531. [Google Scholar] [CrossRef]
- Yang, C.; Ahmad, A.A.; Bao, P.J.; Guo, X.; Wu, X.Y.; Liu, J.B.; Chu, M.; Liang, C.N.; Pei, J.; Long, R.J.; et al. Increasing dietary energy level improves growth performance and lipid metabolism through up-regulating lipogenic gene expression in yak (Bos grunniens). Anim. Feed. Sci. Technol. 2020, 263, 114455. [Google Scholar] [CrossRef]
- Wu, S.R.; Cui, Z.H.; Chen, X.D.; Zheng, L.X.; Ren, H.; Wang, D.D.; Yao, J.H. Diet-ruminal microbiome-host crosstalk contributes to differential effects of calf starter and alfalfa hay on rumen epithelial development and pancreatic α-amylase activity in yak calves. J. Dairy Sci. 2021, 104, 4326–4340. [Google Scholar] [CrossRef] [PubMed]
- Schneeman, B.O.; Richter, D. Changes in plasma and hepatic lipids, small intestinal histology and pancreatic enzyme activity due to aging and dietary fiber in rats. J. Nutr. 1993, 123, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Zou, X.P.; Li, X.Z.; Guo, G.; Ji, P.; Wang, Y.; Li, S.L.; Wang, Y.J.; Cao, Z.J. Substituting oat hay or maize silage for portion of alfalfa hay affects growth performance, ruminal fermentation, and nutrient digestibility of weaned calves. Asian-Australas. J. Anim. Sci. 2018, 31, 369. [Google Scholar] [CrossRef]
- Tang, Q.; Li, R.; Zhang, R.; Chen, X.C. Effect of intrauterine growth retardation on pancreatic development. Chin. J. Comp. Med. 2020, 30, 108–113. [Google Scholar]
- Twersky, Y.; Bank, S. Nutritional deficiencies in chronic pancreatitis. Gastroenterol. Clin. N. Am. 1989, 18, 543–565. [Google Scholar] [CrossRef]
- Svensson, A.M.; Jansson, L.; Hellerström, C. The volume and area of the capillaries in the endocrine and exocrine pancreas of the rat. Histochemistry 1988, 90, 43–46. [Google Scholar] [CrossRef]
- Calvert, R.; Schneeman, B.O.; Satchithanandam, S.; Cassidy, M.M.; Vahouny, G.V. Dietary fiber and intestinal adaptation: Effects on intestinal and pancreatic digestive enzyme activities. Am. J. Clin. Nutr. 1985, 41, 1249–1256. [Google Scholar] [CrossRef]
- Ribichini, E.; Stigliano, S.; Rossi, S.; Zaccari, P.; Sacchi, M.C.; Bruno, G.; Badiali, D.; Severi, C. Role of fibre in nutritional management of pancreatic diseases. Nutrients 2019, 11, 2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.M.; Su, Y.Y.; Jia, Z.T.; Liu, X.L.; Wang, C.Z.; Shi, Y.H.; Li, D.F.; Li, Z.T.; Zhu, X.Y. Combination effects of oat hay and alfalfa hay and their effects on dairy cows’ performance, milk quality, serum biochemical indices and apparent digestibility of nutrients. Chin. J. Anim. Nutr. 2021, 33, 6572–6583. [Google Scholar]
- Li, R. Comparative Study of Glucose Affecting Digestive Enzyme Secretion and MTOR and ISR Signaling Pathways in Pancreatic Tissues of Different Animals In Vitro; Inner Mongolia Agricultural University: Hohhot, China, 2021. [Google Scholar]
- Gemede, H.F.; Ratta, N. Antinutritional factors in plant foods: Potential health benefits and adverse effects. Int. J. Nutr. Food Sci. 2014, 3, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Subandi; Wijaya, M.; Sudarmo, T.P.B.; Suarsini, E. Saponin isolates from cucumber (Cucumis sativus L.) fruit mesocarp and their activity as pancreatic lipase inhibitor. AIP Conf. Proc. 2021, 1, 070016. [Google Scholar]
- Andersen, D.B.; Holst, J.J. Peptides in the regulation of glucagon secretion. Peptides 2022, 148, 170683. [Google Scholar] [CrossRef]
- Zhang, S. Cloning and Functional Analysis of Wheat Dehydroascorbate Reductase DHAR Gene; Northwest Agriculture & Forestry University: Xianyang, China, 2019. [Google Scholar]
- Zhang, H.; Wang, J.Y.; Li, L.X.; Chai, N.N.; Chen, Y.H.; Wu, F.X.; Zhang, W.H.; Wang, L.N.; Shi, S.; Zhang, L.; et al. Spermine and spermidine reversed age-related cardiac deterioration in rats. Oncotarget 2017, 8, 64793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parthasarathy, A.; Savka, M.A.; Hudson, A.O. The synthesis and role of β-alanine in plants. Front. Plant Sci. 2019, 10, 921. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Ge, F.; Yao, X.X.; Guo, X.; Bao, P.J.; Ma, X.M.; Wu, X.Y.; Chu, M.; Yan, P.; Liang, C.N. Microbiome and metabolomics reveal the effects of different feeding systems on the growth and ruminal development of Yaks. Front. Microbiol. 2021, 12, 1440. [Google Scholar] [CrossRef]
- Wu, X.J.; Cao, W.; Jia, G.; Zhao, H.; Chen, X.L.; Wu, C.M.; Tang, J.Y.; Wang, J.; Liu, G.M. New insights into the role of spermine in enhancing the antioxidant capacity of rat spleen and liver under oxidative stress. Anim. Nutr. 2017, 3, 85–90. [Google Scholar] [CrossRef]
- Wang, X.S.; Hu, M.X.; Guan, Q.X.; Men, L.H.; Liu, Z.Y. Metabolomics analysis reveals the renal protective effect of Panax ginseng CA Mey in type 1 diabetic rats. Chin. J. Nat. Med. 2022, 20, 378–386. [Google Scholar] [PubMed]
- Huang, C.F.; Chen, A.; Lin, S.Y.; Cheng, M.L.; Shiao, M.S.; Mao, T.Y. A metabolomics approach to investigate the proceedings of mitochondrial dysfunction in rats from prediabetes to diabetes. Saudi J. Biol. Sci. 2021, 28, 4762–4769. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Milk Replacer Powder | Concentrates | Oat Hay | Alfalfa Hay |
|---|---|---|---|---|
| Dry matter (DW) | 95.00 | 87.90 | 93.00 | 95.00 |
| Crude protein (CP) | 26.24 | 20.00 | 4.08 | 14.19 |
| Ether extract (EE) | 27.79 | 4.70 | 4.20 | 3.39 |
| Neutral detergent fiber (NDF) | - | 10.90 | 84.40 | 64.86 |
| Acid detergent fiber (ADF) | - | 4.10 | 45.18 | 46.13 |
| Calcium (Ca) | 2.50 | 0.80 | 1.05 | 1.55 |
| Phosphorus (P) | 1.40 | 0.45 | 0.15 | 0.74 |
| Items | OP 1 | AP 2 | AOP 3 | p-Value |
|---|---|---|---|---|
| Initial body weight (kg) | 35.8 ± 0.8 | 36.0 ± 1.6 | 37.6 ± 0.6 | 0.053 |
| Average daily gain (g/day) | 290.0 ± 3.1 | 293.3 ± 3.1 | 310.0 ± 10.0 | 0.095 |
| Final body weight (kg) | 70.6 ± 0.6 b | 71.2 ± 1.8 b | 74.8 ± 3.1 a | 0.017 |
| Dry matter intake (kg/day) | 1.00 | 1.00 | 1.00 | 1.000 |
| Pancreas weight (g) | 50.34 ± 2.01 b | 55.39 ± 2.67 ab | 61.44 ± 0.85 a | 0.007 |
| Pancreas index (%) | 0.07 ± 0.03 b | 0.08 ± 0.03 b | 0.09 ± 0.03 a | 0.022 |
| Items | OP 1 | AP 2 | AOP 3 | p-Value |
|---|---|---|---|---|
| Share of the total pancreatic endocrine component | 0.049 ± 0.001 b | 0.047 ± 0.003 b | 0.037 ± 0.002 a | <0.001 |
| Share of the total pancreatic exocrine portion | 0.87 ± 0.01 b | 0.91 ± 0.01 a | 0.93 ± 0.01 a | 0.009 |
| Area ratio of the exocrine and endocrine pancreas | 0.06 ± 0.00 b | 0.05 ± 0.00 b | 0.04 ± 0.00 a | <0.001 |
| Items | OP 1 | AP 2 | AOP 3 | p-Value |
|---|---|---|---|---|
| Pancreatic amylase | 33.8 ± 0.4 c | 35.9 ± 0.2 b | 36.9 ± 0.2 a | <0.001 |
| Pancreatic lipase | 114.7 ± 4.9 b | 121.4 ± 3.8 ab | 133.2 ± 3.5 a | 0.012 |
| Pancreatic protease | 163.3 ± 1.1 | 163.5 ± 2.3 | 166.7 ± 1.4 | 0.290 |
| Chymotrypsin | 124.6 ± 1.4 b | 129.3 ± 6.6 ab | 144.4 ± 6.3 a | 0.037 |
| Items | OP 1 | AP 2 | AOP 3 | p-Value |
|---|---|---|---|---|
| Glucagon | 0.30 ± 0.01 b | 0.34 ± 0.01 a | 0.35 ± 0.01 a | 0.002 |
| Insulin | 1.08 ± 0.02 b | 1.13 ± 0.04 ab | 1.20 ± 0.04 a | 0.033 |
| Pancreatic polypeptide | 2.68 ± 0.11 b | 3.02 ± 0.07 a | 3.25 ± 0.09 a | 0.001 |
| Growth inhibitors | 0.29 ± 0.00 c | 0.32 ± 0.01 b | 0.33 ± 0.00 a | <0.001 |
| No. | Metabolic Pathways | Differential Metabolites | p-Value |
|---|---|---|---|
| 1 | Glutathione metabolism | L-Ascorbate ↑ *; spermidine ↑; spermine ↑ | <0.001 |
| 2 | beta-Alanine metabolism | Spermidine ↑; spermine ↑ | 0.028 |
| 3 | Tyrosine metabolism | Dopaquinone ↑ | 0.037 |
| 4 | Arginine and proline metabolism | Spermidine ↑; spermine ↑ | 0.056 |
| 5 | Ascorbate and aldarate metabolism | L-Ascorbate ↑ | 0.061 |
| 6 | HIF-1 signaling pathway | L-Ascorbate ↑ | 0.061 |
| 7 | Bile secretion | Spermidine ↑; spermine ↑ | 0.111 |
| 8 | Phenylalanine metabolism | Hippuric acid ↓ | 0.173 |
| 9 | Tryptophan metabolism | 6-Hydroxymelatonin ↓ | 0.273 |
| 10 | ABC transporters | Spermidine ↑ | 0.364 |
| 11 | Steroid hormone biosynthesis | 5β-Androstane-3,17-dione ↓ | 0.481 |
| 12 | Vitamin digestion and absorption | L-Ascorbate ↑ | 0.481 |
| 13 | Metabolic pathways | Dopaquinone ↑ | 1.000 |
| No. | Metabolic Pathways | Differential Metabolites | p-Value |
|---|---|---|---|
| 1 | Arginine and proline metabolism | cis-4-Hydroxy-D-proline ↑ | 0.071 |
| 2 | Pantothenate and CoA biosynthesis | Pantetheine ↑ | 0.072 |
| 3 | Antifolate resistance | dUMP * ↑ | 0.072 |
| 4 | Bile secretion | Deoxycholic acid ↓ | 0.238 |
| 5 | Pyrimidine metabolism | dUMP ↑ | 0.268 |
| No. | Metabolic Pathways | Differential Metabolites | p-Value |
|---|---|---|---|
| 1 | Glutathione metabolism | Spermine ↓ * | 0.040 |
| 2 | Phenylalanine metabolism | Phenylacetylglycine ↓ | 0.048 |
| 3 | beta-Alanine metabolism | Spermine ↓ | 0.051 |
| 4 | Arginine and proline metabolism | Spermine ↓ | 0.071 |
| 5 | Tyrosine metabolism | Dopaquinone ↓ | 0.072 |
| 6 | Bile secretion | Spermine ↓ | 0.101 |
| 7 | Metabolic pathways | Spermine ↓ | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, Y.; Zhou, Y.; Liu, S.; Yang, D.; Li, J.; Sun, L.; Cui, Z. The Effect of Oat Hay, Alfalfa Hay, and Their Combined Diets on the Morphology and Function of the Pancreas in Preweaning Yak Calves. Animals 2023, 13, 293. https://doi.org/10.3390/ani13020293
Jiao Y, Zhou Y, Liu S, Yang D, Li J, Sun L, Cui Z. The Effect of Oat Hay, Alfalfa Hay, and Their Combined Diets on the Morphology and Function of the Pancreas in Preweaning Yak Calves. Animals. 2023; 13(2):293. https://doi.org/10.3390/ani13020293
Chicago/Turabian StyleJiao, Yang, Yanan Zhou, Shujie Liu, Deyu Yang, Jilan Li, Lu Sun, and Zhanhong Cui. 2023. "The Effect of Oat Hay, Alfalfa Hay, and Their Combined Diets on the Morphology and Function of the Pancreas in Preweaning Yak Calves" Animals 13, no. 2: 293. https://doi.org/10.3390/ani13020293
APA StyleJiao, Y., Zhou, Y., Liu, S., Yang, D., Li, J., Sun, L., & Cui, Z. (2023). The Effect of Oat Hay, Alfalfa Hay, and Their Combined Diets on the Morphology and Function of the Pancreas in Preweaning Yak Calves. Animals, 13(2), 293. https://doi.org/10.3390/ani13020293