

Simulating Hunting Effects on the Wild Boar Population and African Swine Fever Expansion Using Agent-Based Modeling
 , , , ,
, , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
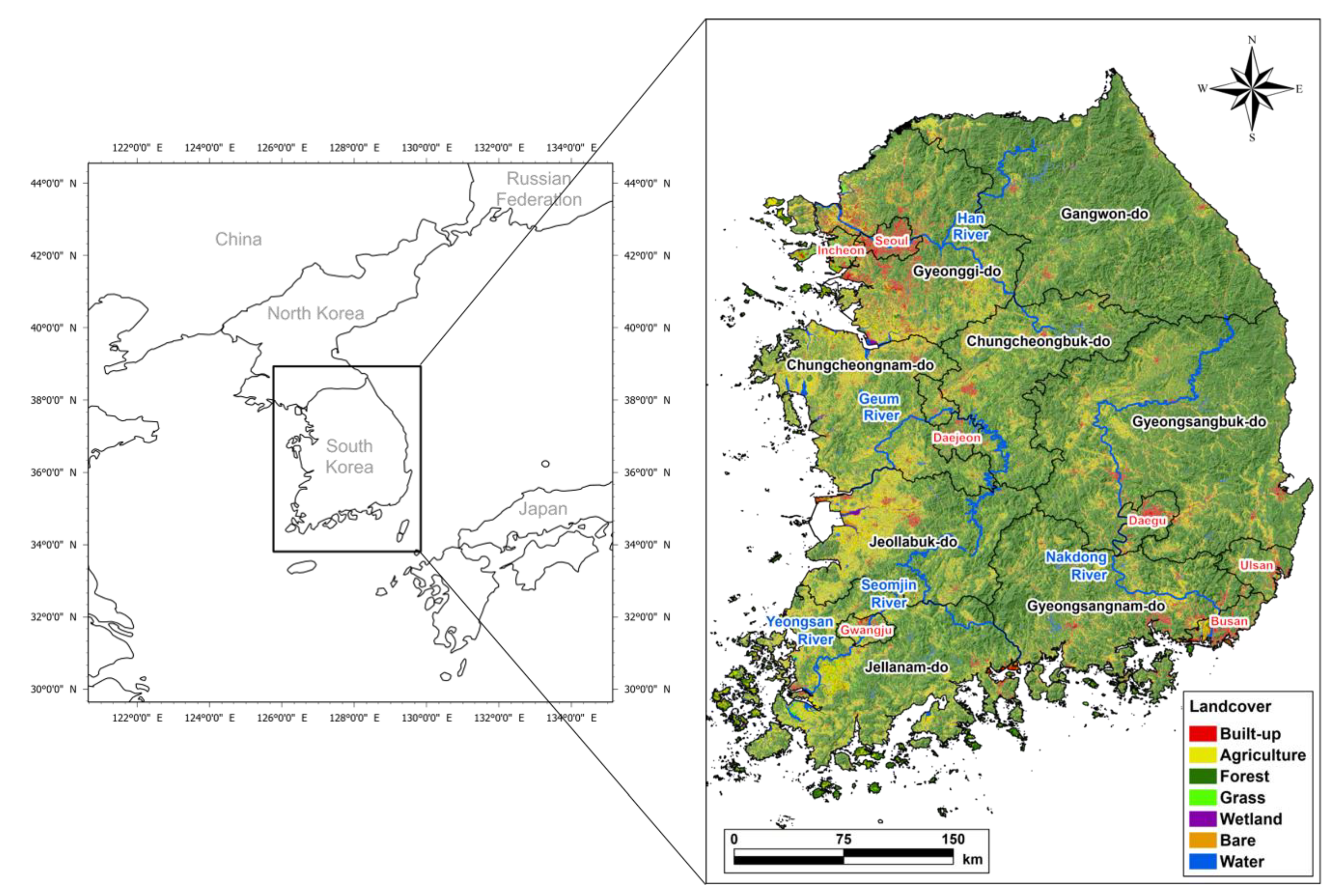
2.1. Study Area
2.2. Model Description
2.2.1. Background World
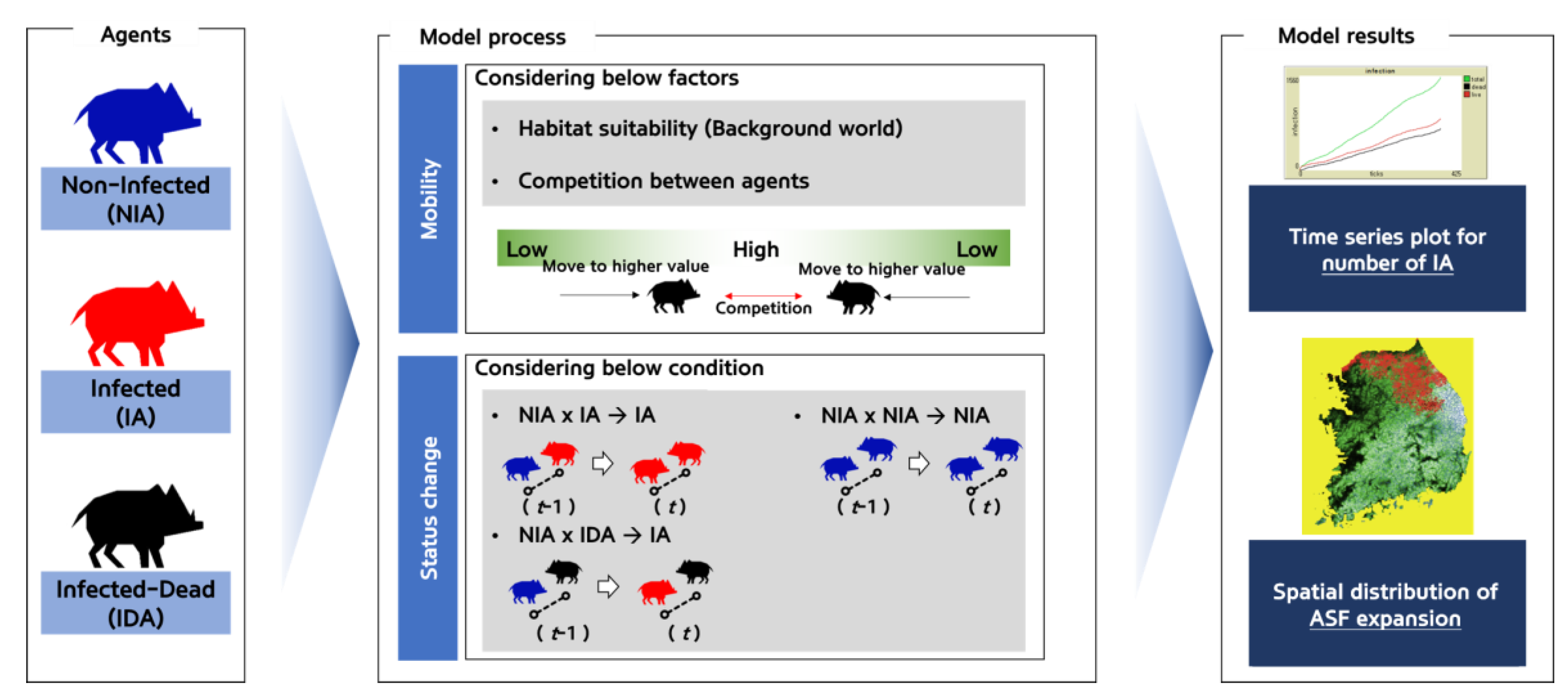
2.2.2. The Agents’ Status and the Status Conversion Process
2.2.3. The Lifecycle of Agents
2.2.4. Agents’ Mobility and ASF
2.2.5. Agent Culling Process
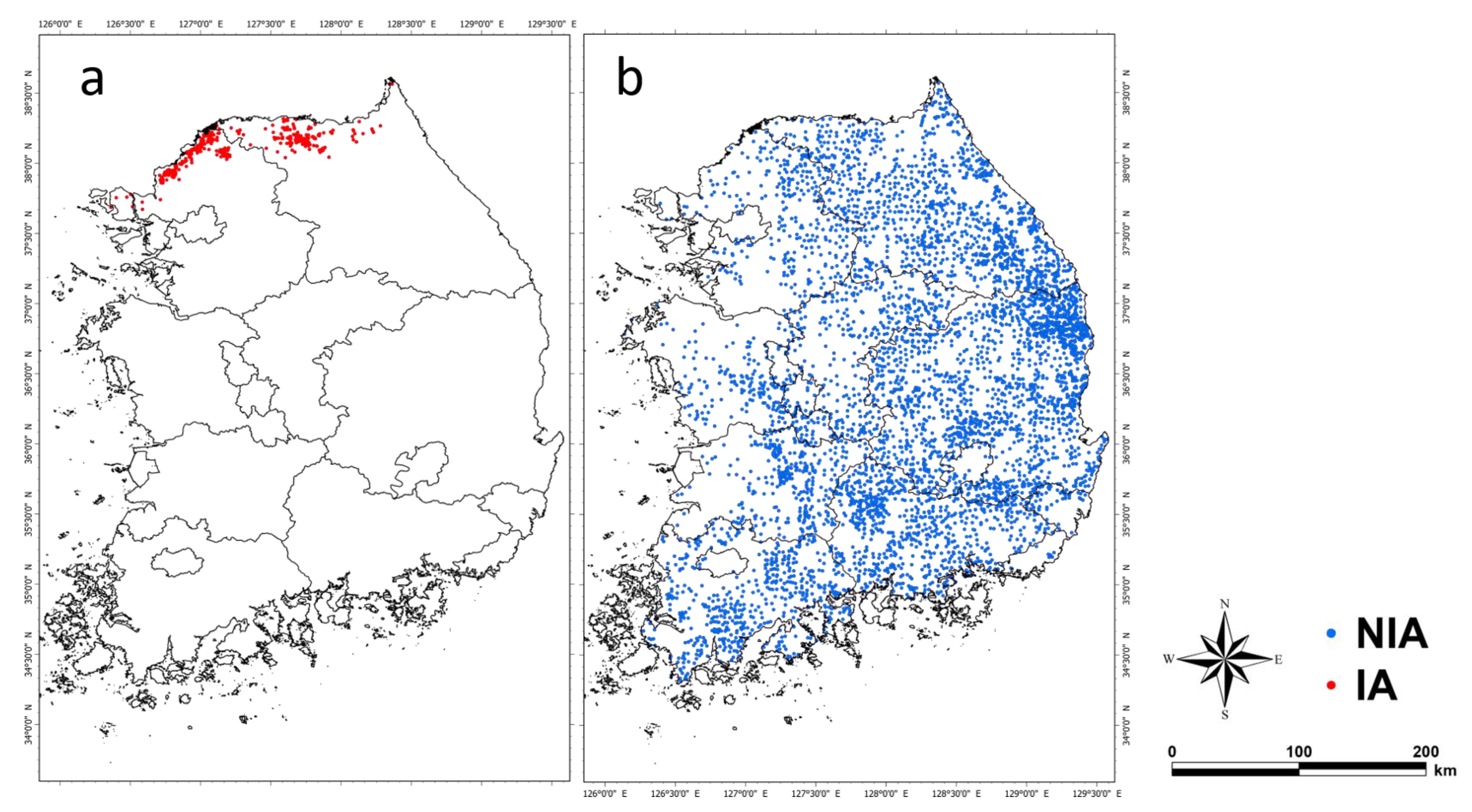
2.3. Model Initialization
2.4. Model Simulation and Validation
3. Results
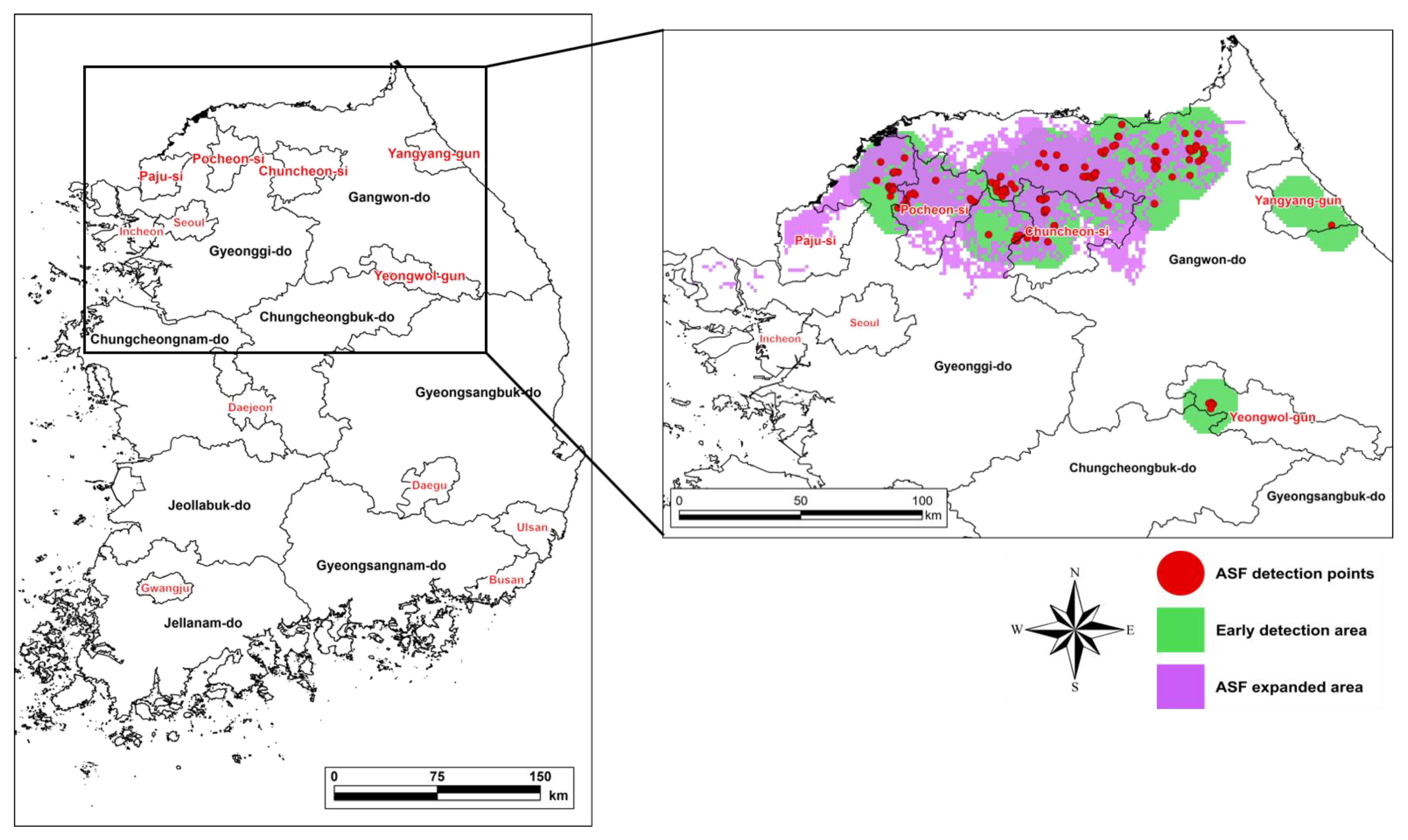
3.1. Model Validation
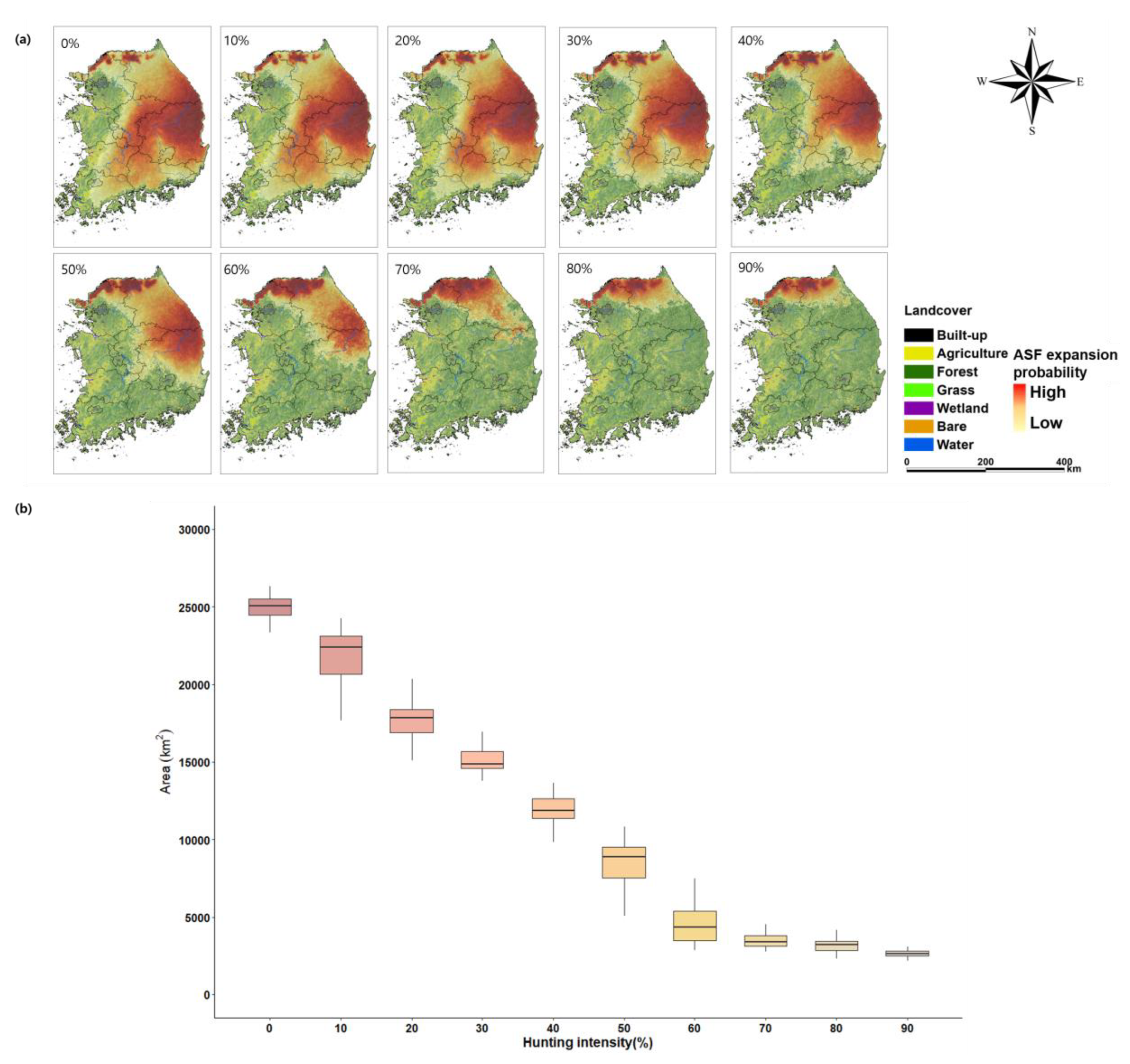
3.2. ASF Expansion in South Korea
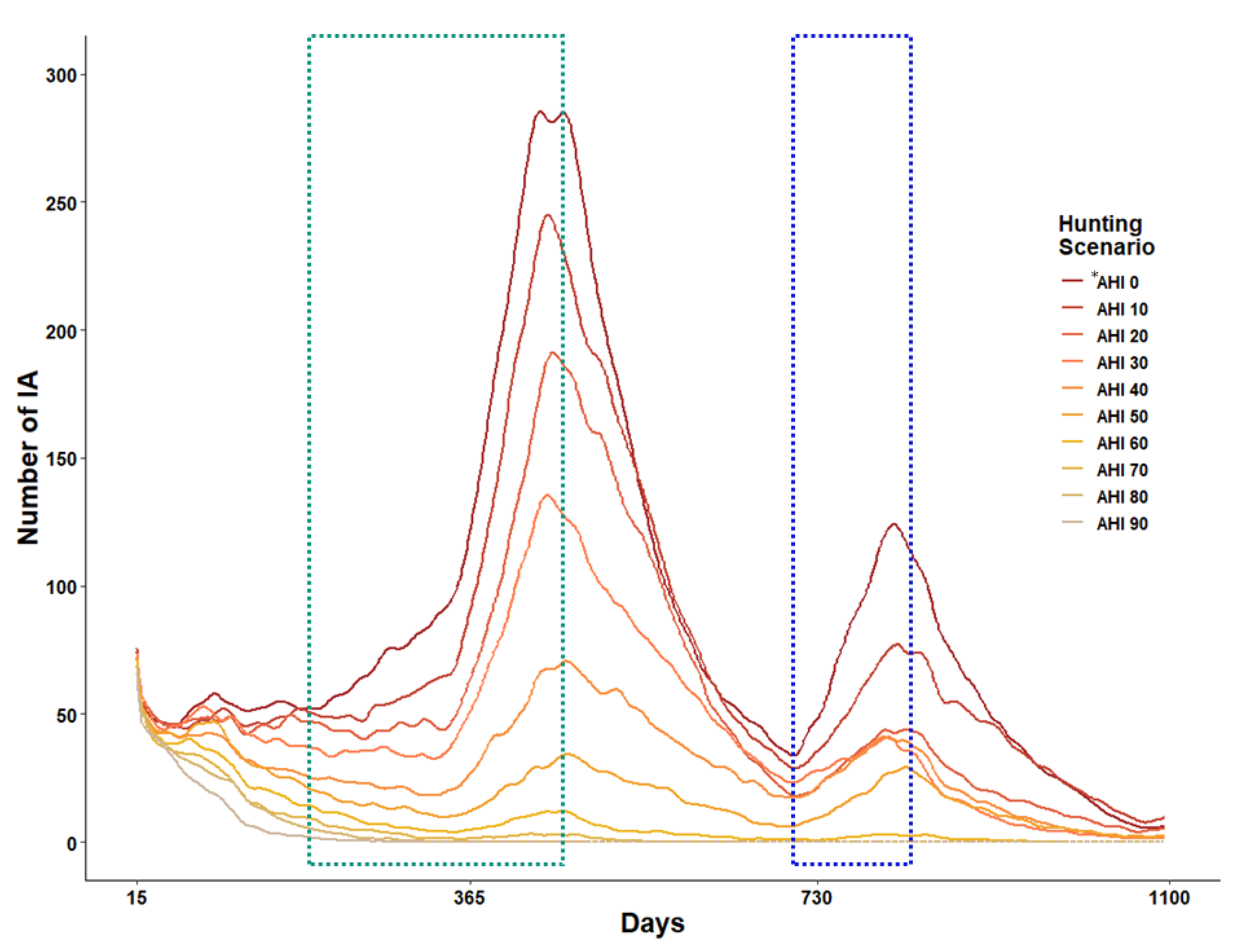
3.3. Number of ASF Infected Sounders
4. Discussion
4.1. Model Validation
4.2. The Spatial Distribution of ASF Expansion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez-Vizcaíno, J.M.; Martínez-López, B.; Martínez-Avilés, M.; Martins, C.; Boinas, F.; Vialc, L.; Michaud, V.; Jori, F.; Etter, E.; Albina, E.; et al. Scientific review on African swine fever. EFSA Support. Publ. 2009, 6, 5E. [Google Scholar] [CrossRef]
- Pepin, K.M.; Golnar, A.J.; Abdo, Z.; Podgórski, T. Ecological drivers of African swine fever virus persistence in wild boar populations: Insight for control. Ecol. Evol. 2020, 10, 2846–2859. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Smith, J.O.; Cross, P.C.; Briggs, C.J.; Daugherty, M.; Getz, W.M.; Latto, J.; Sanchez, M.S.; Smith, A.B.; Swei, A. Should we expect population thresholds for wildlife disease? Trends Ecol. Evol. 2005, 20, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, Y.J.; Lee, S.H.; Lee, E.H.; Park, J.J.; Park, K.H.; Sunwoo, S.Y.; Lee, J.G. A Study on Improvement Plan for Establishing a Farm Level ASF Control Strategies; Gyeonggi Research Institute: Suwon, Korea, 2020; pp. 1–133. (In Korean) [Google Scholar]
- Fischer, M.; Hühr, J.; Blome, S.; Conraths, F.J.; Probst, C. Stability of African swine fever virus in carcasses of domestic pigs and wild boar experimentally infected with the ASFV “Estonia 2014” isolate. Viruses 2020, 12, 1118. [Google Scholar] [CrossRef] [PubMed]
- Mebus, C.; Arias, M.; Pineda, J.M.; Tapiador, J.; House, C.; Sanchez-Vizcaino, J.M. Survival of several porcine viruses in different Spanish dry-cured meat products. Food Chem. 1997, 59, 555–559. [Google Scholar] [CrossRef]
- Agriculture, Forestry, and Livestock Quarantine Headquarters; ASF Prevalence in Farm Household in South Korea. Forestry and Livestock Quarantine Headquarters, Korea Disease Control and Prevention Agency. 2021. Available online: https://www.qia.go.kr/ (accessed on 28 April 2022).
- Ministry of Environment. Wild Boar Population Reduction Outcomes against African Swine Fever. Department of Biodiversity, Ministry of Environment. 2020. Available online: http://www.me.go.kr/home/web/board/read.do?pagerOffset=0&maxPageItems=10&maxIndexPages=10&searchKey=&searchValue=&menuId=286&orgCd=&boardMasterId=1&boardCategoryId=&boardId=1407580&decorator=/ (accessed on 14 May 2022).
- Jo, Y.S.; Gortázar, C. African swine fever in wild boar: Assessing interventions in South Korea. Transbound. Emerg. Dis. 2021, 68, 2878–2889. [Google Scholar] [CrossRef]
- Ministry of Environment. ASF Prevalence in South Korea. Department of Biodiversity, Ministry of Environment. 2022. Available online: https://www.me.go.kr/home/web/policy_data/read.do?pagerOffset=0&maxPageItems=10&maxIndexPages=10&searchKey=title&searchValue=ASF&menuId=10259&orgCd=&condition.toInpYmd=null&condition.fromInpYmd=null&condition.deleteYn=N&condition.deptNm=null&seq=7815/ (accessed on 30 April 2022).
- Bosch, J.; Iglesias, I.; Muñoz, M.J.; De la Torre, A. A cartographic tool for managing African swine fever in Eurasia: Mapping wild boar distribution based on the quality of available habitats. Transbound. Emerg. Dis. 2017, 64, 1720–1733. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.Y.; Lee, Y.S.; Park, C.H. Home-range of wild boar, Sus scrofa living in the Jirisan National Park, Korea. J. Ecol. Environ. 2006, 29, 253–257. [Google Scholar]
- Kurz, J.C.; Marchinton, R.L. Radiotelemetry studies of feral hogs in South Carolina. J. Wildl. Manag. 1972, 1240–1248. [Google Scholar] [CrossRef]
- Choi, T.Y.; Choi, H.M. Wildlife Tracks Guide; Dolbegae: Paju, Korea, 2007; pp. 234–242. [Google Scholar]
- Lee, S.M.; Lee, E.J. Diet of the wild boar (Sus scrofa): Implications for management in forest-agricultural and urban environments in South Korea. PeerJ. 2009, 7, e7835. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Kim, W.M.; Kim, K.T.; Jeong, S.G.; Kim, Y.J.; Lee, K.J.; Kim, H.G.; Park, C.; Song, W.K. A Study on the Connectivity Modeling Considering the Habitat and Movement Characteristics of Wild Boars (Sus scrofa). J. Korea Soc. Environ. Restor. Technol. 2022, 25, 33–47. (In Korean) [Google Scholar]
- Massei, G.; Genov, P.V.; Staines, B.W.; Gorman, M.L. Factors influencing home range and activity of wild boar (Sus scrofa) in a Mediterranean coastal area. J. Zool. 1997, 242, 411–423. [Google Scholar] [CrossRef]
- Fritzemeier, J.; Teuffert, J.; Greiser-Wilke, I.; Staubach, C.; Schlüter, H.; Moennig, V. Epidemiology of classical swine fever in Germany in the 1990s. Vet. Microbiol. 2000, 77, 29–41. [Google Scholar] [CrossRef]
- Podgórski, T.; Baś, G.; Jędrzejewska, B.; Sönnichsen, L.; Śnieżko, S.; Jędrzejewski, W.; Okarma, H. Spatiotemporal behavioral plasticity of wild boar (Sus scrofa) under contrasting conditions of human pressure: Primeval forest and metropolitan area. J. Mammal. 2013, 94, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Held, S.; Baumgartner, J.; Kilbride, A.; Byrne, R.W.; Mendl, M. Foraging behaviour in domestic pigs (Sus scrofa): Remembering and prioritizing food sites of different value. Anim. Cogn. 2005, 8, 114–121. [Google Scholar] [CrossRef]
- Singer, F.J.; Otto, D.K.; Tipton, A.R.; Hable, C.P. Home ranges, movements, and habitat use of European wild boar in Tennessee. J. Wildl. Manag. 1981, 45, 343–353. [Google Scholar] [CrossRef]
- Bertolotto, E.S. Behavioural Ecology of Wild Board (Sus scrofa) in an Apennine Environment. Ph.D. Thesis, University of Sassari, Sassari, Italy, 2010. [Google Scholar]
- Kim, C.H.; Kang, J.H.; Kim, M. Status and Development of National Ecosystem Survey in Korea. J. Environ. Impact Assess. 2013, 22, 725–738. (In Korean) [Google Scholar] [CrossRef] [Green Version]
- National Institute of Ecology. Guidelines for the 5th National Natural Environment Survey; National Institute of Ecology: Seocheon, Korea, 2019; p. 282. (In Korean) [Google Scholar]
- Belsare, A.V.; Gompper, M.E.; Keller, B.; Sumners, J.; Hansen, L.; Millspaugh, J.J. An agent-based framework for improving wildlife disease surveillance: A case study of chronic wasting disease in Missouri white-tailed deer. Ecol. Model. 2020, 417, 108919. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, H.; Gao, X.; Xiao, J.; Wang, H. African swine fever emerging in China: Distribution characteristics and high-risk areas. Prev. Vet. Med. 2020, 175, 104861. [Google Scholar] [CrossRef]
- Hayes, B.H.; Andraud, M.; Salazar, L.G.; Rose, N.; Vergne, T. Mechanistic modelling of African swine fever: A systematic review. Prev. Vet. Med. 2021, 191, 105358. [Google Scholar] [CrossRef] [PubMed]
- Viboud, C.; Boëlle, P.Y.; Carrat, F.; Valleron, A.J.; Flahault, A. Prediction of the spread of influenza epidemics by the method of analogues. Am. J. Epidemiol. 2003, 158, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Adde, A.; Roucou, P.; Mangeas, M.; Ardillon, V.; Desenclos, J.C.; Rousset, D.; Girod, R.; Briolant, S.; Quenel, P.; Flamand, C. Predicting dengue fever outbreaks in French Guiana using climate indicators. PLoS Negl. Trop. Dis. 2006, 10, e0004681. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, X.; Kok, S.Y.; Rajarethinam, J.; Liang, S.; Yap, G.; Chong, C.S.; Lee, K.S.; Tan, S.S.; Chin, C.K. Three-month real-time dengue forecast models: An early warning system for outbreak alerts and policy decision support in Singapore. Environ. Health Perspect. 2006, 124, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Vargas Amado, M.E.; Grütter, R.; Fischer, C.; Suter, S.; Bernstein, A. Free-ranging wild boar (Sus scrofa) in Switzerland: Casual observations and model-based projections during open and closed season for hunting. Schweiz. Arch. Tierheilkd. 2020, 162, 365–376. [Google Scholar] [CrossRef]
- Seo, C.; Park, C. Wild Boar (Sus scrofa coreanus Heude) Habitat Modeling Using GIS and Logistic Regression. J. GIS Assoc. Korea 2000, 8, 85–99. (In Korean) [Google Scholar]
- Cadenas-Fernández, E.; Ito, S.; Aguilar-Vega, C.; Sánchez-Vizcaíno, J.M.; Bosch, J. The Role of the Wild Boar Spreading African Swine Fever Virus in Asia: Another Underestimated Problem. Front. Vet. Sci. 2022, 9, 844209. [Google Scholar] [CrossRef]
- Gharakhanlou, N.M.; Hooshangi, N.; Helbich, M. A spatial agent-based model to assess the spread of malaria in relation to anti-malaria interventions in Southeast Iran. ISPRS Int. J. Geo-Inf. 2020, 9, 549. [Google Scholar] [CrossRef]
- Ko, C.; Cho, W.; Hwang, B.; Ko, D.W.; Kang, W. The Analysis of African Swine Fever Disease Spread using Agent-Based Model. J. Korean Cadast. Inf. Assoc. 2021, 23, 87–98. (In Korean) [Google Scholar]
- Gervasi, V.; Gubertì, V. Combining hunting and intensive carcass removal to eradicate African swine fever from wild boar populations. Prev. Vet. Med. 2022, 203, 105633. [Google Scholar] [CrossRef] [PubMed]
- Korea Meteorological Administration Home Page. Available online: https://www.weather.go.kr/w/obs-climate/climate/korea-climate/korea-char.do (accessed on 7 November 2022).
- DeAngelis, D.L. Individual-Based Models and Approaches in Ecology: Populations, Communities, and Ecosystems; Chapman and Hall: New York, NY, USA, 1992. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Thurfjell, H.; Spong, G.; Ericsson, G. Effects of hunting on wild boar Sus scrofa behaviour. Wildl. Biol. 2013, 19, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Morelle, K.; Podgórski, T.; Prévot, C.; Keuling, O.; Lehaire, F.; Lejeune, P. Towards understanding wild boar Sus scrofa movement: A synthetic movement ecology approach. Mammal Rev. 2015, 45, 15–29. [Google Scholar] [CrossRef]
- Lim, J.S.; Vergne, T.; Pak, S.I.; Kim, E. Modelling the Spatial Distribution of ASF-Positive wild boar carcasses in South Korea using 2019–2020 national surveillance data. Animals 2021, 11, 1208. [Google Scholar] [CrossRef]
- Sodeikat, G.; Pohlmeyer, K. Impact of drive hunts on daytime resting site areas of wild boar family groups (Sus scrofa L.). Wildl. Biol. Pract. 2007, 3, 28–38. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, C.M. The effect of hunter-wild boar interactions and landscape heterogeneity on wild boar population size: A simulation study. Ecol. Model. 2022, 464, 109847. [Google Scholar] [CrossRef]
- More, S.; Miranda, M.A.; Bicout, D.; Bøtner, A.; Butterworth, A.; Calistri, P.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Michel, V. African swine fever in wild boar. EFSA J. 2018, 16, e05344. [Google Scholar] [PubMed] [Green Version]
- Croft, S.; Franzetti, B.; Gill, R.; Massei, G. Too many wild boar? Modelling fertility control and culling to reduce wild boar numbers in isolated populations. PloS ONE 2020, 15, e0238429. [Google Scholar] [CrossRef] [PubMed]
- Benavides, J.; Walsh, P.D.; Meyers, L.A.; Raymond, M.; Caillaud, D. Transmission of infectious diseases en route to habitat hotspots. PLoS ONE 2012, 7, e31290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukielka, E.; Barasona, J.A.; Cowie, C.E.; Drewe, J.A.; Gortazar, C.; Cotarelo, I.; Vicente, J. Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps. Prev. Vet. Med. 2013, 112, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.; Oļševskis, E.; Staubach, C.; Lamberga, K.; Seržants, M.; Cvetkova, S.; Conraths, F.J.; Sauter-Louis, C. Epidemiological evaluation of Latvian control measures for African swine fever in wild boar on the basis of surveillance data. Sci. Rep. 2019, 9, 4189. [Google Scholar] [PubMed] [Green Version]
- Kouidere, A.; Balatif, O.; Rachik, M. Analysis and optimal control of a mathematical modeling of the spread of African swine fever virus with a case study of South Korea and cost-effectiveness. Chaos Solit. Fractals 2021, 146, 110867. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, C.; Cho, W.; Hwang, B.; Chang, B.; Kang, W.; Ko, D.W. Simulating Hunting Effects on the Wild Boar Population and African Swine Fever Expansion Using Agent-Based Modeling. Animals 2023, 13, 298. https://doi.org/10.3390/ani13020298
Ko C, Cho W, Hwang B, Chang B, Kang W, Ko DW. Simulating Hunting Effects on the Wild Boar Population and African Swine Fever Expansion Using Agent-Based Modeling. Animals. 2023; 13(2):298. https://doi.org/10.3390/ani13020298
Chicago/Turabian StyleKo, Chanwoo, Wonhee Cho, Byungmook Hwang, Byungwoo Chang, Wanmo Kang, and Dongwook W. Ko. 2023. "Simulating Hunting Effects on the Wild Boar Population and African Swine Fever Expansion Using Agent-Based Modeling" Animals 13, no. 2: 298. https://doi.org/10.3390/ani13020298
APA StyleKo, C., Cho, W., Hwang, B., Chang, B., Kang, W., & Ko, D. W. (2023). Simulating Hunting Effects on the Wild Boar Population and African Swine Fever Expansion Using Agent-Based Modeling. Animals, 13(2), 298. https://doi.org/10.3390/ani13020298