
Isolation and Molecular Characterization of Corynebacterium pseudotuberculosis: Association with Proinflammatory Cytokines in Caseous Lymphadenitis Pyogranulomas
 ,
,  ,
,  ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Media and Culture Conditions
2.2. Identification of C. pseudotuberculosis
2.3. Antimicrobial Susceptibility
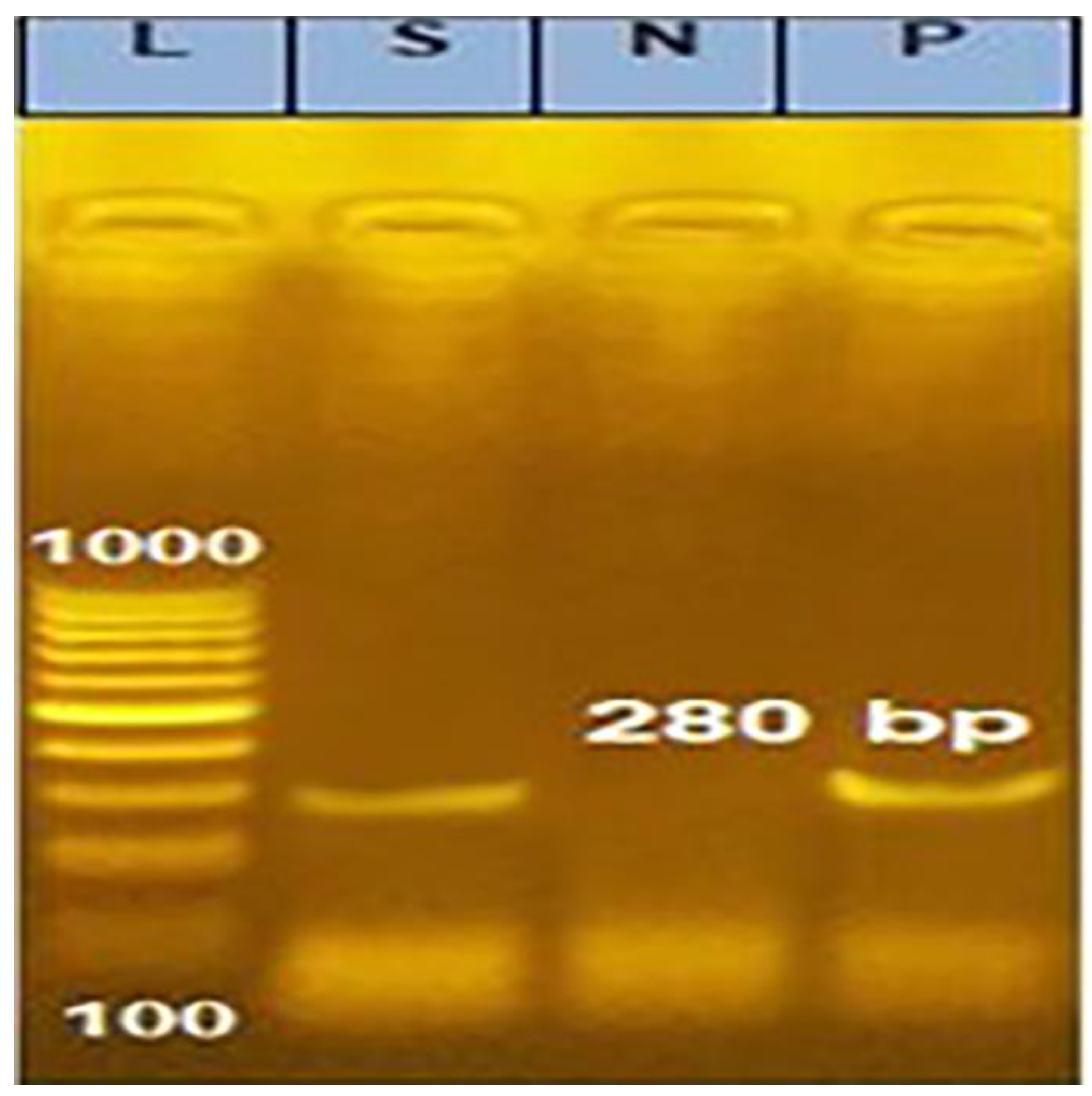
2.4. Integron1 Gene Cassette Detection:
2.4.1. DNA Extraction
2.4.2. Oligonucleotide Primer
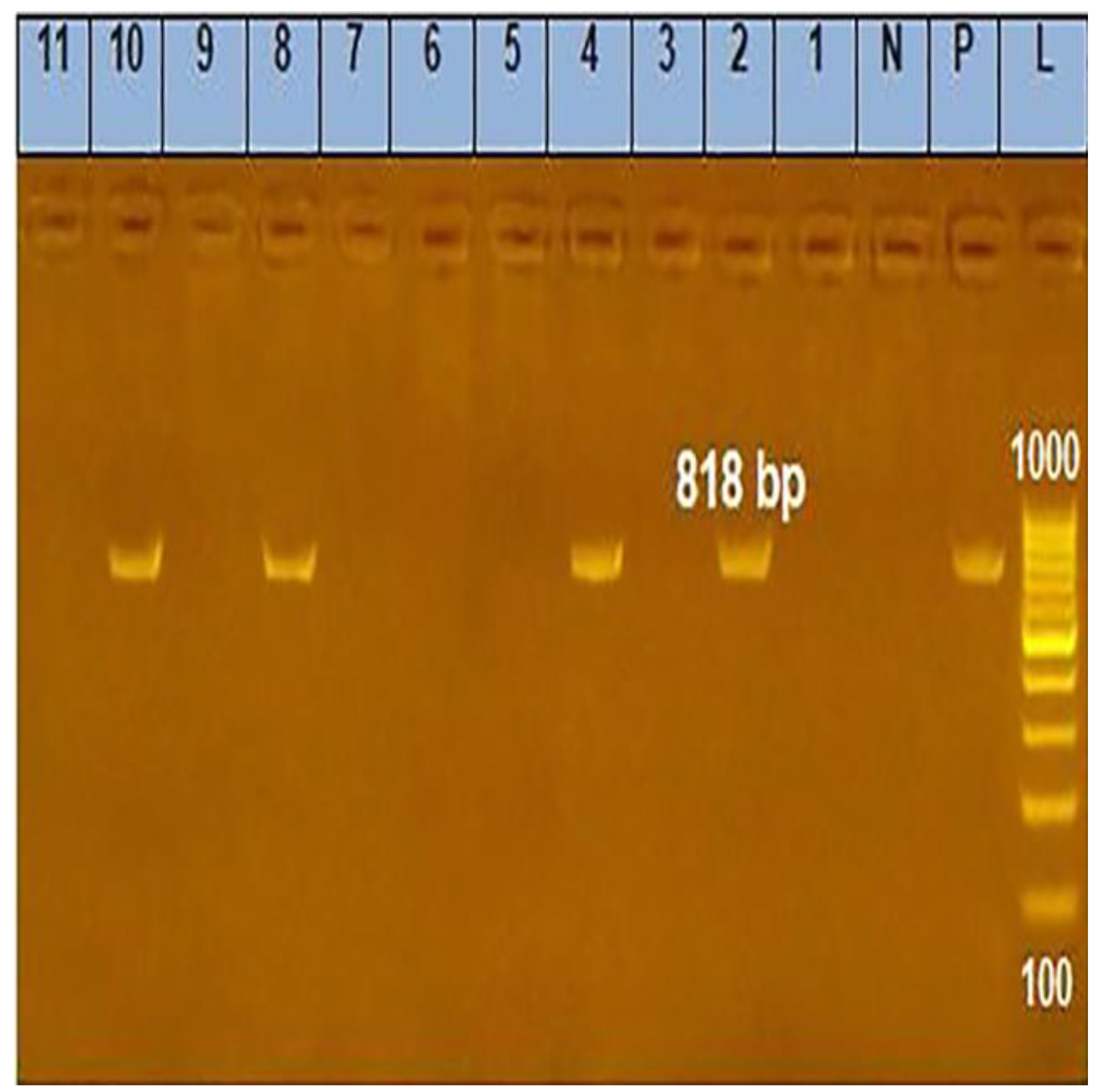
2.4.3. Analysis of the PCR Products
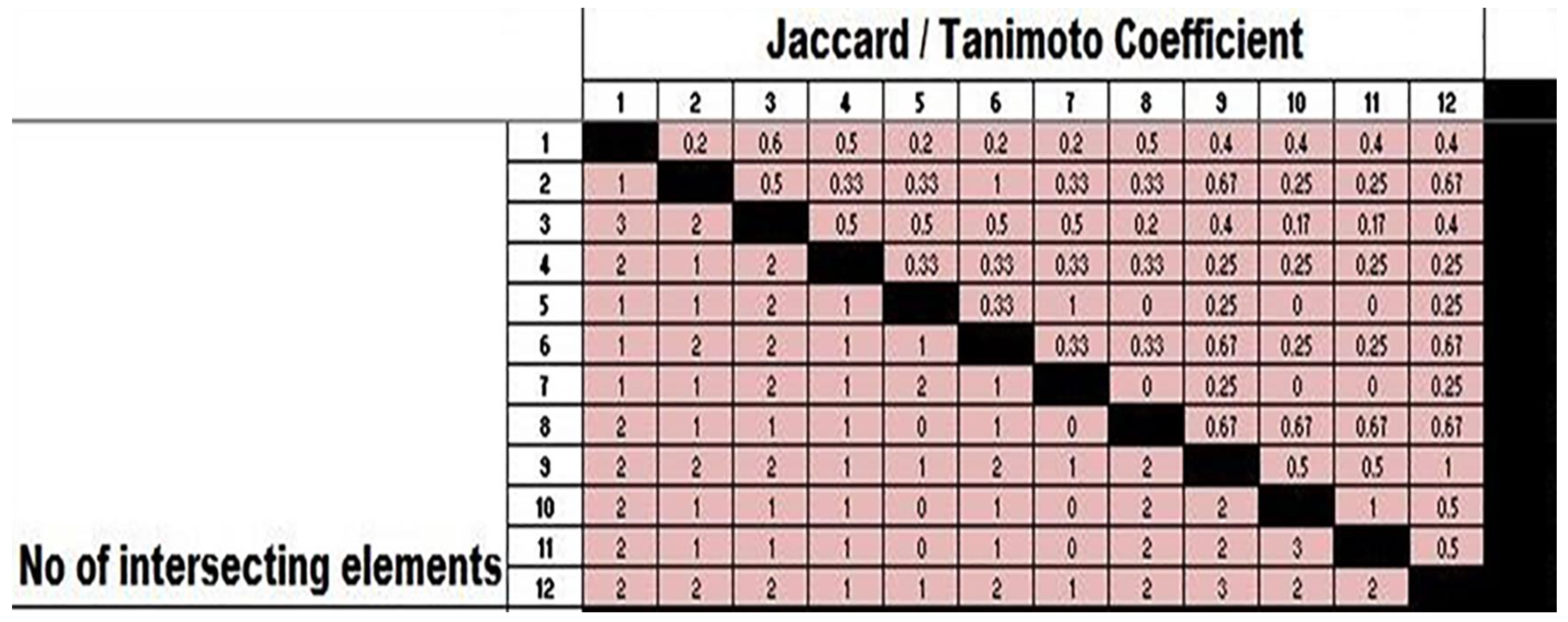
2.5. ERIC-PCR
- ERIC1, 5- ‘‘ATG TAA GCT CCT GGG GAT TCA C-3’’
- ERIC-2, 5- ‘‘AAG TAA GTG ACT GGG GTG AGG G-3’’
2.6. Sample Collection for Histopathological Examination
2.7. Evaluation of the Expression of NF-κB, IL1β, and TNF by Immunohistochemistry
3. Results
3.1. Bacteriological Identification and Antimicrobial Susceptibility

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Erythromycin | R | S | R | R | R | S | R | S | R | R | R | S |
| Doxycycline | S | S | S | S | S | S | S | S | S | S | S | S |
| Ciprofloxacin | S | S | S | S | S | S | S | S | S | S | S | S |
| Azithromycin | S | S | S | R | S | S | S | S | S | S | S | S |
| Cefaclor | R | S | R | S | R | S | R | R | R | R | R | R |
| Amoxicillin | R | S | R | S | R | S | R | R | R | R | R | R |
| Gentamicin | S | S | S | S | R | S | R | S | R | R | S | R |
| Kanamycin | S | S | S | S | S | S | S | S | R | R | S | S |
| Mupirocin | S | S | S | R | R | S | R | R | S | R | R | R |
| Clindamycin | S | S | S | S | S | S | R | S | S | S | S | S |
| Ampicillin | R | S | R | S | R | S | S | S | R | R | R | S |
| Number of Examined Isolates | Antibiotic Resistance Pattern | Resistance to Antimicrobial Classes |
|---|---|---|
| 3 | CF, AMX, AMP | 2 classes |
| 2 | MUP, AMX, GEN, CF and AMP | 4 classes |
| 1 | MUP, AMX, AMP, CF, K, GEN | 4 classes |
| 1 | GEN, AMX, AMP, MUP, CF | 4 classes |
| 1 | AMX, CF, MUP | 3 classes |
| 1 | K, AMX, AMP, CF, GEN | 3 classes |
| 1 | AMX, AMP, CF, MUP | 3 classes |
| 1 | AT, MUP | 2 classes |
| 1 | MUP, CF, E, GEN, AMX, CD | 6 classes |
3.2. Gross Pathology
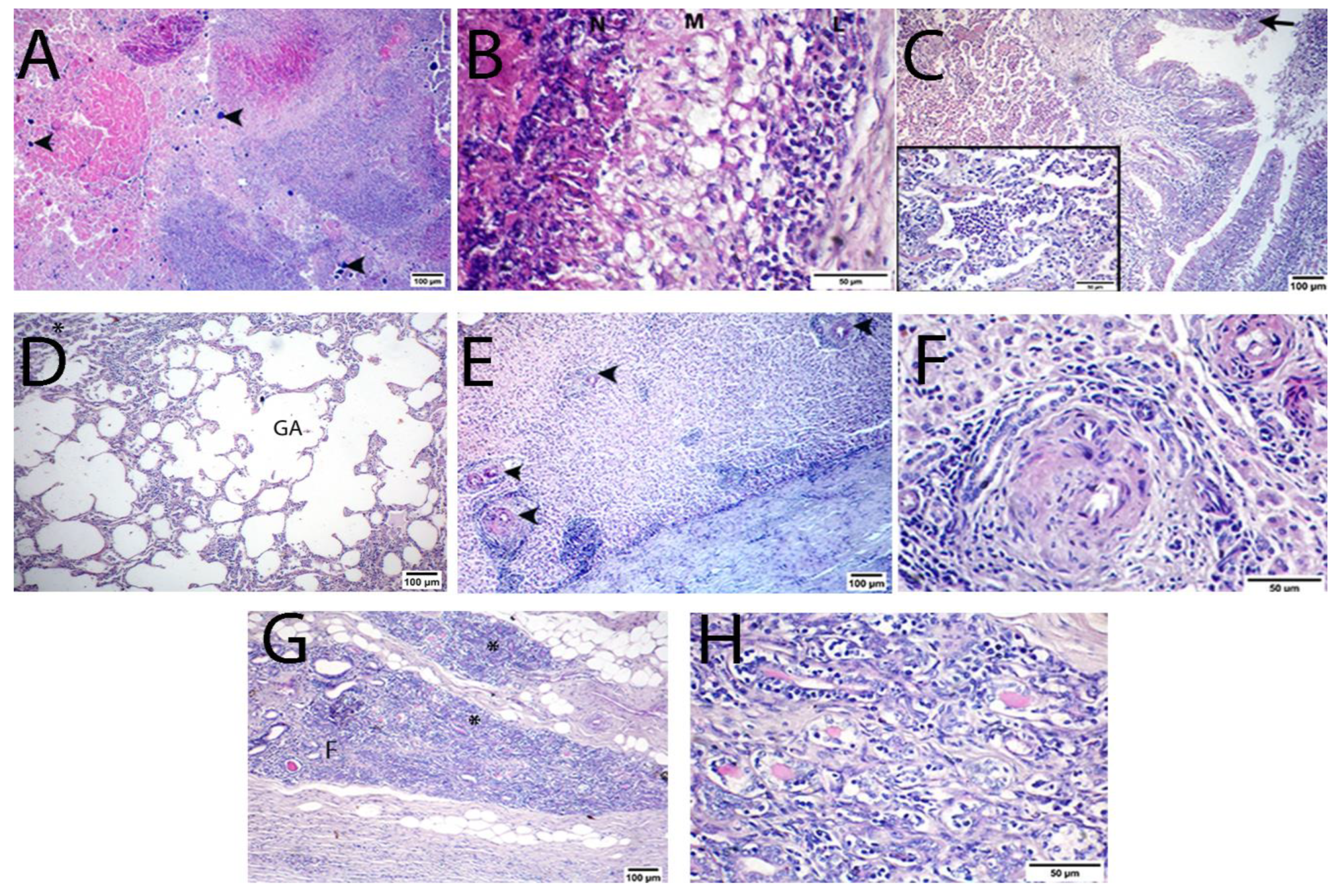
3.3. Histopathological Results
3.4. Immunohistochemical (IHC) Evaluation of NF-κB p65, IL1β and TNF Proteins
3.5. Pathogenicity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williamson, L.H. Caseous lymphadenitis in small ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2001, 17, 359–371. [Google Scholar] [CrossRef]
- Join-Lambert, O.F.; Ouache, M.; Canioni, D.; Beretti, J.-L.; Blanche, S.; Berche, P.; Kayal, S. Corynebacterium pseudotuberculosis necrotizing lymphadenitis in a twelve-year-old patient. Pediatr. Infect. Dis. J. 2006, 25, 848–851. [Google Scholar] [CrossRef]
- Connor, K.M.; Fontaine, M.C.; Rudge, K.; Baird, G.J.; Donachie, W. Molecular genotyping of multinational ovine and caprine Corynebacterium pseudotuberculosis isolates using pulsed-field gel electrophoresis. Vet. Res. 2007, 38, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohier, M.S.; Selim, S.; Bakry, M.; Elgabry, E.; Rehab, M.A. Comparison of protection against caseous lymphadenitis in sheep induced by local isolated strain of corynebacterium pseudotuberculosis by toxoid pld & toxoid pld With Bacterin Vaccine. Int. J. Dev. Res. 2017, 7, 15326–15334. [Google Scholar]
- Bastos, B.; Portela, R.D.; Dorella, F.; Ribeiro, D.; Seyffert, N.; Castro, T.; Miyoshi, A.; Oliveira, S.; Meyer, R.; Azevedo, V. Corynebacterium pseudotuberculosis: Immunological responses in animal models and zoonotic potential. J. Clin. Cell. Immunol. 2012, 4, 5. [Google Scholar] [CrossRef]
- Baird, G.; Fontaine, M. Corynebacterium pseudotuberculosis and its role in ovine caseous lymphadenitis. J. Comp. Pathol. 2007, 137, 179–210. [Google Scholar] [CrossRef] [PubMed]
- Pépin, M.; Fontaine, J.-J.; Pardon, P.; Marly, J.; Parodi, A. Histopathology of the early phase during experimental Corynebacterium pseudotuberculosis infection in lambs. Vet. Microbiol. 1991, 29, 123–134. [Google Scholar] [CrossRef]
- Dorella, F.A.; Pacheco, L.G.C.; Oliveira, S.C.; Miyoshi, A.; Azevedo, V. Corynebacterium pseudotuberculosis: Microbiology, biochemical properties, pathogenesis and molecular studies of virulence. Vet. Res. 2006, 37, 201–218. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, Z.; Jesse, F.; Saharee, A.; Jasni, S.; Yusoff, R.; Wahid, H. Clinio-pathological changes in goats challenged with Corynebacterium Peudotuberculosis and its exotoxin (PLD). Am. J. Anim. Vet. Sci. 2015, 10, 112–132. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, M.; Baird, G. Caseous lymphadenitis. Small Rumin. Res. 2008, 76, 42–48. [Google Scholar] [CrossRef]
- Ruiz-Bolivar, Z.; Carrascal-Camacho, A.K.; Neuque-Rico, M.C.; Gutierrez-Trivintilde, C.; Rodriguez-Bocanegra, M.X.; Poutou-Pintilde, R.A.; Mattar, S. Enterobacterial repetitive intergenic consensus-polymerase chain reaction (ERIC-PCR) fingerprinting reveals intra-serotype variations among circulating Listeria monocytogenes strains. Afr. J. Microbiol. Res. 2011, 5, 1586–1598. [Google Scholar]
- Anjay, A.K.; Agarwal, R.; Ramees, T.; Dubal, Z.; Kaushik, P.; Kumar, M.S.; Dudhe, N.; Milton, A.; Abhishek, B.; Shagufta, B. Molecular typing of Salmonella typhimurium and S. enteritidis serovars from diverse origin by ERIC-PCR. J. Pure Appl. Microbiol. 2015, 9, 2627–2634. [Google Scholar]
- Houf, K.; De Zutter, L.; Van Hoof, J.; Vandamme, P. Assessment of the genetic diversity among arcobacters isolated from poultry products by using two PCR-based typing methods. Appl. Environ. Microbiol. 2002, 68, 2172–2178. [Google Scholar] [CrossRef]
- Barakat, A.; Selim, S.; Atef, A.; Saber, M.; Nafie, E.; El-Edeeby, A. Two serotypes of Corynebacterium pseudotuberculosis isolated from different animal species. Rev. Sci. Et Tech. 1984, 3, 151–163. [Google Scholar] [CrossRef] [PubMed]
- de Sa Guimaraes, A.; Dorneles, E.M.S.; Andrade, G.I.; Lage, A.P.; Miyoshi, A.; Azevedo, V.; Gouveia, A.M.G.; Heinemann, M.B. Molecular characterization of Corynebacterium pseudotuberculosis isolates using ERIC-PCR. Vet. Microbiol. 2011, 153, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Christman, J.; Lancaster, L.; Blackwell, T. Nuclear factor k B: A pivotal role in the systemic inflammatory response syndrome and new target for therapy. Intensive Care Med. 1998, 24, 1131. [Google Scholar] [CrossRef]
- Al-Thomali, A.W.; Al-kuraishy, H.M.; Al-Gareeb, A.I.; Al-buhadiliy, A.K.; De Waard, M.; Sabatier, J.-M.; Khan Khalil, A.A.; Saad, H.M.; Batiha, G.E.-S. Role of Neuropilin 1 in COVID-19 Patients with Acute Ischemic Stroke. Biomedicines 2022, 10, 2032. [Google Scholar] [CrossRef]
- Vijayaraj, S.L.; Feltham, R.; Rashidi, M.; Frank, D.; Liu, Z.; Simpson, D.S.; Ebert, G.; Vince, A.; Herold, M.J.; Kueh, A. The ubiquitylation of IL-1β limits its cleavage by caspase-1 and targets it for proteasomal degradation. Nat. Commun. 2021, 12, 2713. [Google Scholar] [CrossRef]
- Babalghith, A.O.; Al-kuraishy, H.M.; Al-Gareeb, A.I.; De Waard, M.; Sabatier, J.-M.; Saad, H.M.; Batiha, G.E.-S. The Potential Role of Growth Differentiation Factor 15 in COVID-19: A Corollary Subjective Effect or Not? Diagnostics 2022, 12, 2051. [Google Scholar] [CrossRef]
- Moura-Costa, L.; Bahia, R.; Carminati, R.; Vale, V.; Paule, B.; Portela, R.; Freire, S.; Nascimento, I.; Schaer, R.; Barreto, L. Evaluation of the humoral and cellular immune response to different antigens of Corynebacterium pseudotuberculosis in Canindé goats and their potential protection against caseous lymphadenitis. Vet. Immunol. Immunopathol. 2008, 126, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Bernheimer, A.W.; Linder, R.; Avigad, L. Stepwise degradation of membrane sphingomyelin by corynebacterial phospholipases. Infect. Immun. 1980, 29, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, D. Association between Fatty Acids Profile of Corynebacterium Pseudotuberculosiswith Pathogenicity in Mice. University of Khartoum (Master thesis), 2015.
- Pacheco, L.G.; Pena, R.R.; Castro, T.L.; Dorella, F.A.; Bahia, R.C.; Carminati, R.; Frota, M.N.; Oliveira, S.C.; Meyer, R.; Alves, F.S. Multiplex PCR assay for identification of Corynebacterium pseudotuberculosis from pure cultures and for rapid detection of this pathogen in clinical samples. J. Med. Microbiol. 2007, 56, 480–486. [Google Scholar] [CrossRef] [Green Version]
- M100Ed32; Performance Standards for Antimicrobial Susceptibility Testing: Approved. CLSI: Berwyn, PA, USA, 2017. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 21 November 2022).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Kashif, J.; Buriro, R.; Memon, J.; Yaqoob, M.; Soomro, J.; Dongxue, D.; Jinhu, H.; Liping, W. Detection of class 1 and 2 integrons, β-lactamase genes and molecular characterization of sulfonamide resistance in Escherichia coli isolates recovered from poultry in China. Pak. Vet. J 2013, 33, 321–324. [Google Scholar]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D.; Saunders, N.; Owen, R. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 1989, 8, 151–156. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: Ch. 8. In Vitro Amplification of DNA by the Polymerase Chain Reaction; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Sambrook, H. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 1989. [Google Scholar]
- Yeh, F.; Yang, R.; Boyle, T. PopGene: Microsoft Window-Based Freeware for Population Genetic Analysis; Version 1.31; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier health sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Dabbs, D.J. Diagnostic Immunohistochemistry E-Book: Theranostic and Genomic Applications; Elsevier Health Sciences: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Connor, K.M.; Quirie, M.M.; Baird, G.; Donachie, W. Characterization of United Kingdom isolates of Corynebacterium pseudotuberculosis using pulsed-field gel electrophoresis. J. Clin. Microbiol. 2000, 38, 2633–2637. [Google Scholar] [CrossRef] [Green Version]
- Hommez, J.; Devriese, L.A.; Vaneechoutte, M.; Riegel, P.; Butaye, P.; Haesebrouck, F. Identification of nonlipophilic corynebacteria isolated from dairy cows with mastitis. J. Clin. Microbiol. 1999, 37, 954–957. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.U.; Dunn, L.; Ivanova, E.P. Evaluation of current molecular approaches for genotyping of Campylobacter jejuni strains. Foodborne Pathog. Dis. 2012, 9, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Mastroeni, P.; Maskell, D. Salmonella Infections: Clinical, Immunological and Molecular Aspects; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Rastogi, N.; Labrousse, V.; Goh, K.S. In vitro activities of fourteen antimicrobial agents against drug susceptible and resistant clinical isolates of Mycobacterium tuberculosis and comparative intracellular activities against the virulent H37Rv strain in human macrophages. Curr. Microbiol. 1996, 33, 167–175. [Google Scholar] [CrossRef]
- Padilla, E.; Llobet, E.; Doménech-Sánchez, A.; Martínez-Martínez, L.; Bengoechea, J.A.; Albertí, S. Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 2010, 54, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Foxman, B.; Zhang, L.; Koopman, J.S.; Manning, S.D.; Marrs, C.F. Choosing an appropriate bacterial typing technique for epidemiologic studies. Epidemiol. Perspect. Innov. 2005, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Raoult, D.; Fournier, P.-E. Bacterial strain typing in the genomic era. FEMS Microbiol. Rev. 2009, 33, 892–916. [Google Scholar] [CrossRef] [Green Version]
- O’reilly, K.; Green, L.; Malone, F.; Medley, G. Parameter estimation and simulations of a mathematical model of Corynebacterium pseudotuberculosis transmission in sheep. Prev. Vet. Med. 2008, 83, 242–259. [Google Scholar] [CrossRef]
- Al-Gaabary, M.H.; Osman, S.A.; Ahmed, M.S.; Oreiby, A.F. Abattoir survey on caseous lymphadenitis in sheep and goats in Tanta, Egypt. Small Rumin. Res. 2010, 94, 117–124. [Google Scholar] [CrossRef]
- Singh, R.; Singh, R.; Kamdi, B.; Kamari, S.; Singh, S.; Singh, S.; Chauhan, P.; Kumar, S. Pathology of Caseous Lymphadenitis in Slaughtered Goats Associated Infection with Corynebacterium Pseudotuberculosis. J. Anim. Res. 2018, 8, 497–500. [Google Scholar] [CrossRef]
- Abdulrahman, R.F.; Abdullah, M.A.; Kareem, K.H.; Najeeb, Z.D.; Hameed, H.M. Histopathological and molecular studies on caseous lymphadenitis in sheep and goats in duhok city, Iraq. Explor. Anim. Med. Res. 2020, 10, 134–140. [Google Scholar]
- Stefanska, I.; Gierynska, M.; Rzewuska, M.; Binek, M. Survival of Corynebacterium pseudotuberculosis within macrophages and induction of phagocytes death. Pol. J. Vet. Sci. 2010, 13, 143. [Google Scholar]
- Heymer, B.; Wirsing von König, C.; Finger, H.; Hof, H.; Emmerling, P. Histomorphology of experimental listeriosis. Infection 1988, 16, S106–S111. [Google Scholar] [CrossRef]
- Abebe, D.; Sisay Tessema, T. Determination of Corynebacterium pseudotuberculosis prevalence and antimicrobial susceptibility pattern of isolates from lymph nodes of sheep and goats at an organic export abattoir, Modjo, Ethiopia. Lett. Appl. Microbiol. 2015, 61, 469–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pépin, M.; Pittet, J.-C.; Olivier, M.; Gohin, I. Cellular composition of Corynebacterium pseudotuberculosis pyogranulomas in sheep. J. Leukoc. Biol. 1994, 56, 666–670. [Google Scholar] [CrossRef]
- Pepin, M.; Cannella, D.; Fontaine, J.-J.; Pittet, J.-C.; Le Pape, A. Ovine mononuclear phagocytes in situ: Identification by monoclonal antibodies and involvement in experimental pyogranulomas. J. Leukoc. Biol. 1992, 51, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, G.; Kumar, J.; Sisodia, S. Etio-pathological study of multiple hepatic abscesses in a goat. Indian J. Vet. Pathol. 2016, 40, 257–260. [Google Scholar] [CrossRef]
- Umer, M.; Jesse, F.F.A.; Saleh, W.M.M.; Chung, E.L.T.; Haron, A.W.; Saharee, A.A.; Lila, M.A.M.; Ariff, A.B.; Mohammad, K.; Sharif, A. Histopathological changes of reproductive organs of goats immunized with Corynebacterium pseudotuberculosis killed vaccine. Microb. Pathog. 2020, 149, 104539. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.; Jackson, H.; Brandon, M.; Meeusen, E. Lymphocyte subpopulations in pyogranulomas of caseous lymphadenitis. Clin. Exp. Immunol. 1991, 86, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, H.; Tian, S.; Yi, W.; Zhou, Y.; Yang, H.; Li, X.; Wu, B.; Li, X.; Wu, J. Critical roles of NLRP3 inflammasome in IL-1β secretion induced by Corynebacterium pseudotuberculosis in vitro. Mol. Immunol. 2019, 116, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Marrakchi, H.; Lanéelle, M.-A.; Daffé, M. Mycolic acids: Structures, biosynthesis, and beyond. Chem. Biol. 2014, 21, 67–85. [Google Scholar] [CrossRef] [Green Version]
- Omar, S.A.; Lawan, A.; Faez, F.; Yusuf, I.; Yusuf, A.; Hazilawati, B.H.; Mohd-Azmi, M.; Abdul, W.; Mohd, Z. Alterations in interleukin-1β and interleukin-6 in mice inoculated through the oral routes using graded doses of P. multocida type B: 2 and its lipopolysaccharide. Am. J. Anim. Vet. Sci. 2015, 10, 1–8. [Google Scholar]
- Othman, A.; Jesse, F.; Adza-Rina, M.; Ilyasu, Y.; Zamri-Saad, M.; Wahid, A.; Saharee, A.; Mohd-Azmi, M. Responses of inflammatory cytokines in non-pregnant boer does inoculated with Corynebacterium pseudotuberculosis via various routes. Res. Opin. Anim. Vet. Sci. 2014, 4, 597–600. [Google Scholar]
- Jesse, F.F.A.; Latif, N.-A.A.; Chung, E.L.T.; Adamu, L.; Sarah, S.A.; Zamri-Saad, M.; Haron, A.W.; Mohd, M.A. Cytokines (IL 1β and IL 6) Responses in Non-Pregnant Does Infected with Corynebacteriumpseudotuberculosis Following Intradermal Route of Infection in Chronic State. Int. J. Livest. Res. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.; Campos, M.; Snyder, M.; Chelak, B.; Haines, D. Local production of tumor necrosis factor-α in corynebacterial pulmonary lesions in sheep. Vet. Pathol. 1995, 32, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Barral, T.D.; Rebouças, M.F.; Loureiro, D.; Raynal, J.T.; Sousa, T.J.; Moura-Costa, L.F.; Azevedo, V.; Meyer, R.; Portela, R.W. Chemokine production induced by Corynebacterium pseudotuberculosis in a murine model. Braz. J. Microbiol. 2022, 53, 1019–1027. [Google Scholar] [CrossRef] [PubMed]








| Target Gene | Primers Sequences | Amplified Segment (bp) | Primary Denaturation | Amplification (35 Cycles) | Final Extension | ||
|---|---|---|---|---|---|---|---|
| Secondary Denaturation | Annealing | Extension | |||||
| Corynebacterium Pseudotuberculosis 16S rRNA | F-ACCGCACTTTAGTGTGTGTG R-TCTCTACGCCGATCTTGTAT | 816 | 94 °C 5 min | 94 °C 30 s | 58 °C 40 s | 72 °C 45 s | 72 °C 10 min |
| Integron1 gene cassette (Int1) | F-CCTCCCGCACGATGATC R-TCCACGCATCGTCAGGC | 280 | 94 °C 5 min | 94 °C 30 s | 50 °C 30 s | 72 °C 30 s | 72 °C 7 min |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torky, H.A.; Saad, H.M.; Khaliel, S.A.; Kassih, A.T.; Sabatier, J.-M.; Batiha, G.E.-S.; Hetta, H.F.; Elghazaly, E.M.; De Waard, M. Isolation and Molecular Characterization of Corynebacterium pseudotuberculosis: Association with Proinflammatory Cytokines in Caseous Lymphadenitis Pyogranulomas. Animals 2023, 13, 296. https://doi.org/10.3390/ani13020296
Torky HA, Saad HM, Khaliel SA, Kassih AT, Sabatier J-M, Batiha GE-S, Hetta HF, Elghazaly EM, De Waard M. Isolation and Molecular Characterization of Corynebacterium pseudotuberculosis: Association with Proinflammatory Cytokines in Caseous Lymphadenitis Pyogranulomas. Animals. 2023; 13(2):296. https://doi.org/10.3390/ani13020296
Chicago/Turabian StyleTorky, Helmy A., Hebatallah M. Saad, Samy A. Khaliel, Asmaa T. Kassih, Jean-Marc Sabatier, Gaber El-Saber Batiha, Helal F. Hetta, Eman M. Elghazaly, and Michel De Waard. 2023. "Isolation and Molecular Characterization of Corynebacterium pseudotuberculosis: Association with Proinflammatory Cytokines in Caseous Lymphadenitis Pyogranulomas" Animals 13, no. 2: 296. https://doi.org/10.3390/ani13020296
APA StyleTorky, H. A., Saad, H. M., Khaliel, S. A., Kassih, A. T., Sabatier, J. -M., Batiha, G. E. -S., Hetta, H. F., Elghazaly, E. M., & De Waard, M. (2023). Isolation and Molecular Characterization of Corynebacterium pseudotuberculosis: Association with Proinflammatory Cytokines in Caseous Lymphadenitis Pyogranulomas. Animals, 13(2), 296. https://doi.org/10.3390/ani13020296