
Effects of Different-Syllable Aggressive Calls on Food Intake and Gene Expression in Vespertilio sinensis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Species
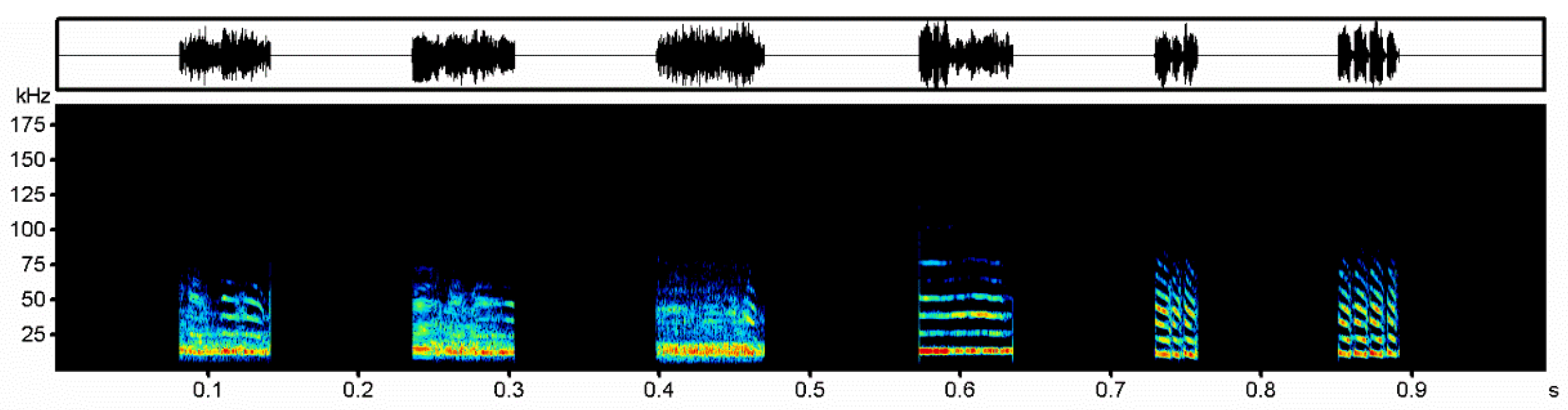
2.2. Recording of Aggressive Calls and Editing of Playback Files
2.3. Playback Experiment
2.4. Differential Gene Expression Analysis
2.5. GO/KEGG Enrichment Analysis for DEGs
3. Results
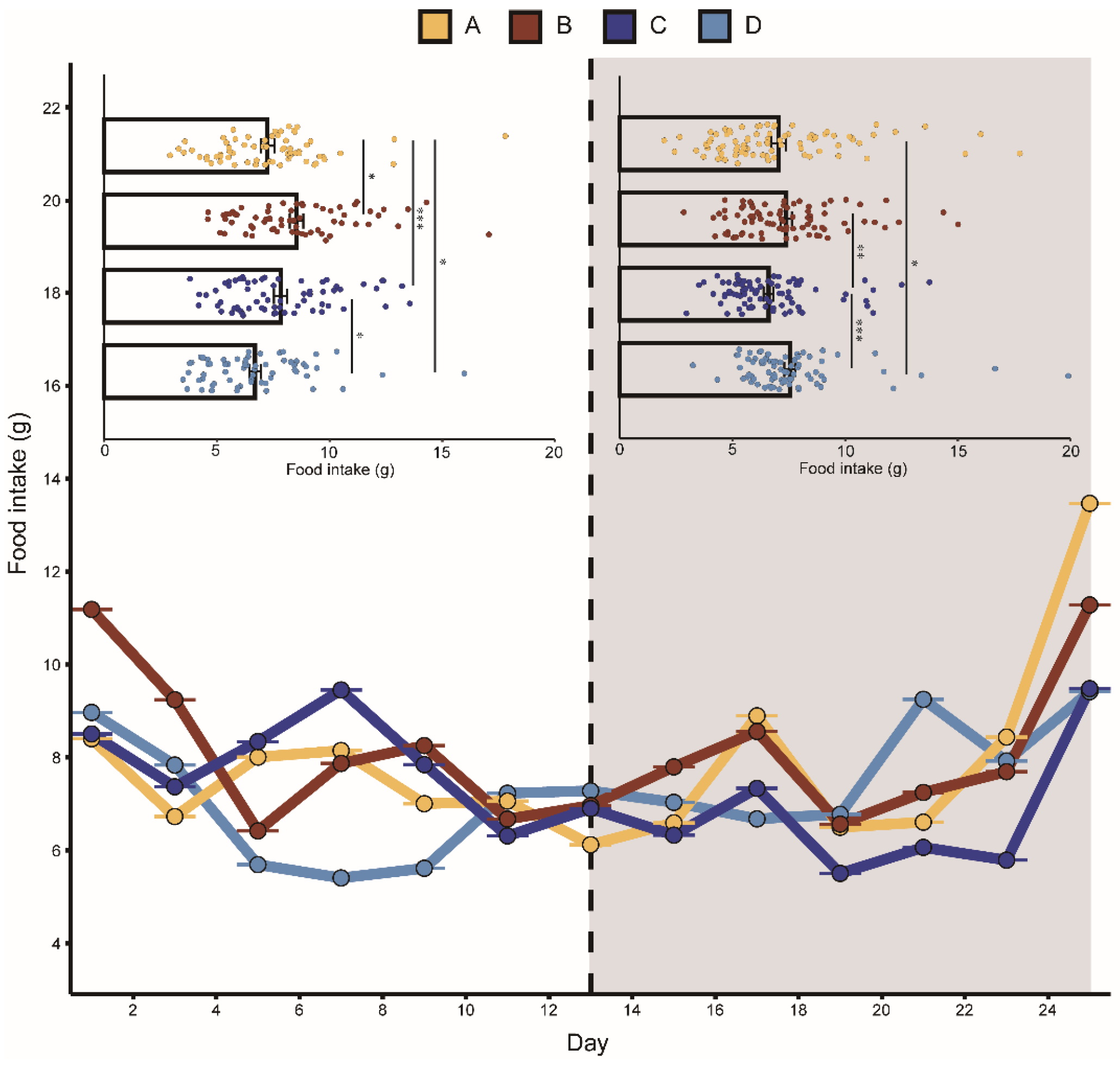
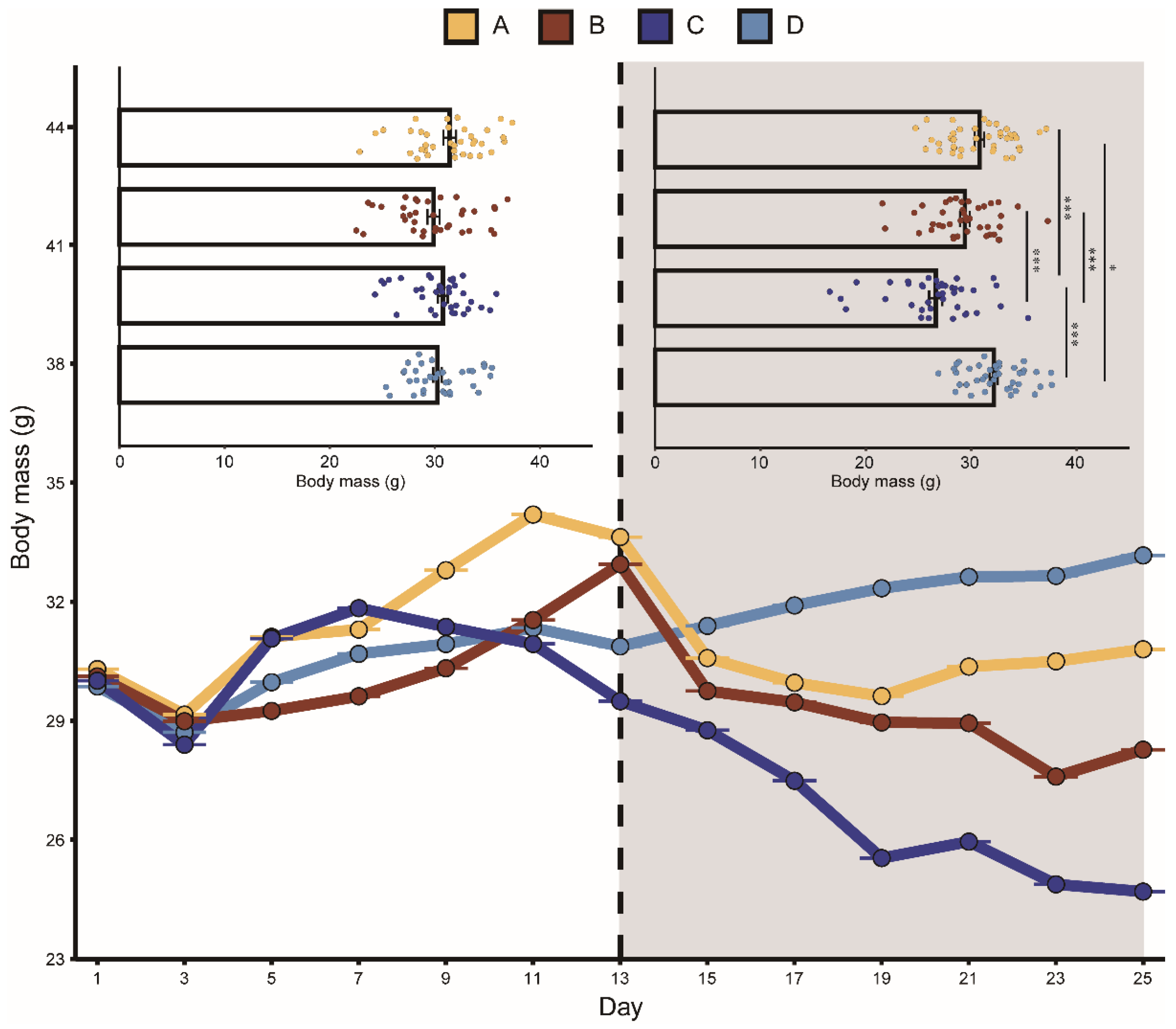
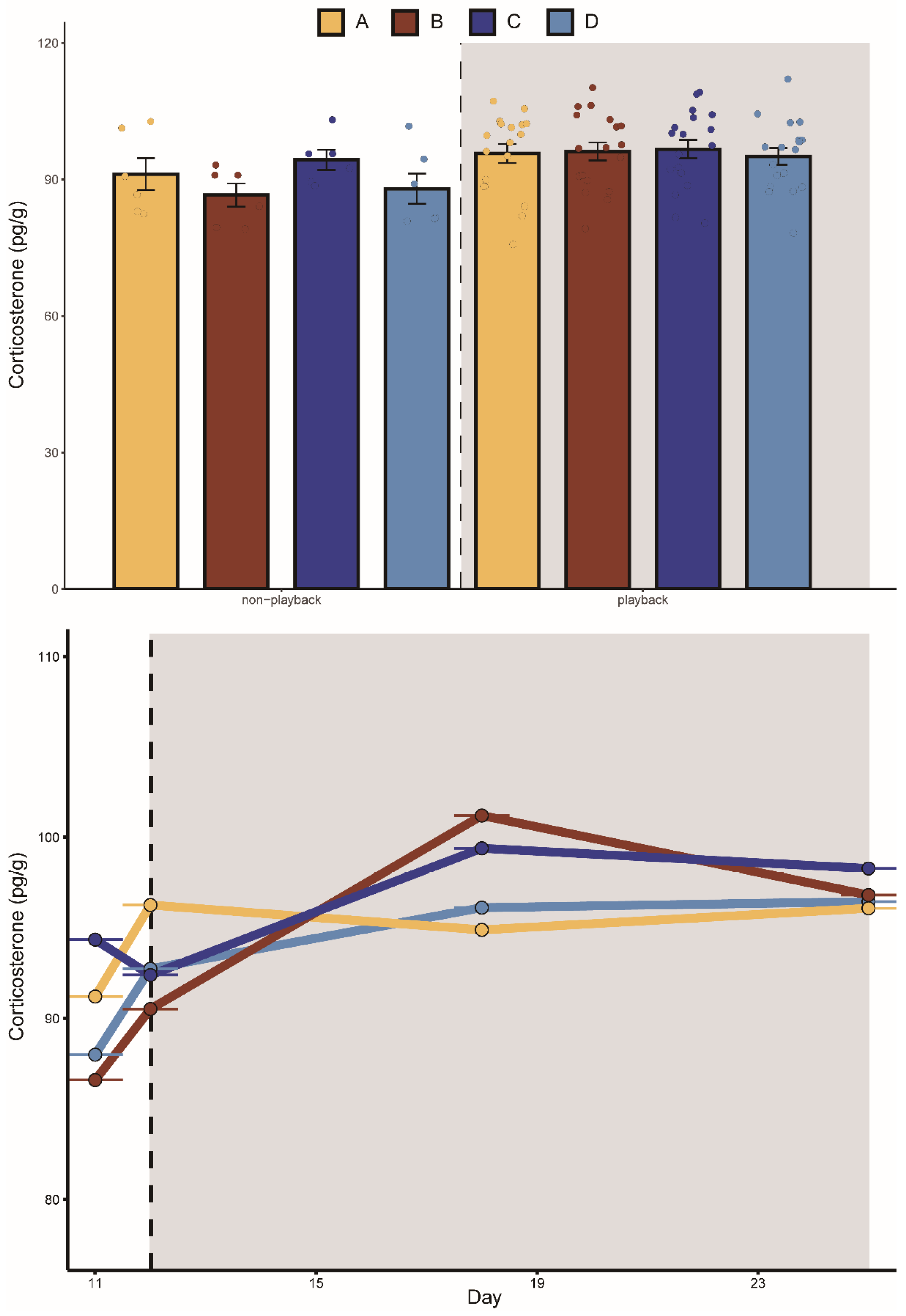
3.1. Effects of Aggressive Calls on Food Intake, Body Mass, and CORT Levels
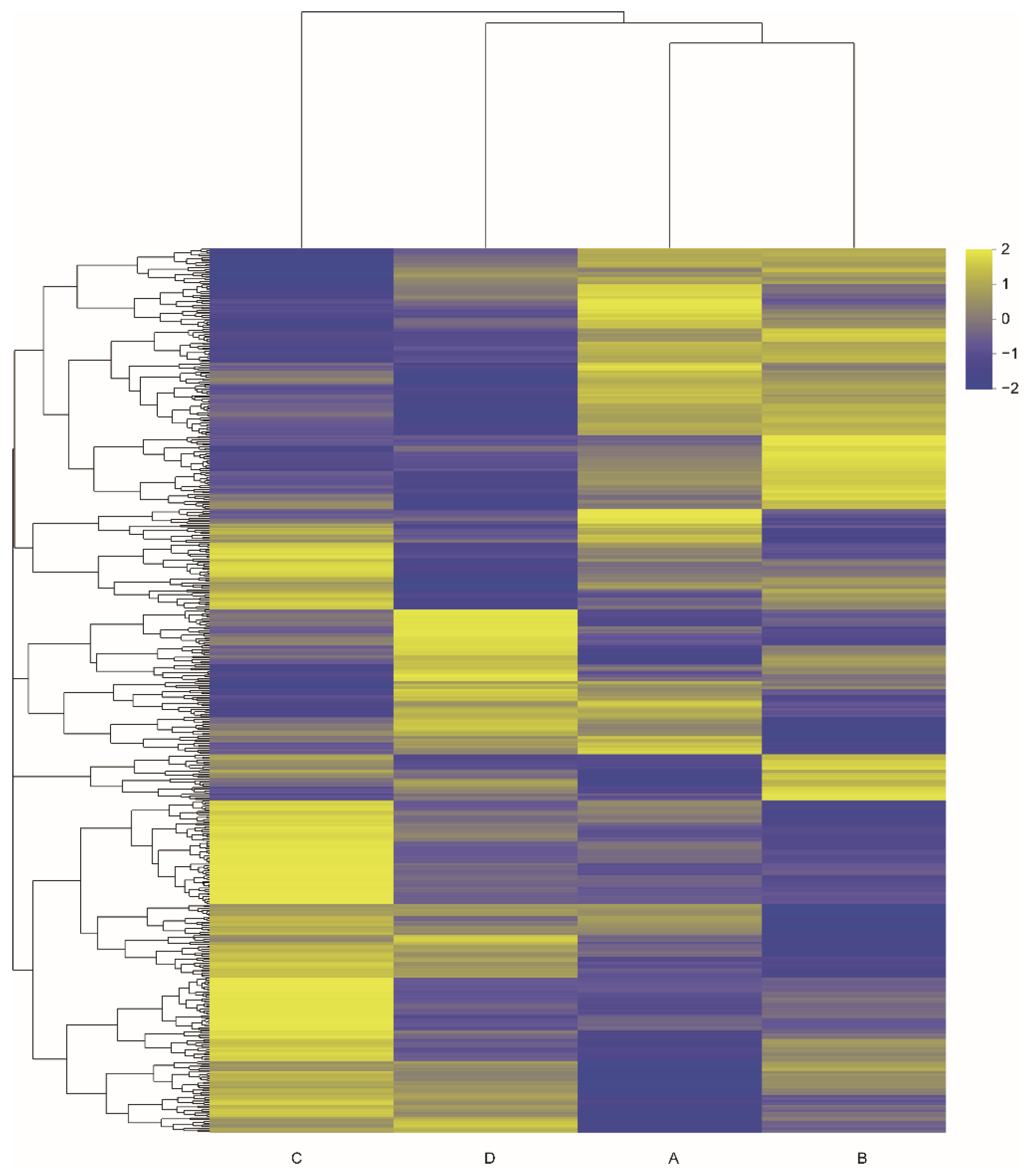
3.2. Effects of Aggressive Calls on Gene Expression
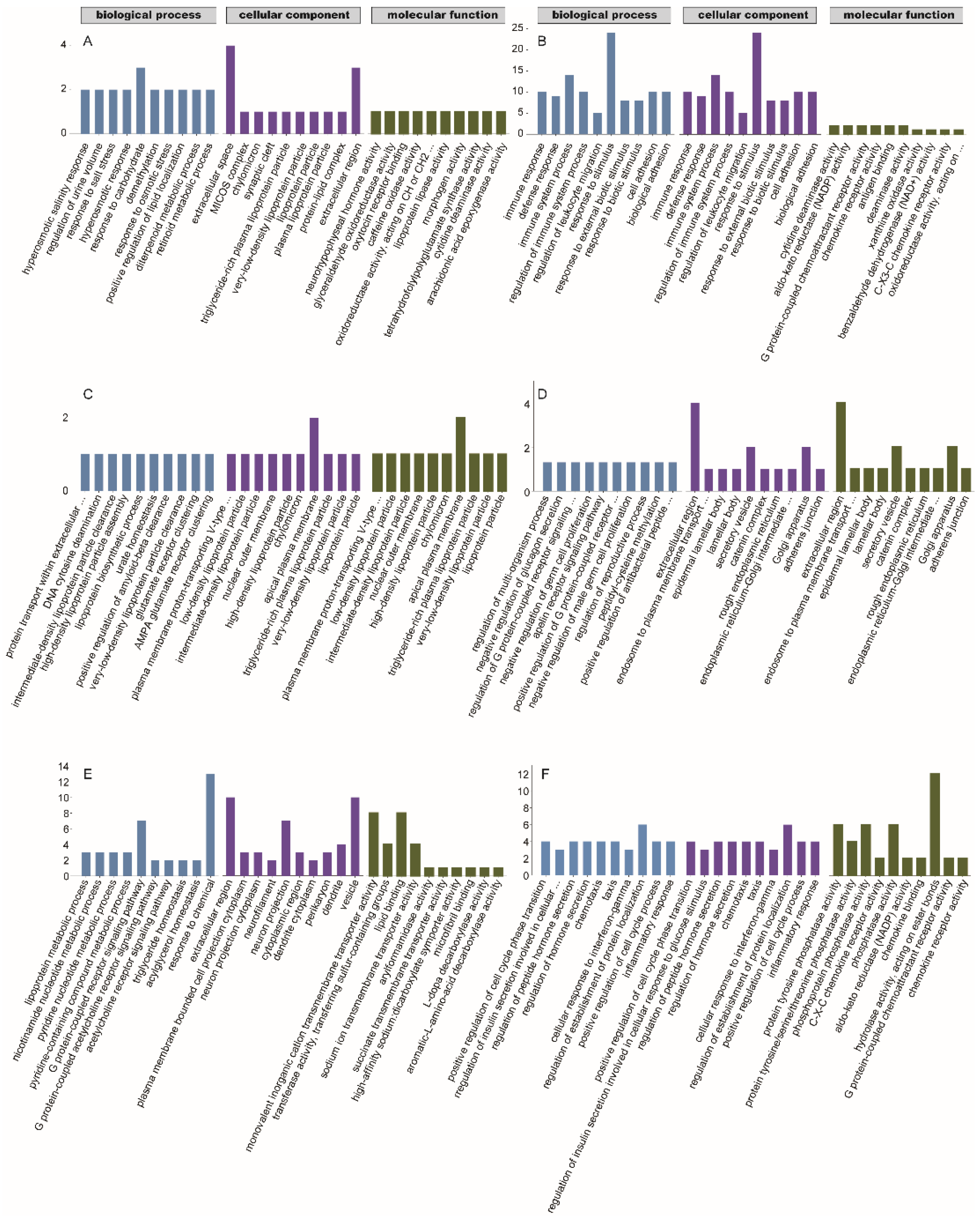
3.3. GO and KEGG Enrichment Analyses of Differential Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, E.O. Sociobiology: The new synthesis. Signs J. Women Cult. Soc. 1975, 46, 28–43. [Google Scholar]
- Lott, D.F. Intraspecific variation in the social systems of wild vertebrates. Behaviour 1984, 88, 266–325. [Google Scholar] [CrossRef]
- Blumstein, D.T. Group living. In Encyclopedia of Evolution; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Beauchamp, G. Social Predation: How Group Living Benefits Predators and Prey; Elsevier: London, UK, 2014. [Google Scholar]
- Bouwknecht, J.A.; Hijzen, T.H.; Vand, G.J. Startle responses, heart rate, and temperature in 5-ht1b receptor knockout mice. Neuroreport 2000, 11, 4097–4102. [Google Scholar] [CrossRef] [PubMed]
- Keeney, A.J.; Hogg, S.; Marsden, C.A. Alterations in core body temperature, locomotor activity, and corticosterone following acute and repeated social defeat of male nmri mice. Physiol. Behav. 2001, 74, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Dugatkin, L.A. Principles of Animal Behavior; W. W. Norton: New York, NY, USA, 2009. [Google Scholar]
- Smith, A.S.; Wang, Z. Salubrious effects of oxytocin on social stress-induced deficits. Horm. Behav. 2012, 61, 320–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, A.J.; Kramarcy, N.R. Neurochemical responses in stress: Relationships between the hypothalamic-pituitary-adrenal and catecholamine systems. In Drugs, Neurotransmitters, and Behavior; Springer: Berlin/Heidelberg, Germany, 1984. [Google Scholar]
- Axelrod, J.; Reisine, T.D. Stress hormones: Their interaction and regulation. Science 1984, 224, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Henry, J. Biological basis of the stress response. Integr. Physiol. Behav. Sci. 1992, 27, 66–83. [Google Scholar] [CrossRef]
- Dallman, M.F.; Akana, S.F.; Scribner, K.A. Stress, feedback and facilitation in the hypothalamo-pituitary-adrenal axis. J. Neuroendocrinol. 1992, 4, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Steffens, A.B.; Damsma, G.; Vand, G.J. Circulating free fatty acids, insulin, and glucose during chemical stimulation of hypothalamus in rats. Am. J. Physiol. 1984, 247, E765. [Google Scholar] [CrossRef]
- Harris, R.B.; Gu, H.; Mitchell, T.D. Increased glucocorticoid response to a novel stress in rats that have been restrained. Physiol. Behav. 2004, 81, 557–568. [Google Scholar] [CrossRef]
- Tamashiro, K.L.K.; Nguyen, M.M.N.; Ostrander, M.M.; Gardner, S.R.; Ma, L.Y.; Woods, S.C.; Sakai, R.R. Social stress and recovery: Implications for body weight and body composition. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2007, 293, R1864–R1874. [Google Scholar] [CrossRef]
- Tryon, M.S.; DeCant, R.; Laugero, K.D. Having your cake and eating it too: A habit of comfort food may link chronic social stress exposure and acute stress-induced cortisol hyporesponsiveness. Physiol. Behav. 2013, 114–115, 32–37. [Google Scholar] [CrossRef]
- Fuchs, E.; Kleinknecht, S. The influence of chronic social confrontation on oxygen consumption of tupaia belangeri under resting conditions. Ztschrift Für Sugetierkunde 1986, 51, 55–57. [Google Scholar]
- Kramer, M.; Hiemke, C.; Fuchs, E. Chronic psychosocial stress and antidepressant treatment in tree shrews: Time-dependent behavioral and endocrine effects. Neurosci. Biobehav. Rev. 1999, 23, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Calcagni, E.; Elenkov, I. Stress system activity, innate and t helper cytokines, and susceptibility to immune-related diseases. Ann. N. Y. Acad. Sci. 2006, 1069, 62–76. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Tsapkini, K.; Webster, K.T.; Ficek, B.N.; Desmond, J.E.; Onyike, C.U.; Rapp, B.; Frangakis, C.E.; Hillis, A.E. Electrical brain stimulation in different variants of primary progressive aphasia: A randomized clinical trial. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2018, 4, 461–472. [Google Scholar] [CrossRef]
- Johnstone, R.A. Eavesdropping and animal conflict. Proc. Natl. Acad. Sci. USA 2001, 98, 9177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Jiang, T.J.; Liu, H.; Wang, Y.Z.; Liu, Y.; Feng, J. Acoustic signalling of aggressive intent in the agonistic encounters of female asian particoloured bats. Anim. Behav. 2019, 149, 65–75. [Google Scholar] [CrossRef]
- Gadziola, M.A.; Grimsley, J.M.S.; Faure, P.A.; Wenstrup, J.J. Social vocalizations of big brown bats vary with behavioral context. PLoS ONE 2012, 7, e44550. [Google Scholar] [CrossRef] [Green Version]
- Poesel, A.; Dabelsteen, T. Three vocalization types in the blue tit cyanistes caeruleus: A test of the different signal-value hypothesis. Behaviour 2006, 143, 1529–1545. [Google Scholar]
- Morris-Drake, A.; Kern, J.M.; Radford, A.N. Experimental evidence for delayed post-conflict management behaviour in wild dwarf mongooses. eLife 2021, 10, e69196. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, P.L.; Souza, R.F.; de Oliveira, E.B.; Nali, R.C. Male aggressiveness in the tree frog dendropsophus branneri (anura, hylidae): Physical combat, fighting call, and escalated behavior. Caldasia 2021, 44, 203–207. [Google Scholar] [CrossRef]
- Chen, Y.M.; Zhang, X.P.; Zhang, Y.; Luo, W.J.; Chen, J. The effects of acute fear stress on emotional behaviors, plasmic hormone levels and the activation of cerebral erk1/2 of rats. Chin. J. Appl. Physiol. 2007, 03, 270–274. [Google Scholar]
- Greif, S.; Siemers, B.M. Innate recognition of water bodies in echolocating bats. Nat. Commun. 2010, 107, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, J.S.; Ohlemiller, S.M.K.; Suga, N. Analysis of acoustic elements and syntax in communication sounds emitted by mustached bats. J. Acoust. Soc. Am. 1994, 96, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Kroodsma, D.E. A re-evaluation of song development in the song sparrow. Anim. Behav. 1977, 25, 390–399. [Google Scholar] [CrossRef]
- Doupe, A.J.; Kuhl, P.K. Birdsong and human speech: Common themes and mechanisms. Annu. Rev. Neurosci. 1999, 22, 567–631. [Google Scholar] [CrossRef] [Green Version]
- Douglas, B.; Martin, M.; Ben, B.; Steve, W. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Li, B.; Dewey, C.N. Rsem: Accurate transcript quantification from rna-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323–340. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.R.; Ye, Z.; Stanton, R. Misuse of rpkm or tpm normalization when comparing across samples and sequencing protocols. RNA 2020, 74922, 120–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S. Kobas 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, C.; de Oliveira, C.M.; de Macedo, I.C.; Quevedo, A.S.; Filho, P.R.M.; da Silva, F.R.; Vercelino, R.; de Souza, I.C.C.; Caumo, I.; Torres, I.L.S. Hypercaloric diet modulates effects of chronic stress: A behavioral and biometric study on rats. Int. J. Biol. Stress 2015, 18(5), 514–523. [Google Scholar] [CrossRef]
- Eskesen, I.G.; Teilmann, J.; Geertsen, B.M.; Desportes, G.; Riget, F.; Dietz, R.; Larsen, F.; Siebert, U. Stress level in wild harbour porpoises (phocoena phocoena) during satellite tagging measured by respiration, heart rate and cortisol. J. Mar. Biol. Assoc. United Kingd. 2009, 89, 885–892. [Google Scholar] [CrossRef]
- Lee, S.B.; Kim, H.G.; Lee, J.S.; Kim, W.Y.; Lee, M.M.; Kim, Y.H.; Lee, J.J.; Kim, H.S.; Son, C.G. Intermittent restraint-induced sympathetic activation attenuates hepatic steatosis and inflammation in a high fat-diet fed-mouse model. AJP Gastrointest. Liver Physiol. 2019, 317, G811–G823. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Ryan, K.K. Neuroendocrine circuits governing energy balance and stress regulation: Functional overlap and therapeutic implications. Cell Metab. 2014, 19, 910–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallman, M.F.; Pecoraro, N.; Akana, S.F.; la Fleur, S.E.; Gomez, F.; Houshyar, H.; Bell, M.E.; Bhatnagar, S.; Laugero, K.K.; Manalo, S. Chronic stress and obesity: A new view of comfort food. Proc. Natl. Acad. Sci. USA 2018, 100, 11696–11701. [Google Scholar] [CrossRef] [Green Version]
- Pastel, E.; Pointud, J.J.; Volat, F.; Martinez, A.; Martinez, A.M. Aldo-keto reductases 1b in endocrinology and metabolism. Front. Pharmacol. 2012, 3, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Albalat, A.; Sánchez-Gurmaches, J.; Gutiérrez, J.; Navarro, I. Regulation of lipoprotein lipase activity in rainbow trout (oncorhynchus mykiss) tissues. Gen. Comp. Endocrinol. 2006, 146, 226–235. [Google Scholar] [CrossRef]
- Nyrén, R.; Makoveichuk, E.; Malla, S.; Kersten, S.; Nilsson, S.K.; Ericsson, M.; Olivecrona, G. Lipoprotein lipase in mouse kidney: Effects of nutritional status and high-fat diet. Am. J. Physiol. Ren. Physiol. 2019, 316, F558–F571. [Google Scholar] [CrossRef]
- Kastrenopoulou, A.; Kypreos, K.; Papachristou, N.I.; Georgopoulos, N.; Mastora, I.; Olivgeri, I.P.; Spentzopoulou, A.; Nikitovic, D.; Kontogeorgakos, V.; Blair, E.C.; et al. Apoa1 deficiency reshapes the phenotypic and molecular characteristics of bone marrow adipocytes in mice. Int. J. Mol. Sci. 2022, 23, 4834. [Google Scholar] [CrossRef] [PubMed]
- Farahani, P.; Fisler, J.S.; Wong, H.; Diament, A.L.; Yi, N.; Warden, C.H. Reciprocal hemizygosity analysis of mouse hepatic lipase reveals influence on obesity. Obes. Res. 2004, 12, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Vlahakis, S.R.; Villasis-Keever, A.; Gomez, T.; Vanegas, M.; Vlahakis, N.; Paya, C.V. G protein-coupled chemokine receptors induce both survival and apoptotic signaling pathways. J. Immunol. 2002, 169, 5546–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Wang, Y.X.; Yu, J.; Yi, S. Critical signaling pathways during wallerian degeneration of peripheral nerve. Neural Regen. Res. 2017, 12, 995–1002. [Google Scholar]
- Kim, S.; Lim, B.M.; Oh, E.Y.; Jeong, C.G.; Kim, W.; Lee, K.; Lee, S.M.; Kim, J.-M. Comprehensive transcriptomic comparison between porcine cd8− and cd8+ gamma delta t cells revealed distinct immune phenotype. Animals 2021, 11, 2165. [Google Scholar] [CrossRef]
- Turrina, N.P.; Plata-Salamán, C.R. Cytokine–cytokine interactions and the brain. Brain Res. Bull. 2000, 51, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xie, S.B.; Wu, C.H.; Hu, Y.; Zhang, Q.; Li, S.; Fan, Y.G.; Leng, R.X.; Pan, H.F.; Xiong, H.B.; et al. Coagulation cascade and complement system in systemic lupus erythematosus. Oncotarget 2018, 9, 14862–14881. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, K.; Yamaguchi, M.; Suguro, M.; Choi, W.; Ji, Y.; Xiao, J.Y.; Zhang, W.; Ogawa, S.; Katayama, B.; Shiku, H.; et al. Gene expression profiling of diffuse large b-cell lymphoma supervised by cd21 expression. BJHaem 2008, 142, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Wiel, M.D.; Porteiro, B.; Belt, S.C.; Vogels, E.W.M.; Bolt, I.; Vermeulen, J.L.M.; de Waart, D.R.; Verheij, J.; Muncan, V.; Oude Elferink, R.P.J.; et al. Differential and organ-specific functions of organic solute transporter alpha and beta in experimental cholestasis. JHEP Rep. Innov. Hepatol. 2022, 4, 100463. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Intake | Body Mass | Corticosterone | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Estimate (β ± SE) | t | p | Estimate (β ± SE) | t | p | Estimate (β ± SE) | t | p | |
| Random effects | |||||||||
| Bat individual identity | 0.91 | 2.38 | 0.00 | ||||||
| Residual | 2.21 | 2.31 | 8.10 | ||||||
| Fixed effects | |||||||||
| Intercept | 6.45 ± 0.48 | 13.32 | <0.001 | 30.33 ± 1.06 | 28.62 | <0.001 | 84.80 ± 3.85 | 22.00 | <0.001 |
| Day | 0.04 ± 0.02 | 1.65 | 0.10 | −0.01 ± 0.03 | −0.42 | 0.67 | 0.29 ± 0.18 | 1.61 | 0.11 |
| Group A | 0.55 ± 0.65 | 0.85 | 0.40 | 1.22 ± 1.47 | 0.83 | 0.41 | 3.21 ± 4.68 | 0.69 | 0.49 |
| Group B | 1.84 ± 0.65 | 2.81 | 0.08 | −0.28 ± 1.47 | −0.19 | 0.85 | −1.4 ± 4.68 | −0.30 | 0.76 |
| Group C | 1.13 ± 0.65 | 1.74 | 0.09 | 0.35 ± 1.47 | 0.24 | 0.81 | 6.35 ± 4.68 | 1.36 | 0.18 |
| Playback | 0.35 ± 0.47 | 0.73 | 0.46 | 2.07 ± 0.66 | 3.11 | 0.002 | 4.98 ± 4.04 | 1.23 | 0.22 |
| Group A: Playback | −1.05 ± 0.51 | −2.04 | 0.04 | −2.58 ± 0.71 | −3.64 | <0.001 | −2.57 ± 5.40 | −0.47 | 0.63 |
| Group B: Playback | −1.20 ± 0.51 | −3.88 | <0.001 | −2.44 ± 0.71 | −3.43 | <0.001 | 2.48 ± 5.40 | 0.46 | 0.65 |
| Group C: Playback | −2.08 ± 0.51 | −4.04 | <0.001 | −5.80 ± 0.71 | −8.16 | <0.001 | −4.8 ± 5.40 | −0.88 | 0.38 |
| Comparative Groups | Up | Down | Total |
|---|---|---|---|
| A vs. D | 28 | 43 | 71 |
| B vs. D | 69 | 38 | 107 |
| C vs. D | 51 | 19 | 70 |
| A vs. B | 18 | 27 | 45 |
| A vs. C | 58 | 134 | 192 |
| B vs. C | 55 | 104 | 159 |
| Pathway Described | ID | p-Value | Number of DEGs |
|---|---|---|---|
| A vs. D | |||
| Folate biosynthesis | ko00790 | 0.001094 | 2 |
| Glycerolipid metabolism | ko00561 | 0.006939 | 2 |
| C vs. D | |||
| Complement and coagulation cascades | ko04610 | 0.006103 | 2 |
| Bile secretion | ko04976 | 0.007234 | 2 |
| A vs. B | |||
| Arachidonic acid metabolism | ko00590 | 0.001225 | 2 |
| Ferroptosis | ko04216 | 0.001382 | 2 |
| Mineral absorption | ko04978 | 0.002842 | 2 |
| B vs. C | |||
| Bile secretion | ko04976 | 0.000972 | 4 |
| Progesterone-mediated oocyte maturation | ko04914 | 0.001312 | 4 |
| Oocyte meiosis | ko04114 | 0.002818 | 4 |
| Mineral absorption | ko04978 | 0.007589 | 3 |
| Platinum drug resistance | ko01524 | 0.009405 | 3 |
| Phenylalanine metabolism | ko00360 | 0.009523 | 2 |
| A vs. C | |||
| Cocaine addiction | ko05030 | 0.000228 | 4 |
| Mineral absorption | ko04978 | 0.000632 | 4 |
| Steroid hormone biosynthesis | ko00140 | 0.001166 | 3 |
| Cholesterol metabolism | ko04979 | 0.001601 | 3 |
| Alcoholism | ko05034 | 0.005462 | 4 |
| Amphetamine addiction | ko05031 | 0.008076 | 3 |
| Phenylalanine metabolism | ko00360 | 0.009040 | 2 |
| Fat digestion and absorption | ko04975 | 0.009533 | 2 |
| B vs. D | |||
| Viral protein interaction with cytokine and cytokine receptor | ko04061 | 0.000005 | 5 |
| Chemokine signaling pathway | ko04062 | 0.000051 | 6 |
| Leishmaniasis | ko05140 | 0.000065 | 5 |
| Cytokine–cytokine receptor interaction | ko04060 | 0.000085 | 6 |
| Cell adhesion molecules | ko04514 | 0.000421 | 5 |
| Pathogenic Escherichia coli infection | ko05130 | 0.000515 | 6 |
| Th1 and Th2 cell differentiation | ko04658 | 0.000599 | 4 |
| NOD-like receptor signaling pathway | ko04621 | 0.000619 | 5 |
| Hematopoietic cell lineage | ko04640 | 0.000671 | 4 |
| Leukocyte transendothelial migration | ko04670 | 0.000876 | 4 |
| Chagas disease | ko05142 | 0.000876 | 4 |
| Rheumatoid arthritis | ko05323 | 0.001321 | 4 |
| Influenza A | ko05164 | 0.003221 | 4 |
| Staphylococcus aureus infection | ko05150 | 0.006513 | 3 |
| Th17 cell differentiation | ko04659 | 0.007996 | 3 |
| Caffeine metabolism | ko00232 | 0.008818 | 1 |
| Phagosome | ko04145 | 0.009057 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhou, R.; Feng, L.; Wang, H.; Feng, J.; Wu, H. Effects of Different-Syllable Aggressive Calls on Food Intake and Gene Expression in Vespertilio sinensis. Animals 2023, 13, 306. https://doi.org/10.3390/ani13020306
Li X, Zhou R, Feng L, Wang H, Feng J, Wu H. Effects of Different-Syllable Aggressive Calls on Food Intake and Gene Expression in Vespertilio sinensis. Animals. 2023; 13(2):306. https://doi.org/10.3390/ani13020306
Chicago/Turabian StyleLi, Xin, Ruizhu Zhou, Lei Feng, Hui Wang, Jiang Feng, and Hui Wu. 2023. "Effects of Different-Syllable Aggressive Calls on Food Intake and Gene Expression in Vespertilio sinensis" Animals 13, no. 2: 306. https://doi.org/10.3390/ani13020306
APA StyleLi, X., Zhou, R., Feng, L., Wang, H., Feng, J., & Wu, H. (2023). Effects of Different-Syllable Aggressive Calls on Food Intake and Gene Expression in Vespertilio sinensis. Animals, 13(2), 306. https://doi.org/10.3390/ani13020306