
Heat Shock Protein Response to Stress in Poultry: A Review

Abstract
:Simple Summary
Abstract
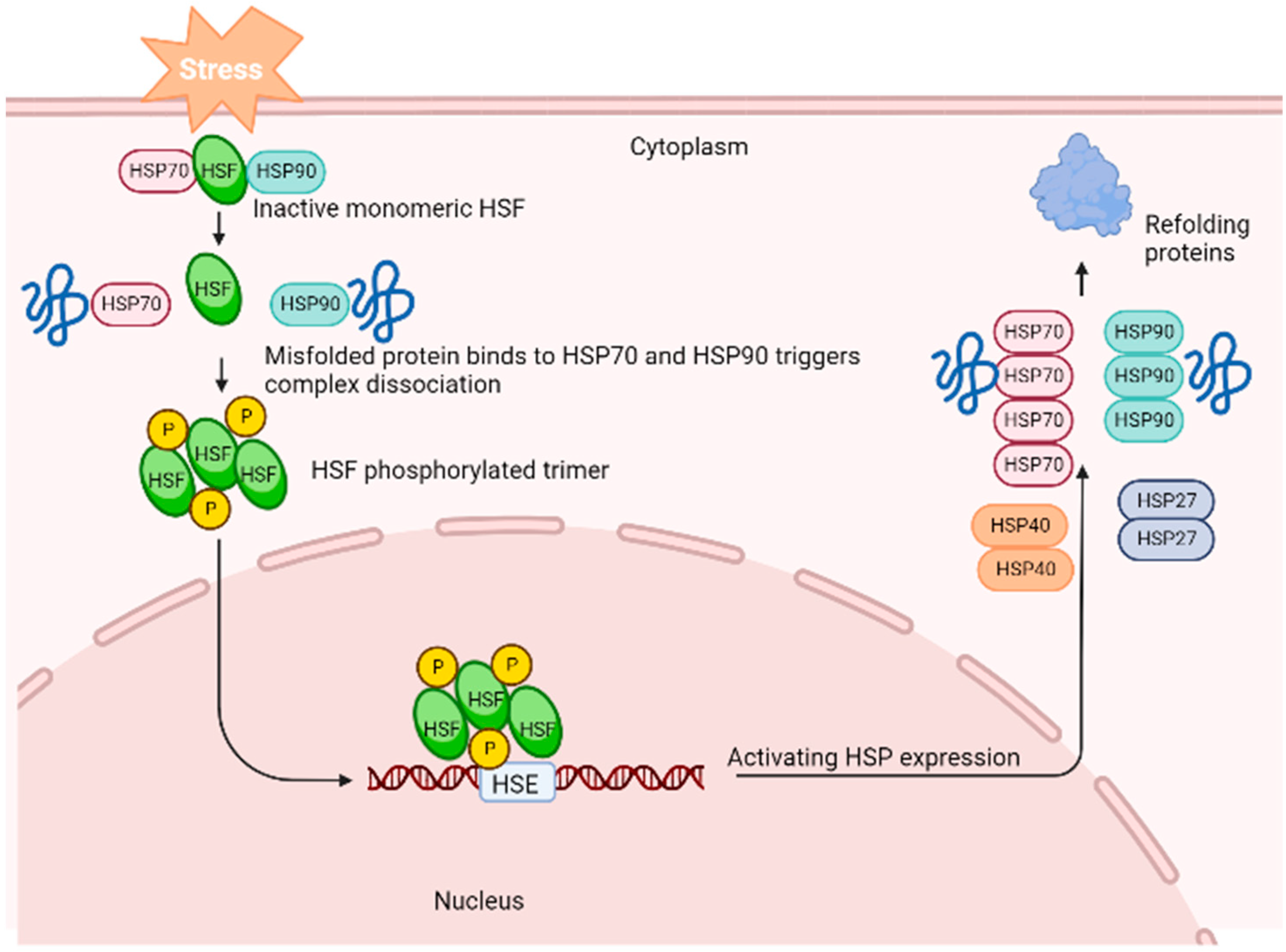
1. Introduction
2. Heat Stress (HS)
3. Cold Stress
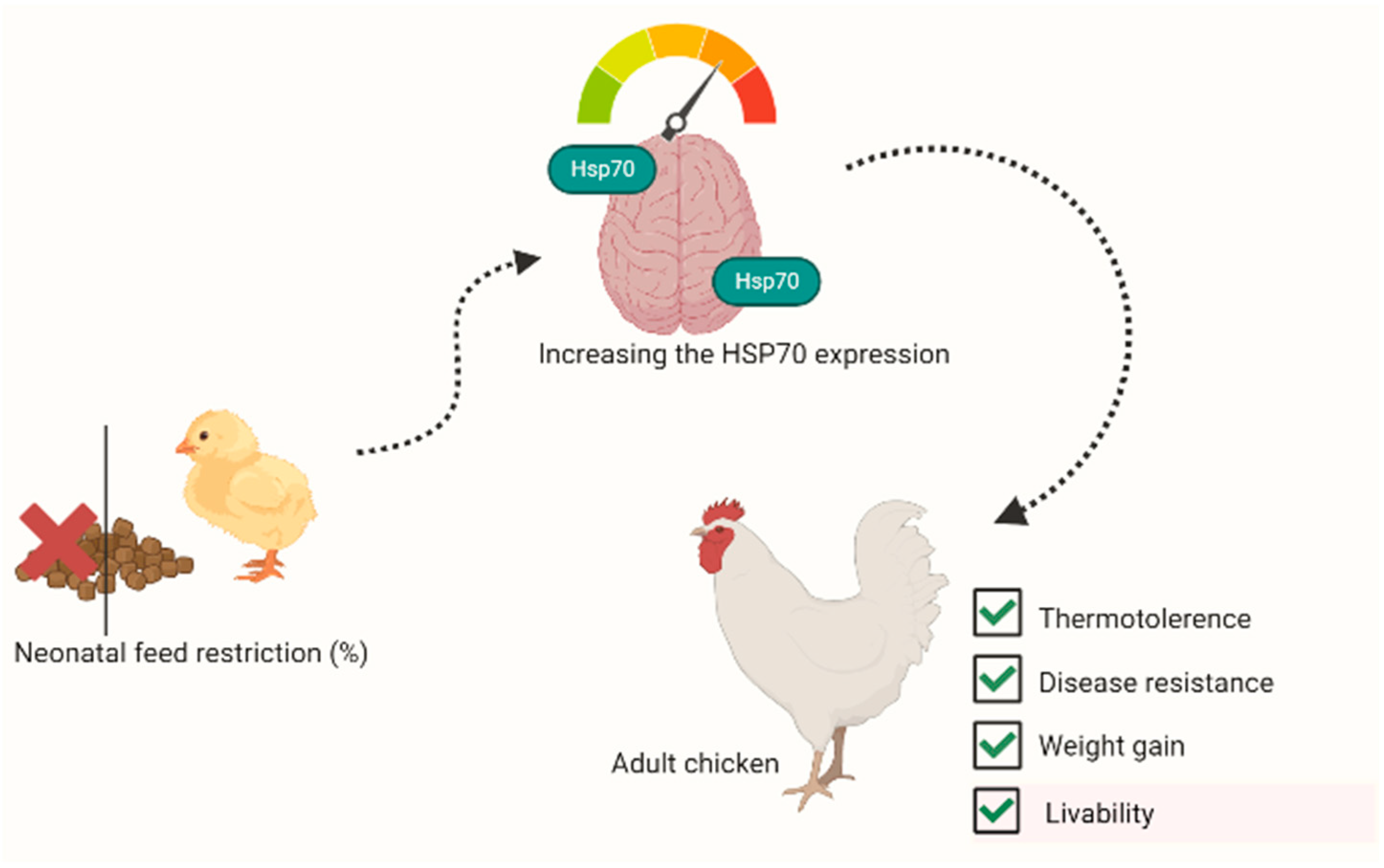
4. Feed Restriction (FR)
5. Pre-Slaughter Operations
6. Social Stress
7. Stocking Density
8. Human Contact
9. Heavy Metals
10. Mycotoxin
11. Factors Modulating HSP Expression
11.1. Nutrition
11.2. Phytochemicals
11.3. Genetics
11.4. Age
11.5. Gender
11.6. Fear
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Gateway to Poultry Production and Products. Available online: http://www.fao.org/poultry-production-products/production/en/ (accessed on 27 September 2022).
- Bessei, W. Impact of animal welfare on worldwide poultry production. World Poult. S. J. 2018, 74, 211–224. [Google Scholar] [CrossRef]
- Broom, D. The use of the concept animal welfare in European conventions, regulations and directives. Food Chain 2001, 2001, 148–151. [Google Scholar]
- Koolhaas, J.M.; Bartolomucci, A.; Buwalda, B.; de Boer, S.F.; Flugge, G.; Korte, S.M.; Meerlo, P.; Murison, R.; Olivier, B.; Palanza, P.; et al. Stress revisited: A critical evaluation of the stress concept. Neurosci. Biobehav. Rev. 2011, 35, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Lai, Y.M.; Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veissier, I.; Boissy, A. Stress and welfare: Two complementary concepts that are intrinsically related to the animal’s point of view. Physiol. Behav. 2007, 92, 429–433. [Google Scholar] [CrossRef]
- Kregel, K.C. Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 92, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef] [Green Version]
- Welch, W.J. How cells respond to stress. Sci. Am. 1993, 268, 56–64. [Google Scholar] [CrossRef]
- Sonna, L.A.; Fujita, J.; Gaffin, S.L.; Lilly, C.M. Invited review: Effects of heat and cold stress on mammalian gene expression. J. Appl. Physiol. 2002, 92, 1725–1742. [Google Scholar] [CrossRef] [Green Version]
- Etches, R.; John, T.; Gibbins, A.V. Behavioural, physiological, neuroendocrine and molecular responses to heat stress. Poult. Prod. Hot Clim. 2008, 2, 48–79. [Google Scholar]
- Ritossa, F. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia 1962, 18, 571–573. [Google Scholar] [CrossRef]
- Tissieres, A.; Mitchell, H.K.; Tracy, U.M. Protein synthesis in salivary glands of Drosophila melanogaster: Relation to chromosome puffs. J. Mol. Biol. 1974, 84, 389–398. [Google Scholar] [CrossRef]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Shende, P.; Bhandarkar, S.; Prabhakar, B. Heat shock proteins and their protective roles in stem cell biology. Stem Cell Rev. 2019, 15, 637–651. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Hunt, C.; Huang, S.Y.; Berg, K.L.; Banerji, S.S. Organization, nucleotide sequence, and transcription of the chicken HSP70 gene. J. Biol. Chem. 1986, 261, 12692–12699. [Google Scholar] [CrossRef]
- Tamzil, M.; Noor, R.; Hardjosworo, P.; Manalu, W.; Sumantri, C. Acute heat stress responses of three lines of chickens with different heat shock protein (HSP)-70 genotypes. Int. J. Poult. Sci. 2013, 12, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Garrido, C.; Solary, E. A role of HSPs in apoptosis through “protein triage”? Cell Death Differ. 2003, 10, 619. [Google Scholar] [CrossRef]
- Madkour, M.; Salman, F.M.; El-Wardany, I.; Abdel-Fattah, S.A.; Alagawany, M.; Hashem, N.M.; Abdelnour, S.A.; El-Kholy, M.S.; Dhama, K. Mitigating the detrimental effects of heat stress in poultry through thermal conditioning and nutritional manipulation. J. Therm. Biol. 2022, 103, 103169. [Google Scholar] [CrossRef]
- Hartl, F.U.; Hayer-Hartl, M. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Tixier-Boichard, M.; Leenstra, F.; Flock, D.; Hocking, P.; Weigend, S. A century of poultry genetics. World Poult. Sci. J. 2012, 68, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Grandin, T.; Deesing, M.J. Genetics and animal welfare. In Genetics and The Behavior of Domestic Animals; Elsevier: Amsterdam, The Netherlands, 2022; pp. 507–548. [Google Scholar]
- Wang, S.; Edens, F. Hsp70 mRNA expression in heat-stressed chickens. Comp. Biochem. Physiol Part. B Comp. Biochem. 1994, 107, 33–37. [Google Scholar] [CrossRef]
- Einat, M.F.; Haberfeld, A.; Shamay, A.; Horev, G.; Hurwitz, S.; Yahav, S. A novel 29-kDa chicken heat shock protein. Poult. Sci. 1996, 75, 1528–1530. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Bao, E.; Yan, J.; Lei, L. Expression and localization of Hsps in the heart and blood vessel of heat-stressed broilers. Cell Stress Chaperones 2008, 13, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Tang, L.; Lu, L.; Zhang, L.; Xi, L.; Liu, H.C.; Odle, J.; Luo, X. Differential expression of heat shock transcription factors and heat shock proteins after acute and chronic heat stress in laying chickens (Gallus gallus). PLoS ONE 2014, 9, e102204. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Yu, J.; Bao, E. Expression of heat shock protein 90 (Hsp90) and transcription of its corresponding mRNA in broilers exposed to high temperature. Br. Poult. Sci. 2009, 50, 504–511. [Google Scholar] [CrossRef]
- Wang, S.H.; Cheng, C.Y.; Tang, P.C.; Chen, C.F.; Chen, H.H.; Lee, Y.P.; Huang, S.Y. Differential gene expressions in testes of L2 strain Taiwan country chicken in response to acute heat stress. Theriogenology 2013, 79, 374–382. [Google Scholar] [CrossRef]
- Wang, S.H.; Cheng, C.Y.; Tang, P.C.; Chen, C.F.; Chen, H.H.; Lee, Y.P.; Huang, S.Y. Acute heat stress induces differential gene expressions in the testes of a broiler-type strain of Taiwan country chickens. PLoS ONE 2015, 10, e0125816. [Google Scholar] [CrossRef]
- Greene, E.S.; Emami, N.K.; Dridi, S. Research Note: Phytobiotics modulate the expression profile of circulating inflammasome and cyto(chemo)kine in whole blood of broilers exposed to cyclic heat stress. Poult. Sci. 2021, 100, 100801. [Google Scholar] [CrossRef]
- Kang, D.; Shim, K. Early heat exposure effect on the heat shock proteins in broilers under acute heat stress. Poult. Sci. 2021, 100, 100964. [Google Scholar] [CrossRef]
- Vinoth, A.; Thirunalasundari, T.; Tharian, J.A.; Shanmugam, M.; Rajkumar, U. Effect of thermal manipulation during embryogenesis on liver heat shock protein expression in chronic heat stressed colored broiler chickens. J. Therm. Biol. 2015, 53, 162–171. [Google Scholar] [CrossRef]
- Siddiqui, S.H.; Kang, D.; Park, J.; Khan, M.; Shim, K. Chronic heat stress regulates the relation between heat shock protein and immunity in broiler small intestine. Sci. Rep. 2020, 10, 18872. [Google Scholar] [CrossRef]
- Toplu, H.D.O.; Tunca, R.; Aypak, S.U.; Coven, F.; Epikmen, E.T.; Karaarslan, S.; Yagin, O. Effects of heat conditioning and dietary ascorbic acid supplementation on heat shock protein 70 expression, blood parameters and fear-related behavior in broilers subjected to heat stress. Acta Sci. Vet. 2014, 42, 1–8. [Google Scholar]
- Al-Zghoul, M.B.; Ismail, Z.B.; Dalab, A.E.; Al-Ramadan, A.; Althnaian, T.A.; Al-Ramadan, S.Y.; Ali, A.M.; Albokhadaim, I.F.; Al Busadah, K.A.; Eljarah, A.; et al. Hsp90, Hsp60 and HSF-1 genes expression in muscle, heart and brain of thermally manipulated broiler chicken. Res. Vet. Sci. 2015, 99, 105–111. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. Physiological responses of 3 chicken breeds to acute heat stress. Poult. Sci. 2011, 90, 1435–1440. [Google Scholar] [CrossRef]
- Zhao, F.Q.; Zhang, Z.W.; Wang, C.; Zhang, B.; Yao, H.D.; Li, S.; Xu, S.W. The role of heat shock proteins in inflammatory injury induced by cold stress in chicken hearts. Cell Stress Chaperones 2013, 18, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Q.; Zhang, Z.W.; Yao, H.D.; Wang, L.L.; Liu, T.; Yu, X.Y.; Li, S.; Xu, S.W. Effects of cold stress on mRNA expression of immunoglobulin and cytokine in the small intestine of broilers. Res. Vet. Sci. 2013, 95, 146–155. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, R.; Su, Y.; Bi, Y.; Li, X.; Zhang, X.; Li, J.; Bao, J. Effects of Acute Cold Stress After Long-Term Cold Stimulation on Antioxidant Status, Heat Shock Proteins, Inflammation and Immune Cytokines in Broiler Heart. Front. Physiol. 2018, 9, 1589. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Li, S.; Xin, H.; Li, J.; Li, X.; Zhang, R.; Li, J.; Bao, J. Proper cold stimulation starting at an earlier age can enhance immunity and improve adaptability to cold stress in broilers. Poult. Sci. 2020, 99, 129–141. [Google Scholar] [CrossRef]
- Tarkhan, A.H.; Saleh, K.M.M.; Al-Zghoul, M.B. HSF3 and Hsp70 Expression during Post-Hatch Cold Stress in Broiler Chickens Subjected to Embryonic Thermal Manipulation. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Leandro, N.S.; Gonzales, E.; Ferro, J.A.; Ferro, M.I.; Givisiez, P.E.; Macari, M. Expression of heat shock protein in broiler embryo tissues after acute cold or heat stress. Mol. Reprod. Dev. 2004, 67, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Che Norma, M.T.; Israf, D.A.; Omar, A.R. The effect of early-age food restriction on heat shock protein 70 response in heat-stressed female broiler chickens. Br. Poult. Sci. 2002, 43, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Soleimani, A.; Khalil, M.; Omar, A.; Raha, A. Inhibition of adrenal steroidogenesis and heat shock protein 70 induction in neonatally feed restricted broiler chickens under heat stress condition. Arch. Für Geflügelkunde 2011, 75, 246–252. [Google Scholar]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. Neonatal feed restriction modulates circulating levels of corticosterone and expression of glucocorticoid receptor and heat shock protein 70 in aged Japanese quail exposed to acute heat stress. Poult. Sci. 2011, 90, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Najafi, P.; Zulkifli, I.; Soleimani, A.F.; Kashiani, P. The effect of different degrees of feed restriction on heat shock protein 70, acute phase proteins, and other blood parameters in female broiler breeders. Poult. Sci. 2015, 94, 2322–2329. [Google Scholar] [CrossRef]
- Najafi, P.; Zulkifli, I.; Soleimani, A.F.; Goh, Y.M. Acute phase proteins response to feed deprivation in broiler chickens. Poult. Sci. 2016, 95, 760–763. [Google Scholar] [CrossRef]
- Najafi, P.; Zulkifli, I.; Soleimani, A.F. Inhibition of corticosterone synthesis and its effect on acute phase proteins, heat shock protein 70, and interleukin-6 in broiler chickens subjected to feed restriction. Poult. Sci. 2018, 97, 1441–1447. [Google Scholar] [CrossRef]
- Zulkifli, I.; Al-Aqil, A.; Omar, A.R.; Sazili, A.Q.; Rajion, M.A. Crating and heat stress influence blood parameters and heat shock protein 70 expression in broiler chickens showing short or long tonic immobility reactions. Poult. Sci. 2009, 88, 471–476. [Google Scholar] [CrossRef]
- Sun, F.; Zuo, Y.Z.; Ge, J.; Xia, J.; Li, X.N.; Lin, J.; Zhang, C.; Xu, H.L.; Li, J.L. Transport stress induces heart damage in newly hatched chicks via blocking the cytoprotective heat shock response and augmenting nitric oxide production. Poult. Sci. 2018, 97, 2638–2646. [Google Scholar] [CrossRef]
- Xing, T.; Wang, M.F.; Han, M.Y.; Zhu, X.S.; Xu, X.L.; Zhou, G.H. Expression of heat shock protein 70 in transport-stressed broiler pectoralis major muscle and its relationship with meat quality. Animal 2017, 11, 1599–1607. [Google Scholar] [CrossRef]
- Delezie, E.; Swennen, Q.; Buyse, J.; Decuypere, E. The effect of feed withdrawal and crating density in transit on metabolism and meat quality of broilers at slaughter weight. Poult. Sci. 2007, 86, 1414–1423. [Google Scholar] [CrossRef]
- Hoekstra, K.A.; Iwama, G.K.; Nichols, C.R.; Godin, D.V.; Cheng, K.M. Increased heat shock protein expression after stress in Japanese quail. Stress 1998, 2, 265–272. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. The relationship between adrenocortical function and Hsp70 expression in socially isolated Japanese quail. Comp. Biochem. Physiol. A Mol. Integr Physiol. 2012, 161, 140–144. [Google Scholar] [CrossRef]
- Najafi, P.; Zulkifli, I.; Jajuli, N.A.; Farjam, A.S.; Ramiah, S.K.; Amir, A.A.; O’Reily, E.; Eckersall, D. Environmental temperature and stocking density effects on acute phase proteins, heat shock protein 70, circulating corticosterone and performance in broiler chickens. Int. J. Biometeorol. 2015, 59, 1577–1583. [Google Scholar] [CrossRef]
- Beloor, J.; Kang, H.; Kim, Y.; Subramani, V.; Jang, I.; Sohn, S.; Moon, Y.S. The effect of stocking density on stress related genes and telomeric length in broiler chickens. Asian-Australas. J. Anim. Sci. 2010, 23, 437–443. [Google Scholar] [CrossRef]
- Shewita, R.S.; El-Naggar, K.; Abd El Naby, W.S. Influence of dietary vitamin c on growth performance, blood biochemical parameters and transcript levels of heat shock proteins in high stocking density reared broiler chickens. Slov. Vet. Zb. 2019, 56, 129–138. [Google Scholar]
- Al-Aqil, A.; Zulkifli, I.; Bejo, M.H.; Sazili, A.; Rajion, M.; Somchit, M. Changes in heat shock protein 70, blood parameters, and fear-related behavior in broiler chickens as affected by pleasant and unpleasant human contact. Poult. Sci. 2013, 92, 33–40. [Google Scholar] [CrossRef]
- Xing, M.; Jin, X.; Wang, J.; Shi, Q.; Cai, J.; Xu, S. The antagonistic effect of selenium on lead-induced immune dysfunction via recovery of cytokine and heat shock protein expression in chicken neutrophils. Biol. Trace Elem. Res. 2018, 185, 162–169. [Google Scholar] [CrossRef]
- Huang, H.; Jiao, X.; Xu, Y.; Han, Q.; Jiao, W.; Liu, Y.; Li, S.; Teng, X. Dietary selenium supplementation alleviates immune toxicity in the hearts of chickens with lead-added drinking water. Avian Pathol. 2019, 48, 230–237. [Google Scholar] [CrossRef]
- Jiayong, Z.; Shengchen, W.; Xiaofang, H.; Gang, S.; Shiwen, X. The antagonistic effect of selenium on lead-induced necroptosis via MAPK/NF-kappaB pathway and HSPs activation in the chicken spleen. Ecotoxicol. Environ. Saf. 2020, 204, 111049. [Google Scholar] [CrossRef]
- Huang, H.; Wang, Y.; An, Y.; Tian, Y.; Li, S.; Teng, X. Selenium for the mitigation of toxicity induced by lead in chicken testes through regulating mRNA expressions of HSPs and selenoproteins. Environ. Sci. Pollut. Res. 2017, 24, 14312–14321. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, M.; Hou, L.; Lin, X.; Pan, S.; Zheng, P.; Zhao, Q. A potential mechanism associated with lead-induced spermatogonia and Leydig cell toxicity and mitigative effect of selenium in chicken. Ecotoxicol. Environ. Saf. 2021, 209, 111671. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, K.; Huang, H.; Gu, X.; Teng, X. Alleviative effect of selenium on inflammatory damage caused by lead via inhibiting inflammatory factors and heat shock proteins in chicken testes. Environ. Sci. Pollut. Res. Int. 2017, 24, 13405–13413. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.S.; Jin, H.T.; Talukder, M.; Ge, J.; Zhang, C.; Lv, M.W.; Yaqoob Ismail, M.A.; Li, J.L. The protective effect of nnano-selenium against cadmium-induced cerebellar injury via the heat shock protein pathway in chicken. Food Chem. Toxicol. 2021, 154, 112332. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, Y.H.; Cheng, X.Y.; Zhang, Z.W.; Xu, S.W. The protection of selenium against cadmium-induced cytotoxicity via the heat shock protein pathway in chicken splenic lymphocytes. Molecules 2012, 17, 14565–14572. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, M.; Cui, J.; Du, Y.; Teng, X.; Zhang, Z. Heat shock proteins took part in oxidative stress-mediated inflammatory injury via NF-kappaB pathway in excess manganese-treated chicken livers. Ecotoxicol. Environ. Saf. 2021, 226, 112833. [Google Scholar] [CrossRef]
- Zhao, P.; Guo, Y.; Zhang, W.; Chai, H.; Xing, H.; Xing, M. Neurotoxicity induced by arsenic in Gallus Gallus: Regulation of oxidative stress and heat shock protein response. Chemosphere 2017, 166, 238–245. [Google Scholar] [CrossRef]
- Zhao, P.; Zhang, K.; Guo, G.; Sun, X.; Chai, H.; Zhang, W.; Xing, M. Heat Shock Protein Alteration in the Gastrointestinal Tract Tissues of Chickens Exposed to Arsenic Trioxide. Biol. Trace Elem. Res. 2016, 170, 224–236. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, P.; Guo, G.; Hu, Z.; Tian, L.; Zhang, K.; Sun, Y.; Zhang, X.; Zhang, W.; Xing, M. Effects of Arsenic Trioxide Exposure on Heat Shock Protein Response in the Immune Organs of Chickens. Biol. Trace Elem. Res. 2016, 169, 134–141. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, P.; Guo, G.; Hu, Z.; Tian, L.; Zhang, K.; Zhang, W.; Xing, M. The Role of Oxidative Stress in Gastrointestinal Tract Tissues Induced by Arsenic Toxicity in Cocks. Biol. Trace Elem. Res. 2015, 168, 490–499. [Google Scholar] [CrossRef]
- Mackei, M.; Orban, K.; Molnar, A.; Pal, L.; Dublecz, K.; Husveth, F.; Neogrady, Z.; Matis, G. Cellular Effects of T-2 Toxin on Primary Hepatic Cell Culture Models of Chickens. Toxins 2020, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Li, Z.; Wang, G.; Yang, Z.; Yang, W.; Zhang, G.; Wu, Y. Effects of Fusarium mycotoxins with yeast cell wall absorbent on hematology, serum biochemistry, and oxidative stress in broiler chickens. J. Appl. Poult. Res. 2014, 23, 165–173. [Google Scholar] [CrossRef]
- Paraskeuas, V.; Griela, E.; Bouziotis, D.; Fegeros, K.; Antonissen, G.; Mountzouris, K.C. Effects of Deoxynivalenol and Fumonisins on Broiler Gut Cytoprotective Capacity. Toxins 2021, 13, 729. [Google Scholar] [CrossRef]
- Shukman, D. Climate: World at Risk of Hitting Temperature Limit Soon. Available online: https://www.bbc.com/news (accessed on 27 May 2021).
- Daghir, N.J. Poultry Production in Hot Climates, 2nd ed.; CABI Wallinford: Oxfordshire, UK, 2008; p. 387. [Google Scholar]
- Oguntunji, A.; Alabi, O. Influence of high environmental temperature on egg production and shell quality: A review. World Poult. Sci. J. 2010, 66, 739–750. [Google Scholar] [CrossRef]
- Yousaf, A.; Jabbar, A.; Rajput, N.; Memon, A.; Shahnawaz, R.; Mukhtar, N.; Farooq, F.; Abbas, M.; Khalil, R. Effect of environmental heat stress on performance and carcass yield of broiler chicks. World 2019, 9, 26–30. [Google Scholar] [CrossRef]
- Kelley, P.M.; Schlesinger, M.J. The effect of amino acid analogues and heat shock on gene expression in chicken embryo fibroblasts. Cell 1978, 15, 1277–1286. [Google Scholar] [CrossRef]
- Johnston, D.; Oppermann, H.; Jackson, J.; Levinson, W. Induction of four proteins in chick embryo cells by sodium arsenite. J. Biol. Chem. 1980, 255, 6975–6980. [Google Scholar] [CrossRef]
- Hightower, L.E. Cultured animal cells exposed to amino acid analogues or puromycin rapidly synthesize several polypeptides. J. Cell Physiol. 1980, 102, 407–427. [Google Scholar] [CrossRef]
- Wang, C.; Gomer, R.H.; Lazarides, E. Heat shock proteins are methylated in avian and mammalian cells. Proc. Natl. Acad. Sci. USA 1981, 78, 3531–3535. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-Y. Steroidal Modulation of Gene Expression for Heat-Shock Proteins in Domestic Chickens. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 1992. [Google Scholar]
- Edens, F.W.; Hill, C.H.; Wang, S. Heat shock protein response in phosphorus-deficient heat-stressed broiler chickens. Comp. Biochem. Physiol. B 1992, 103, 827–831. [Google Scholar] [CrossRef]
- Zhen, F.S.; Du, H.L.; Xu, H.P.; Luo, Q.B.; Zhang, X.Q. Tissue and allelic-specific expression of hsp70 gene in chickens: Basal and heat-stress-induced mRNA level quantified with real-time reverse transcriptase polymerase chain reaction. Br. Poult. Sci. 2006, 47, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Al-Aqil, A.; Zulkifli, I. Changes in heat shock protein 70 expression and blood characteristics in transported broiler chickens as affected by housing and early age feed restriction. Poult. Sci. 2009, 88, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.J.; Golic, K.G. Loss of Hsp70 in Drosophila is pleiotropic, with effects on thermotolerance, recovery from heat shock and neurodegeneration. Genetics 2006, 172, 275–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, J.E.; Ferro, J.A.; Stefani, R.M.; Ferro, M.I.; Gomes, S.L.; Macari, M. Effect of acute heat stress on heat shock protein 70 messenger RNA and on heat shock protein expression in the liver of broilers. Br. Poult. Sci. 1996, 37, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Bao, E. Effect of acute heat stress on heat shock protein 70 and its corresponding mRNA expression in the heart, liver, and kidney of broilers. Asian-Australas. J. Anim. Sci. 2008, 21, 1116–1126. [Google Scholar] [CrossRef]
- Mahmoud, K.Z.; Edens, F.W.; Eisen, E.J.; Havenstein, G.B. The effect of dietary phosphorus on heat shock protein mRNAs during acute heat stress in male broiler chickens (Gallus gallus). Comp. Biochem. Physiol. C Toxicol. Pharm. 2004, 137, 11–18. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B. Thermal manipulation during broiler chicken embryogenesis increases basal mRNA levels and alters production dynamics of heat shock proteins 70 and 60 and heat shock factors 3 and 4 during thermal stress. Poult. Sci. 2018, 97, 3661–3670. [Google Scholar] [CrossRef]
- Tang, S.; Zhou, S.; Yin, B.; Xu, J.; Di, L.; Zhang, J.; Bao, E. Heat stress-induced renal damage in poultry and the protective effects of HSP60 and HSP47. Cell Stress Chaperones 2018, 23, 1033–1040. [Google Scholar] [CrossRef]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Liu, Y.-T.; Wang, Q.-H.; Wang, Z.; Bao, E. Localizations of HSP70 and HSP70 mRNA in the tissues of heat stressed broilers. J. Agri. Biotech. 2007, 15, 404–408. [Google Scholar]
- Yu, J. The Mechanism of Heat Shock Protein Expressions and Tissue Damage in Heat Stressed Broilers. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2009; p. 133. [Google Scholar]
- McDaniel, C.D.; Bramwell, R.K.; Wilson, J.L.; Howarth, B., Jr. Fertility of male and female broiler breeders following exposure to elevated ambient temperatures. Poult. Sci. 1995, 74, 1029–1038. [Google Scholar] [CrossRef]
- Karaca, A.G.; Parker, H.M.; Yeatman, J.B.; McDaniel, C.D. Role of seminal plasma in heat stress infertility of broiler breeder males. Poult. Sci. 2002, 81, 1904–1909. [Google Scholar] [CrossRef]
- Oehler, R.; Pusch, E.; Zellner, M.; Dungel, P.; Hergovics, N.; Homoncik, M.; Eliasen, M.M.; Brabec, M.; Roth, E. Cell type-specific variations in the induction of hsp70 in human leukocytes by feverlike whole body hyperthermia. Cell Stress Chaperones 2001, 6, 306–315. [Google Scholar] [CrossRef]
- Yahav, S.; Rath, R.S.; Shinder, D. The effect of thermal manipulations during embryogenesis of broiler chicks (Gallus domesticus) on hatchability, body weight and thermoregulation after hatch. J. Therm. Biol. 2004, 29, 245–250. [Google Scholar] [CrossRef]
- Piestun, Y.; Harel, M.; Barak, M.; Yahav, S.; Halevy, O. Thermal manipulations in late-term chick embryos have immediate and longer term effects on myoblast proliferation and skeletal muscle hypertrophy. J. Appl. Physiol. 2009, 106, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Willemsen, H.; Kamers, B.; Dahlke, F.; Han, H.; Song, Z.; Ansari Pirsaraei, Z.; Tona, K.; Decuypere, E.; Everaert, N. High- and low-temperature manipulation during late incubation: Effects on embryonic development, the hatching process, and metabolism in broilers. Poult. Sci. 2010, 89, 2678–2690. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Animal Physiology: Adaptation and Environment; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Vinoth, A.; Thirunalasundari, T.; Shanmugam, M.; Uthrakumar, A.; Suji, S.; Rajkumar, U. Evaluation of DNA methylation and mRNA expression of heat shock proteins in thermal manipulated chicken. Cell Stress Chaperones 2018, 23, 235–252. [Google Scholar] [CrossRef]
- Liew, P.K.; Zulkifli, I.; Hair-Bejo, M.; Omar, A.R.; Israf, D.A. Effects of early age feed restriction and heat conditioning on heat shock protein 70 expression, resistance to infectious bursal disease, and growth in male broiler chickens subjected to heat stress. Poult. Sci. 2003, 82, 1879–1885. [Google Scholar] [CrossRef]
- Yahav, S.; Shamai, A.; Haberfeld, A.; Horev, G.; Hurwitz, S.; Einat, M. Induction of thermotolerance in chickens by temperature conditioning: Heat shock protein expression. Ann. N. Y. Acad. Sci. 1997, 813, 628–636. [Google Scholar] [CrossRef]
- Wang, S.; Edens, F.W. Heat-stress response of broiler cockerels to manipulation of the gonadal steroids, testosterone and estradiol. Comp. Biochem. Physiol. B 1993, 106, 629–633. [Google Scholar] [CrossRef]
- Wang, S.; Edens, F.W. Heat conditioning induces heat shock proteins in broiler chickens and turkey poults. Poult. Sci. 1998, 77, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Hanafi, S.A.; Zulkifli, I.; Ramiah, S.K.; Chung, E.L.T.; Awad, E.A.; Sazili, A.Q. Prenatal thermal stress affects acute-phase proteins, heat shock protein 70, and circulating corticosterone in developing broiler embryos and neonates. J. Therm. Biol. 2022, 109, 103328. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.J.; Kong, L.L.; Zhu, L.X.; Hu, X.Y.; Busye, J.; Song, Z.G. Effects of cold stress on growth performance, serum biochemistry, intestinal barrier molecules, and adenosine monophosphate-activated protein kinase in broilers. Animal 2021, 15, 100138. [Google Scholar] [CrossRef] [PubMed]
- Hangalapura, B.N.; Nieuwland, M.G.; de Vries Reilingh, G.; Heetkamp, M.J.; van den Brand, H.; Kemp, B.; Parmentier, H.K. Effects of cold stress on immune responses and body weight of chicken lines divergently selected for antibody responses to sheep red blood cells. Poult. Sci. 2003, 82, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. The effect of cold stress on the pathogenesis of necrotic enteritis in broiler chicks. Avian. Pathol. 2015, 44, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Q.; Zhang, Z.W.; Qu, J.P.; Yao, H.D.; Li, M.; Li, S.; Xu, S.W. Cold stress induces antioxidants and Hsps in chicken immune organs. Cell Stress Chaperones 2014, 19, 635–648. [Google Scholar] [CrossRef]
- Huff, G.R.; Huff, W.E.; Rath, N.C.; Solis de Los Santos, F.; Farnell, M.B.; Donoghue, A.M. Influence of hen age on the response of turkey poults to cold stress, Escherichia coli challenge, and treatment with a yeast extract antibiotic alternative. Poult. Sci. 2007, 86, 636–642. [Google Scholar] [CrossRef]
- Aminoroaya, K.; Sadeghi, A.A.; Ansari-Pirsaraei, Z.; Kashan, N. Effect of cyclical cold stress during embryonic development on aspects of physiological responses and HSP70 gene expression of chicks. J. Therm. Biol. 2016, 61, 50–54. [Google Scholar] [CrossRef]
- Buhr, R.J.; Bourassa, D.V.; Hinton, A., Jr.; Fairchild, B.D.; Ritz, C.W. Impact of litter Salmonella status during feed withdrawal on Salmonella recovery from the broiler crop and ceca. Poult. Sci. 2017, 96, 4361–4369. [Google Scholar] [CrossRef]
- Dawkins, M.S. From an animal’s point of view: Motivation, fitness, and animal welfare. Behav. Brain Sci. 1990, 13, 1–9. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Hair-Bejo, M.; Omar, A.R.; Raha, A.R. The role of heat shock protein 70 in resistance to Salmonella enteritidis in broiler chickens subjected to neonatal feed restriction and thermal stress. Poult. Sci. 2012, 91, 340–345. [Google Scholar] [CrossRef]
- Sohn, S.; Subramani, V.; Moon, Y.; Jang, I. Telomeric DNA quantity, DNA damage, and heat shock protein gene expression as physiological stress markers in chickens. Poult. Sci. 2012, 91, 829–836. [Google Scholar] [CrossRef]
- Zulkifli, I. Review of human-animal interactions and their impact on animal productivity and welfare. J. Anim. Sci. Biotech. 2013, 4, 1–7. [Google Scholar] [CrossRef]
- Lyons, D.M.; Parker, K.J.; Schatzberg, A.F. Animal models of early life stress: Implications for understanding resilience. Dev. Psychobiol. 2010, 52, 402–410. [Google Scholar] [CrossRef]
- Zulkifli, I.; Dunnington, E.A.; Gross, W.B.; Siegel, P.B. Food restriction early or later in life and its effect on adaptability, disease resistance, and immunocompetence of heat-stressed dwarf and nondwarf chickens. Br. Poult. Sci. 1994, 35, 203–213. [Google Scholar] [CrossRef]
- Zulkifli, I.; Dunnington, E.A.; Gross, W.B.; Siegel, P.B. Inhibition of adrenal steroidogenesis, food restriction and acclimation to high ambient temperatures in chickens. Br. Poult. Sci. 1994, 35, 417–426. [Google Scholar] [CrossRef]
- Zulkifli, I.; Che Norma, M.T.; Israf, D.A.; Omar, A.R. The effect of early age feed restriction on subsequent response to high environmental temperatures in female broiler chickens. Poult. Sci. 2000, 79, 1401–1407. [Google Scholar] [CrossRef]
- Craig, E.A. The heat shock respons. Crit. Rev. Biochem. 1985, 18, 239–280. [Google Scholar] [CrossRef]
- Zulkifli, I.; Najafi, P.; Nurfarahin, A.; Soleimani, A.; Kumari, S.; Aryani, A.A.; O’Reilly, E.; Eckersall, P. Acute phase proteins, interleukin 6, and heat shock protein 70 in broiler chickens administered with corticosterone. Poult. Sci. 2014, 93, 3112–3118. [Google Scholar] [CrossRef]
- Terlouw, E.M.; Arnould, C.; Auperin, B.; Berri, C.; Le Bihan-Duval, E.; Deiss, V.; Lefevre, F.; Lensink, B.J.; Mounier, L. Pre-slaughter conditions, animal stress and welfare: Current status and possible future research. Animal 2008, 2, 1501–1517. [Google Scholar] [CrossRef] [Green Version]
- Benincasa, N.C.; Sakamoto, K.S.; da Silva, I.J.O.; Lobos, C.M.V. Animal welfare: Impacts of pre-slaughter operations on the current poultry industry. J. Anim. Beh. Biometeorol. 2020, 8, 104–110. [Google Scholar] [CrossRef]
- Saraiva, S.; Esteves, A.; Oliveira, I.; Mitchell, M.; Stilwell, G. Impact of pre-slaughter factors on welfare of broilers. Vet. Anim. Sci. 2020, 10, 100146. [Google Scholar] [CrossRef] [PubMed]
- Kannan, G.; Mench, J.A. Influence of different handling methods and crating periods on plasma corticosterone concentrations in broilers. Br. Poult. Sci. 1996, 37, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Che Norma, M.T.; Chong, C.H.; Loh, T.C. Heterophil to lymphocyte ratio and tonic immobility reactions to preslaughter handling in broiler chickens treated with ascorbic acid. Poult. Sci. 2000, 79, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Che Norma, M.T.; Chong, C.H.; Loh, T.C. The effects of crating and road transportation on stress and fear responses of broiler chickens treated with ascorbic acid. Arch. Fur. Geflugelkd. 2001, 65, 33–37. [Google Scholar]
- Duncan, I. The assessment of welfare during the handling and transport of broilers. In Proceedings of the 3rd European Symposium on Poultry Welfare; World’s Poultry Science Association: Tours, France, 1989; pp. 93–107. [Google Scholar]
- Yubao, L.; Lanying, P.; Endong, B. Relationship between distribution, transcription level of HSPs mRNA and immunity tissue pathological lesion of transport stressed pigs. Sci. Agric. Sin. 2008, 46, 1832–1837. [Google Scholar]
- Lentfer, T.L.; Gebhardt-Henrich, S.G.; Fröhlich, E.K.; von Borell, E. Influence of nest site on the behaviour of laying hens. Appl. Anim. Behav. Sci. 2011, 135, 70–77. [Google Scholar] [CrossRef]
- Rodriguez-Aurrekoetxea, A.; Estevez, I. Aggressiveness in the domestic fowl: Distance versus ‘attitude’. Appl. Anim. Behav. Sci. 2014, 153, 68–74. [Google Scholar] [CrossRef]
- Widowski, T.M.; Caston, L.J.; Hunniford, M.E.; Cooley, L.; Torrey, S. Effect of space allowance and cage size on laying hens housed in furnished cages, Part I: Performance and well-being. Poult. Sci. 2017, 96, 3805–3815. [Google Scholar] [CrossRef]
- Clark, J.A. Environmental aspects of housing for animal production; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Young, J.F.; Stagsted, J.; Jensen, S.K.; Karlsson, A.H.; Henckel, P. Ascorbic acid, alpha-tocopherol, and oregano supplements reduce stress-induced deterioration of chicken meat quality. Poult. Sci. 2003, 82, 1343–1351. [Google Scholar] [CrossRef]
- Estevez, I. Density allowances for broilers: Where to set the limits? Poult. Sci. 2007, 86, 1265–1272. [Google Scholar] [CrossRef]
- Law, F.; Zulkifli, I.; Soleimani, A.; Liang, J.; Awad, E. Effects of reduced-protein diets supplemented with protease in broiler chickens under high stocking density. Anim. Prod. Sci. 2019, 59, 2212–2221. [Google Scholar] [CrossRef]
- Puron, D.; Santamaria, R.; Segura, J.C.; Alamilla, J.L. Broiler performance at different stocking densities. J. Appl. Poult. Res. 1995, 4, 55–60. [Google Scholar] [CrossRef]
- Thaxton, J.P.; Dozier, W.A., 3rd; Branton, S.L.; Morgan, G.W.; Miles, D.W.; Roush, W.B.; Lott, B.D.; Vizzier-Thaxton, Y. Stocking density and physiological adaptive responses of broilers. Poult. Sci. 2006, 85, 819–824. [Google Scholar] [CrossRef]
- Imaeda, N. Influence of the stocking density and rearing season on incidence of sudden death syndrome in broiler chickens. Poult. Sci. 2000, 79, 201–204. [Google Scholar] [CrossRef]
- Mahmoud, K.Z.; Edens, F.W.; Eisen, E.J.; Havenstein, G.B. Ascorbic acid decreases heat shock protein 70 and plasma corticosterone response in broilers (Gallus gallus domesticus) subjected to cyclic heat stress. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2004, 137, 35–42. [Google Scholar] [CrossRef]
- Sahin, N.; Tuzcu, M.; Orhan, C.; Onderci, M.; Eroksuz, Y.; Sahin, K. The effects of vitamin C and E supplementation on heat shock protein 70 response of ovary and brain in heat-stressed quail. Br. Poult. Sci. 2009, 50, 259–265. [Google Scholar] [CrossRef]
- Suarez, S.D.; Gallup, G.G. Open-field behavior in chickens: The experimenter is a predator. J. Comp. Physiol. Psychol. 1982, 96, 432. [Google Scholar] [CrossRef]
- Jones, R.B. Fear and adaptability in poultry: Insights, implications and imperatives. World’s Poult. Sci. J. 1996, 52, 131–174. [Google Scholar] [CrossRef]
- Zulkifli, I.; Azah, A.S.N. Fear and stress reactions, and the performance of commercial broiler chickens subjected to regular pleasant and unpleasant contacts with human being. Appl. Anim. Behav. Sci. 2004, 88, 77–87. [Google Scholar] [CrossRef]
- Zulkifli, I.; Gilbert, J.; Liew, P.; Ginsos, J. The effects of regular visual contact with human beings on fear, stress, antibody and growth responses in broiler chickens. Appl. Anim. Behav. Sci. 2002, 79, 103–112. [Google Scholar] [CrossRef]
- Hemsworth, P.; Coleman, G.; Barnett, J.; Jones, R. Behavioural responses to humans and the productivity of commercial broiler chickens. Appl. Anim. Behav. Sci. 1994, 41, 101–114. [Google Scholar] [CrossRef]
- Hemsworth, P. Human-livestock interaction. In The Well-Being of Farm Animals, Challenges and Solutions; Benson, G.J., Rollin, B.E., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2004. [Google Scholar]
- Hemsworth, P.H. Ethical stockmanship. Aust. Vet. J. 2007, 85, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.M. The chronic toxicity of aluminium, cadmium, mercury, and lead in birds: A review. Environ. Pollut. 1987, 46, 263–295. [Google Scholar] [CrossRef] [PubMed]
- Bernstam, L.; Nriagu, J. Molecular aspects of arsenic stress. J. Toxicol. Environ. Health B Crit. Rev. 2000, 3, 293–322. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, C.; Ge, J.; Lv, M.W.; Talukder, M.; Guo, K.; Li, Y.H.; Li, J.L. Ameliorative effects of resveratrol against cadmium-induced nephrotoxicity via modulating nuclear xenobiotic receptor response and PINK1/Parkin-mediated Mitophagy. Food Funct. 2020, 11, 1856–1868. [Google Scholar] [CrossRef]
- Liu, X.; Song, J.; Zheng, Z.; Guan, H.; Nan, X.; Zhang, N. Effects of Excess Manganese on the Oxidative Status, and the Expression of Inflammatory Factors and Heat Shock Proteins in Cock Kidneys. Biol. Trace. Elem. Res. 2020, 197, 639–650. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Phillips, T.D.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef]
- Hassen, W.; El Golli, E.; Baudrimont, I.; Mobio, A.T.; Ladjimi, M.M.; Creppy, E.E.; Bacha, H. Cytotoxicity and Hsp 70 induction in Hep G2 cells in response to zearalenone and cytoprotection by sub-lethal heat shock. Toxicology 2005, 207, 293–301. [Google Scholar] [CrossRef]
- El Golli, E.; Hassen, W.; Bouslimi, A.; Bouaziz, C.; Ladjimi, M.M.; Bacha, H. Induction of Hsp 70 in Vero cells in response to mycotoxins cytoprotection by sub-lethal heat shock and by Vitamin E. Toxicol. Lett. 2006, 166, 122–130. [Google Scholar] [CrossRef]
- Shang, Q.H.; Yang, Z.B.; Yang, W.R.; Li, Z.; Zhang, G.G.; Jiang, S.Z. Toxicity of Mycotoxins from Contaminated Corn with or withoutYeast Cell Wall Adsorbent on Broiler Chickens. Asian-Australas. J. Anim. Sci. 2016, 29, 674–680. [Google Scholar] [CrossRef] [Green Version]
- Zulkifli, I.; Abdulllah, N.; Azrin, N.M.; Ho, Y.W. Growth performance and immune response of two commercial broiler strains fed diets containing Lactobacillus cultures and oxytetracycline under heat stress conditions. Br. Poult. Sci. 2000, 41, 593–597. [Google Scholar] [CrossRef]
- Aarseth, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.C.; Yan, F.F.; Hu, J.Y.; Amen, O.A.; Cheng, H.W. Supplementation of Bacillus subtilis-based probiotic reduces heat stress-related behaviors and inflammatory response in broiler chickens. J. Anim. Sci. 2018, 96, 1654–1666. [Google Scholar] [CrossRef]
- Jiang, S.; Mohammed, A.A.; Jacobs, J.A.; Cramer, T.A.; Cheng, H.W. Effect of synbiotics on thyroid hormones, intestinal histomorphology, and heat shock protein 70 expression in broiler chickens reared under cyclic heat stress. Poult. Sci. 2020, 99, 142–150. [Google Scholar] [CrossRef]
- Awad, E.; Zulkifli, I.; Ramiah, S.; Khalil, E.; Abdallh, M. Prebiotics supplementation: An effective approach to mitigate the detrimental effects of heat stress in broiler chickens. World Poult. Sci. J. 2021, 77, 135–151. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.; Nitsch, S.; Abdel-Raheem, S.; Böhm, J. Effects of transportation on stress and fear responses of growing broilers supplemented with prebiotic or probiotic. Int. J. Poult. Sci. 2008, 7, 678–685. [Google Scholar] [CrossRef]
- Ghareeb, K.; Böhm, J. Stress indicators to pre-slaughter transportation of broiler chickens fed diets supplemented with a synbiotic. Int. J. Poult. Sci. 2009, 8, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Bello, U.A.; Idrus, Z.; Yong Meng, G.; Awad, E.A.; Soleimani Farjam, A. Gut microbiota and transportation stress response affected by tryptophan supplementation in broiler chickens. Ital. J. Anim. Sci. 2018, 17, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.M.; Nazarizadeh, H.; Ahani, S.; Vakili Azghandi, M. Effects of mannan oligosaccharide and Curcuma xanthorrhiza essential oil on the intestinal morphologyand stress indicators of broilers subjected to cyclic heat stress. Arch. Anim. Breed. 2016, 59, 285–291. [Google Scholar] [CrossRef]
- Rokade, J.; Kagate, M.; Bhanja, S.; Mehra, M.; Goel, A.; Vispute, M.; Mandal, A. Effect of mannan-oligosaccharides (MOS) supplementation on performance, immunity and HSP70 gene expression in broiler chicken during hot-dry summer. Indian J. Anim. Res. 2018, 52, 868–874. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Ho, Y.; Abdullah, N.; Jalaaudin, S. Probiotic in Poultry, modes of action. World Poult. Sci. J. 1997, 53, N4. [Google Scholar] [CrossRef]
- Zhang, P.; Yan, T.; Wang, X.; Kuang, S.; Xiao, Y.; Lu, W.; Bi, D. Probiotic mixture ameliorates heat stress of laying hens by enhancing intestinal barrier function and improving gut microbiota. Ital. J. Anim. Sci. 2017, 16, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.Z.; Kumbhar, S.; Hamid, M.; Afzal, S.; Parveen, F.; Liu, Y.; Shu, H.; Mengistu, B.M.; Huang, K. Effects of Selenium-Enriched Probiotics on Heart Lesions by Influencing the mRNA Expressions of Selenoproteins and Heat Shock Proteins in Heat Stressed Broiler Chickens. Pak. Vet. J. 2016, 36, 460–464. [Google Scholar]
- Khan, A.Z.; Kumbhar, S.; Liu, Y.; Hamid, M.; Pan, C.; Nido, S.A.; Parveen, F.; Huang, K. Dietary Supplementation of Selenium-Enriched Probiotics Enhances Meat Quality of Broiler Chickens (Gallus gallus domesticus) Raised Under High Ambient Temperature. Biol. Trace Elem. Res. 2018, 182, 328–338. [Google Scholar] [CrossRef]
- Mohammed, A.A.; Jacobs, J.A.; Murugesan, G.R.; Cheng, H.W. Effect of dietary synbiotic supplement on behavioral patterns and growth performance of broiler chickens reared under heat stress. Poult. Sci. 2018, 97, 1101–1108. [Google Scholar] [CrossRef]
- Hu, J.; Mohammed, A.; Murugesan, G.; Cheng, H. Effect of a synbiotic supplement as an antibiotic alternative on broiler skeletal, physiological, and oxidative parameters under heat stress. Poult. Sci. 2022, 101, 101769–101780. [Google Scholar] [CrossRef]
- Tan, G.Y.; Yang, L.; Fu, Y.Q.; Feng, J.H.; Zhang, M.H. Effects of different acute high ambient temperatures on function of hepatic mitochondrial respiration, antioxidative enzymes, and oxidative injury in broiler chickens. Poult. Sci. 2010, 89, 115–122. [Google Scholar] [CrossRef]
- Mujahid, A.; Akiba, Y.; Warden, C.H.; Toyomizu, M. Sequential changes in superoxide production, anion carriers and substrate oxidation in skeletal muscle mitochondria of heat-stressed chickens. FEBS Lett. 2007, 581, 3461–3467. [Google Scholar] [CrossRef] [Green Version]
- Meydani, M.; Evans, W.; Handelman, G.; Fielding, R.A.; Meydani, S.N.; Fiatarone, M.A.; Blumberg, J.B.; Cannon, J.G. Antioxidant response to exercise-induced oxidative stress and protection by vitamin E. Ann. N. Y. Acad. Sci. 1992, 669, 363–364. [Google Scholar] [CrossRef]
- Puthpongsiriporn, U.; Scheideler, S.E.; Sell, J.L.; Beck, M.M. Effects of vitamin E and C supplementation on performance, in vitro lymphocyte proliferation, and antioxidant status of laying hens during heat stress. Poult. Sci. 2001, 80, 1190–1200. [Google Scholar] [CrossRef]
- Traber, M.G.; Stevens, J.F. Vitamins C and E: Beneficial effects from a mechanistic perspective. Free Radic. Biol. Med. 2011, 51, 1000–1013. [Google Scholar] [CrossRef] [Green Version]
- Attia, Y.A.; Al-Harthi, M.A.; El-Shafey, A.S.; Rehab, Y.A.; Kim, W.K. Enhancing tolerance of broiler chickens to heat stress by supplementation with vitamin E, vitamin C and/or probiotics. Ann. Anim. Sci. 2017, 17, 1155. [Google Scholar] [CrossRef] [Green Version]
- Jang, I.S.; Ko, Y.H.; Moon, Y.S.; Sohn, S.H. Effects of Vitamin C or E on the Pro-inflammatory Cytokines, Heat Shock Protein 70 and Antioxidant Status in Broiler Chicks under Summer Conditions. Asian-Australas. J. Anim. Sci. 2014, 27, 749–756. [Google Scholar] [CrossRef]
- Cheng, T.K.; Hamre, M.L.; Coon, C.N. Effect of constant and cyclic environmental temperatures, dietary protein, and amino acid levels on broiler performance. J. Appl. Poult. Res. 1999, 8, 426–439. [Google Scholar] [CrossRef]
- Faria Filho, D.; Rosa, P.; Vieira, B.; Macari, M.; Furlan, R.L. Protein levels and environmental temperature effects on carcass characteristics, performance, and nitrogen excretion of broiler chickens from 7 to 21 days of age. Braz. J. Poult. Sci. 2005, 7, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Esquerra, R.; Leeson, S. Effects of acute versus chronic heat stress on broiler response to dietary protein. Poult. Sci. 2005, 84, 1562–1569. [Google Scholar] [CrossRef]
- Awad, E.A.; Zulkifli, I.; Farjam, A.S.; Chwen, L.T. Amino acids fortification of low-protein diet for broilers under tropical climate. 2. Nonessential amino acids and increasing essential amino acids. Ital. J. Anim. Sci. 2014, 13, 3297. [Google Scholar] [CrossRef]
- Zulkifli, I.; Akmal, A.F.; Soleimani, A.F.; Hossain, M.A.; Awad, E.A. Effects of low-protein diets on acute phase proteins and heat shock protein 70 responses, and growth performance in broiler chickens under heat stress condition. Poult. Sci. 2018, 97, 1306–1314. [Google Scholar] [CrossRef]
- Awad, E.; Zulkifli, I.; Soleimani, A.; Law, F.; Ramiah, S.; Mohamed-Yousif, I.; Hussein, E.; Khalil, E. Response of broilers to reduced-protein diets under heat stress conditions. World Poult. Sci. J. 2019, 75, 583–598. [Google Scholar] [CrossRef]
- Berres, J.; Vieira, S.; Dozier Iii, W.; Cortês, M.; De Barros, R.; Nogueira, E.; Kutschenko, M. Broiler responses to reduced-protein diets supplemented with valine, isoleucine, glycine, and glutamic acid. J. Appl. Poult. Res. 2010, 19, 68–79. [Google Scholar] [CrossRef]
- Kriseldi, R.; Tillman, P.; Jiang, Z.; Dozier III, W. Effects of glycine and glutamine supplementation to reduced crude protein diets on growth performance and carcass characteristics of male broilers during a 41-day production period. Jf. Appl. Poult. Res. 2017, 26, 558–572. [Google Scholar] [CrossRef]
- Lee, D.T.; Lee, J.T.; Ruan, C.; Rochell, S.J. Influence of increasing glycine concentrations in reduced crude protein diets fed to broilers from 0 to 48 days. Poult. Sci. 2022, 101, 102038. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.G.; Stoll, B. Metabolic fate and function of dietary glutamate in the gut. Am. J. Clin. Nutr. 2009, 90, 850S–856S. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.F.; Wang, L.K.; Wen, A.Y.; Wang, L.X.; Jin, G.M. Dietary glutamine supplementation improves growth performance, meat quality and colour stability of broilers under heat stress. Br. Poult. Sci. 2009, 50, 333–340. [Google Scholar] [CrossRef]
- Dai, S.; Gao, F.; Zhang, W.; Song, S.; Xu, X.; Zhou, G. Effects of dietary glutamine and gamma-aminobutyric acid on performance, carcass characteristics and serum parameters in broilers under circular heat stress. Anim. Feed. Sci. Technol. 2011, 168, 51–60. [Google Scholar] [CrossRef]
- Jazideh, F.; Farhoomand, P.; Daneshyar, M.; Najafi, G. The effects of dietary glutamine supplementation on growth performance and intestinal morphology of broiler chickens reared under hot conditions. Turk. J. Vet. Anim. Sci. 2014, 38, 264–270. [Google Scholar] [CrossRef]
- Morrison, A.L.; Dinges, M.; Singleton, K.D.; Odoms, K.; Wong, H.R.; Wischmeyer, P.E. Glutamine’s protection against cellular injury is dependent on heat shock factor-1. Am. J. Physiol-Cell Physiol. 2006, 290, C1625–C1632. [Google Scholar] [CrossRef]
- Olubodun, J.; Zulkifli, I.; Hair-Bejo, M.; Kasim, A.; Soleimani, A. Physiological response of glutamine and glutamic acid supplemented broiler chickens to heat stress. Eur. Poult. Sci. 2015, 79, 1–12. [Google Scholar]
- Hu, H.; Chen, L.; Dai, S.; Li, J.; Bai, X. Effect of Glutamine on Antioxidant Capacity and Lipid Peroxidation in the Breast Muscle of Heat-stressed Broilers via Antioxidant Genes and HSP70 Pathway. Animals 2020, 10, 404. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, M.; Zulkifli, İ.; Oskoueian, E.; Shakeri, M.; Oskoueian, A.; Ebrahimi, M. Response to dietary supplementation of glutamine in broiler chickens subjected to transportation stress. İstanbul Üniversitesi Vet. Fakültesi Derg. 2016, 42, 122–131. [Google Scholar] [CrossRef]
- Wu, G. Intestinal mucosal amino acid catabolism. J. Nutr. 1998, 128, 1249–1252. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336 Pt 1, 1–17. [Google Scholar] [CrossRef]
- Olubodun, J.O.; Zulkifli, I.; Farjam, A.S.; Hair-Bejo, M.; Kasim, A. Glutamine and glutamic acid supplementation enhances performance of broiler chickens under the hot and humid tropical condition. Ital. J. Anim. Sci. 2015, 14, 3263. [Google Scholar] [CrossRef]
- Hamiel, C.R.; Pinto, S.; Hau, A.; Wischmeyer, P.E. Glutamine enhances heat shock protein 70 expression via increased hexosamine biosynthetic pathway activity. Am. J. Physiol. Cell. Physiol. 2009, 297, C1509–C1519. [Google Scholar] [CrossRef] [Green Version]
- Rafiei, F.; Khajali, F. Flavonoid antioxidants in chicken meat production: Potential application and future trends. World Poult. Sci. J. 2021, 77, 347–361. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharm. Rev. 2000, 52, 673–751. [Google Scholar]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef]
- Hosokawa, N.; Hirayoshi, K.; Kudo, H.; Takechi, H.; Aoike, A.; Kawai, K.; Nagata, K. Inhibition of the activation of heat shock factor in vivo and in vitro by flavonoids. Mol. Cell Biol. 1992, 12, 3490–3498. [Google Scholar] [CrossRef]
- Sugito, S.; Etriwati, E.; Akmal, M.; Rahmi, E.; Delima, M.; Muchlisin, Z.A.; Hasan, D.I. Immunohistochemical Expression of AQP2 and HSP70 in Broiler Kidney Tissue Treated with Salix tetrasperma Roxb. Extract under Heat Exposure. Sci. World J. 2021, 2021, 8711286. [Google Scholar] [CrossRef] [PubMed]
- Elia, G.; Santoro, M.G. Regulation of heat shock protein synthesis by quercetin in human erythroleukaemia cells. Biochem. J. 1994, 300 Pt 1, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Liu, Y.; Cheng, X.; Qiao, X. The Alleviative Effects of Quercetin on Cadmium-Induced Necroptosis via Inhibition ROS/iNOS/NF-kappaB Pathway in the Chicken Brain. Biol. Trace Elem. Res. 2021, 199, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Kamboh, A.A.; Hang, S.Q.; Bakhetgul, M.; Zhu, W.Y. Effects of genistein and hesperidin on biomarkers of heat stress in broilers under persistent summer stress. Poult. Sci. 2013, 92, 2411–2418. [Google Scholar] [CrossRef] [PubMed]
- Hajati, H.; Hassanabadi, A.; Golian, A.; Nassiri-Moghaddam, H.; Nassiri, M.R. The effect of grape seed extract and vitamin C feed supplementation on some blood parameters and HSP70 gene expression of broiler chickens suffering from chronic heat stress. Ital. J. Anim. Sci. 2015, 14, 3273. [Google Scholar] [CrossRef]
- Sun, J.; Jiao, Z.; Zhu, W.; Li, X.; Wang, P.; Wang, J.; Tai, T.; Wang, Y.; Wang, H.; Shi, G. Astilbin Attenuates Cadmium-Induced Adipose Tissue Damage by Inhibiting NF-kappaB Pathways and Regulating the Expression of HSPs in Chicken. Biol. Trace. Elem. Res. 2022, 18, 1–2. [Google Scholar] [CrossRef]
- Sahin, K.; Akdemir, F.; Orhan, C.; Tuzcu, M.; Hayirli, A.; Sahin, N. Effects of dietary resveratrol supplementation on egg production and antioxidant status. Poult. Sci. 2010, 89, 1190–1198. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Akdemir, F.; Tuzcu, M.; Iben, C.; Sahin, N. Resveratrol protects quail hepatocytes against heat stress: Modulation of the Nrf2 transcription factor and heat shock proteins. J. Anim. Physiol. Anim. Nutr. 2012, 96, 66–74. [Google Scholar] [CrossRef]
- Liu, L.L.; He, J.H.; Xie, H.B.; Yang, Y.S.; Li, J.C.; Zou, Y. Resveratrol induces antioxidant and heat shock protein mRNA expression in response to heat stress in black-boned chickens. Poult. Sci. 2014, 93, 54–62. [Google Scholar] [CrossRef]
- Liu, L.; Fu, C.; Yan, M.; Xie, H.; Li, S.; Yu, Q.; He, S.; He, J. Resveratrol modulates intestinal morphology and HSP70/90, NF-kappaB and EGF expression in the jejunal mucosa of black-boned chickens on exposure to circular heat stress. Food Funct. 2016, 7, 1329–1338. [Google Scholar] [CrossRef]
- Hu, R.; He, Y.; Arowolo, M.A.; Wu, S.; He, J. Polyphenols as potential attenuators of heat stress in poultry production. Antioxidants 2019, 8, 67. [Google Scholar] [CrossRef]
- Zhang, J.F.; Bai, K.W.; Su, W.P.; Wang, A.A.; Zhang, L.L.; Huang, K.H.; Wang, T. Curcumin attenuates heat-stress-induced oxidant damage by simultaneous activation of GSH-related antioxidant enzymes and Nrf2-mediated phase II detoxifying enzyme systems in broiler chickens. Poult. Sci. 2018, 97, 1209–1219. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Z.; Lu, C.; Yang, M.; Zhang, L.; Wang, T. Dietary curcumin supplementation protects against heat-stress-impaired growth performance of broilers possibly through a mitochondrial pathway. J. Anim. Sci. 2015, 93, 1656–1665. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.w.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin attenuates hepatic mitochondrial dysfunction through the maintenance of thiol pool, inhibition of mtDNA damage, and stimulation of the mitochondrial thioredoxin system in heat-stressed broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef]
- Akhavan-Salamat, H.; Ghasemi, H.A. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: Dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop. Anim. Health Prod. 2016, 48, 181–188. [Google Scholar] [CrossRef]
- Hasheimi, S.R.; Zulkifli, I.; Somchit, M.N.; Zunita, Z.; Loh, T.C.; Soleimani, A.F.; Tang, S.C. Dietary supplementation of Zingiber officinale and Zingiber zerumbet to heat-stressed broiler chickens and its effect on heat shock protein 70 expression, blood parameters and body temperature. J. Anim. Physiol. Anim. Nutr. 2013, 97, 632–638. [Google Scholar] [CrossRef]
- Mack, L.A.; Felver-Gant, J.N.; Dennis, R.L.; Cheng, H.W. Genetic variations alter production and behavioral responses following heat stress in 2 strains of laying hens. Poult. Sci. 2013, 92, 285–294. [Google Scholar] [CrossRef]
- Wang, Y.; Saelao, P.; Chanthavixay, K.; Gallardo, R.; Bunn, D.; Lamont, S.J.; Dekkers, J.M.; Kelly, T.; Zhou, H. Physiological responses to heat stress in two genetically distinct chicken inbred lines. Poult. Sci. 2018, 97, 770–780. [Google Scholar] [CrossRef]
- Awad, E.A.; Najaa, M.; Zulaikha, Z.A.; Zulkifli, I.; Soleimani, A.F. Effects of heat stress on growth performance, selected physiological and immunological parameters, caecal microflora, and meat quality in two broiler strains. Asian-Australas. J. Anim. Sci. 2020, 33, 778–787. [Google Scholar] [CrossRef]
- Adu-Asiamah, P.; Zhang, Y.; Amoah, K.; Leng, Q.Y.; Zheng, J.H.; Yang, H.; Zhang, W.L.; Zhang, L. Evaluation of physiological and molecular responses to acute heat stress in two chicken breeds. Animal 2021, 15, 100106. [Google Scholar] [CrossRef]
- Heck, A.; Onagbesan, O.; Tona, K.; Metayer, S.; Putterflam, J.; Jego, Y.; Trevidy, J.J.; Decuypere, E.; Williams, J.; Picard, M.; et al. Effects of ad libitum feeding on performance of different strains of broiler breeders. Br. Poult. Sci. 2004, 45, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.D.; Mirò, S.M.; Tarantola, M.; Bergagna, S.; Mellia, E.; Gennero, M.S.; Schiavone, A. Effect of genotype and transport on tonic immobility and heterophil/lymphocyte ratio in two local Italian breeds and Isa Brown hens kept under free-range conditions. Ital. J. Anim. Sci. 2013, 12, e78. [Google Scholar] [CrossRef]
- Peixoto, M.; Karrow, N.A.; Newman, A.; Widowski, T.M. Effects of Maternal Stress on Measures of Anxiety and Fearfulness in Different Strains of Laying Hens. Front. Vet. Sci. 2020, 7, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.; Xiao, J.; Chen, D.; Turner, S.P.; Li, Z.; Liu, H.; Liu, W.; Liu, J.; Chen, S.; Zhao, X. Feed Restriction Induced Changes in Behavior, Corticosterone, and Microbial Programming in Slow- and Fast-Growing Chicken Breeds. Animals 2021, 11, 141. [Google Scholar] [CrossRef] [PubMed]
- Cedraz, H.; Gromboni, J.G.G.; Garcia, A.A.P.J.; Farias Filho, R.V.; Souza, T.M.; Oliveira, E.R.; Oliveira, E.B.; Nascimento, C.S.D.; Meneghetti, C.; Wenceslau, A.A. Heat stress induces expression of HSP genes in genetically divergent chickens. PLoS ONE 2017, 12, e0186083. [Google Scholar] [CrossRef]
- Radwan, L.M.; Mahrous, M.Y. Genetic selection for growth performance and thermal tolerance under high ambient temperature after two generations using heat shock protein 90 expression as an index. Anim. Prod. Sci. 2019, 59, 628–633. [Google Scholar] [CrossRef]
- Radwan, L.M. Genetic improvement of egg laying traits in Fayoumi chickens bred under conditions of heat stress through selection and gene expression studies. J. Therm. Biol. 2020, 89, 102546–102553. [Google Scholar] [CrossRef]
- Sohn, S.H.; Cho, E.J.; Park, D.B.; Jang, I.S.; Moon, Y.S. Comparison of stress response between Korean Native Chickens and Single Comb White Leghorns subjected to a high stocking density. Korean J. Poult Sci 2014, 41, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Yiallouris, A.; Tsioutis, C.; Agapidaki, E.; Zafeiri, M.; Agouridis, A.P.; Ntourakis, D.; Johnson, E.O. Adrenal Aging and Its Implications on Stress Responsiveness in Humans. Front. Endocrinol. 2019, 10, 54–66. [Google Scholar] [CrossRef]
- Pluvinage, J.V.; Wyss-Coray, T. Systemic factors as mediators of brain homeostasis, ageing and neurodegeneration. Nat. Rev. Neurosci. 2020, 21, 93–102. [Google Scholar] [CrossRef]
- Hu, C.; Yang, J.; Qi, Z.; Wu, H.; Wang, B.; Zou, F.; Mei, H.; Liu, J.; Wang, W.; Liu, Q. Heat shock proteins: Biological functions, pathological roles, and therapeutic opportunities. Med. Comm. 2022, 3, e161. [Google Scholar] [CrossRef]
- Wang, K.; Shang, Y.; Dou, F. Brain Aging: Hsp90 and Neurodegenerative Diseases. Adv. Exp. Med. Biol. 2018, 1086, 93–103. [Google Scholar] [CrossRef]
- Lowman, Z.S.; Edens, F.W.; Ashwell, C.M.; Nolin, S.J. Actigen® Influence on the Gene Expression of Heat Shock Proteins in Ross 708 Broiler Chickens. Int. J. Poult. Sci. 2014, 13, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, D.; Gertler, A.; Cabello, G.; Decuypere, E.; Buyse, J.; Dridi, S. Leptin downregulates heat shock protein-70 (HSP-70) gene expression in chicken liver and hypothalamus. Cell Tissue Res. 2007, 329, 91–101. [Google Scholar] [CrossRef]
- Wang, S.; Edens, F. Involvement of steroids hormones, corticosterone and testosterone, in synthesis of heat shock proteins i broiler chickens. Int. J. Poult. Sci. 2008, 7, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Paroo, Z.; Dipchand, E.S.; Noble, E.G. Estrogen attenuates postexercise HSP70 expression in skeletal muscle. Am. J. Physiol. Cell. Physiol. 2002, 282, C245–C251. [Google Scholar] [CrossRef] [Green Version]
- Paroo, Z.; Tiidus, P.M.; Noble, E.G. Estrogen attenuates HSP 72 expression in acutely exercised male rodents. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 180–184. [Google Scholar] [CrossRef]
- Jones, R.B. Fear and distress. In Animal Welfare; Aplleby, M.C., Hughes, B.O., Eds.; CAB International: Wallingford, UK, 1997; pp. 75–87. [Google Scholar]
- Craig, J.V.; Adams, A.W. Behaviour and Well-being of Hens (Gallus Domesticus) in Alternative Housing Environments. World Poult. Sci. J. 1984, 40, 221–240. [Google Scholar] [CrossRef]
- Jones, R.B. Assessment of fear in adult laying hens: Correlational analysis of methods and measures. Br. Poult. Sci. 1987, 28, 319–326. [Google Scholar] [CrossRef]
- Jones, R.B. The tonic immobility reaction of the domestic fowl: A review. World Poult. Sci. J. 1986, 42, 82–96. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Stress | Types of Samples | Major Classes of HSP | Type of Expression | Effects | Reference |
|---|---|---|---|---|---|
| Thermal stressors | |||||
| Heat stress | |||||
| Leukocytes, testes; bursa of Fabricius | 70 | mRNA | HSP70: ↑ | [24] | |
| Heart; lungs | 90, 70, 29; 27 | Protein | HSP 90, 70, 29; 27: ↑ | [25] | |
| Heart | 90, 70; 60 | mRNA | 2 h of heat stress; HSP 60, 70; 90: ↑ 3 h of heat stress; HSP 60; 90; ↓, HSP 70: - | [26] | |
| Protein | 2 h of heat stress; HSP 60, 70; 90: ↑ 3 h of heat stress; HSP 90: ↓, HSP 60; 70: - | ||||
| Muscle | 90; 70 | mRNA | HSP 70: ↑, HSP 90: - | [27] | |
| Liver | HSP 70: ↑, HSP 90: - | ||||
| Heart | HSP 90: ↑, HSP 70: - | ||||
| Liver | 90 | mRNA | 2 h of heat stress; HSP 90: ↑ 10 h of heat stress; HSP 90: - | [28] | |
| Protein | 2 h of heat stress; HSP 90: ↑ 10 h of heat stress; HSP 90: - | ||||
| Kidney | mRNA | 2 h of heat stress; HSP 90: ↑ 10 h of heat stress; HSP 90: - | |||
| Protein | 2 h of heat stress; HSP 90: ↑ 10 h of heat stress; HSP 90: ↑ | ||||
| Heart | mRNA | 2 h of heat stress; HSP 90: ↑ 10 h of heat stress; HSP 90: ↓ | |||
| Protein | 2 h of heat stress; HSP 90: ↑ 10 h of heat stress; HSP 90: ↑ | ||||
| Testes | 90AA1, 70; 25 | mRNA | HSP 90AA1, 70; 25: ↑ | [29] | |
| Testes | 90AA1, 70, 25, A8, A5, H1; D1 | mRNA | HSP 90AA1, 70, 25, A8, A5, H1; D1: ↑ | [30] | |
| Blood | 90, 70, 60; 27 | mRNA | HSP 90, 70, 60; 27: ↓ | [31] | |
| Liver | 70, 60; 40 | mRNA | HSP 70, 60; 40: ↑ | [32] | |
| 70, 60; 40 | Protein | HSP 40: ↑ HSP 70; 60: - | |||
| Brain | 70 | mRNA | HSP 70: ↑ | [33] | |
| Duodenum | 70, 60; 47 | mRNA | 3 h of heat stress; HSP 70, 60; 47: ↑ | ||
| Protein | 6 h of heat stress; HSP 70; 60: ↑ 3 h of heat stress; HSP 47: ↑ | ||||
| Jejunum | mRNA | 3 h of heat stress; HSP 70, 60; 47: ↑ | [34] | ||
| Protein | 6 h of heat stress; HSP 70; 60: ↑ | ||||
| Ileum | mRNA | 3 h of heat stress; HSP 70, 60; 47: ↑ | |||
| Protein | 3 h of heat stress: HSP 47: ↑ | ||||
| Muscle | 90; 70 | mRNA | HSP 90; 70: - | [27] | |
| Liver | HSP 90: ↑, HSP 70: - | ||||
| Heart | HSP 70: ↑, HSP 90: - | ||||
| Kidney, brain; liver | 70 | Protein | HSP 70: ↓ | [35] | |
| Muscle, heart; brain | 90; 60 | mRNA | D10; HSP 90; 60: ↑ D28; HSP 90; 60: ↑ | [36] | |
| Liver | 90β, 70; 27 | mRNA; protein | HSP 90β, 70; 27: ↓ | [37] | |
| Cold stress | |||||
| Heart | 70, 60, 40, 27 | mRNA | HSP 70, 60, 40; 27: ↑ | [38] | |
| 90 | mRNA; protein | HSP 90: ↓ | |||
| Immune system organs | 90, 70, 60, 40; 27 | Protein | HSP 90, 70, 60, 40; 27: | [39] | |
| Heart | 90, 70; 27 | mRNA | HSP 90, 70; 27: ↓ | [40] | |
| Ileum | 90, 70; 60 | Protein | HSP 90, 70; 60: ↓ | [41] | |
| Hepatic cells | 70 | mRNA | HSP 70: ↓ | [42] | |
| Splenic cells | 70 | mRNA | HSP 70: ↓ | ||
| Heart; muscle | 70 | Protein | HSP 70: ↑ | [43] | |
| Feed restriction | |||||
| Brain | 70 | Protein | HSP 70: ↑ | [44] | |
| Brain | 70 | Protein | HSP 70: ↑ | [45] | |
| Hippocampus | 70 | mRNA | HSP 70: ↑ | [46] | |
| Brain | 70 | Protein | HSP 70: ↑ | [47] | |
| Brain | 70 | Protein | HSP 70: ↑ | [48] | |
| Brain | 70 | Protein | HSP 70: ↑ | [49] | |
| Pre-slaughter operations | |||||
| Crating | Brain | 70 | Protein | HSP 70: ↑ | [50] |
| Transportation | Heart | 110, 90, 70, 60, 47; 40 | mRNA | 2 h of transportation stress; HSP 110, 90, 70, 60, 47; 40: ↑ 8 h of transportation stress; HSP 110, 90, 70, 60, 47; 40: ↓ | [51] |
| 90, 70; 60 | Protein | 2 h of transportation stress; HSP 90, 70; 60: ↑ 8 h of transportation stress; HSP 90, 70; 60: ↓ | |||
| Muscle | 70 | mRNA; protein | HSP 70: ↑ | [52] | |
| Crating and transportation | Liver | 70 | mRNA | HSP 70: ↑ | [53] |
| Social stress | |||||
| Heart | 90, 70, 60; 30 | Protein | HSP 70: ↑ HSP 90, 60; 30: - | [54] | |
| Heart; brain | 70 | Protein | HSP 70: ↑ | [55] | |
| Stocking density | |||||
| Brain | 70 | Protein | HSP 70: ↑ | [56] | |
| Liver | 90; 70 | mRNA | HSP 70: ↑ HSP 90: - | [57] | |
| Heart; liver | 90; 70 | mRNA | HSP 90; 70: ↑ | [58] | |
| Human contact | |||||
| Brain | 70 | Protein | HSP 70: ↑ | [59] | |
| Heavy metal | |||||
| Neutrophils, spleen, testes; heart | 90, 70, 60, 40; 27 | mRNA | HSP 90, 70, 60, 40; 27: ↑ | [60,61,62,63] | |
| Spermatogonia; testis | 90, 70, 60, 40; 27 | mRNA; protein | HSP 90, 70, 60, 40; 27: ↑ | [64] | |
| Testes | 90, 70; 60 | Protein | HSP 90, 70; 60: ↑ | [65] | |
| Cerebellum | 110, 90, 70, 60, 47, 40, 27, 25; 10 | mRNA; protein | HSP 110, 90, 70, 60, 47, 40, 27, 25; 10: ↓ | [66] | |
| Lymphocytes | 90, 70, 60, 40; 27 | mRNA | HSP 90, 70, 60, 40; 27: ↓ | [67] | |
| Liver | 90, 70, 60, 40; 27 | mRNA | HSP 90, 70, 60, 40; 27: ↑ | [68] | |
| Brain, spleen, thymus, jejunum, ileum; duodenum | 90, 70, 60, 40; 27 | mRNA | HSP 90, 70, 60, 40; 27: ↑ | [69,70,71,72] | |
| Mycotoxins | |||||
| Liver | 70 | Protein | HSP 70: ↑ | [73] | |
| Liver | 70 | mRNA | HSP 70: ↑ | [74] | |
| Cecum, ileum; jejunum | 90, 70 | mRNA | HSP 90: (except jejunum) ↑ | [75] | |
| HSP 70: (except ileum) ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balakrishnan, K.N.; Ramiah, S.K.; Zulkifli, I. Heat Shock Protein Response to Stress in Poultry: A Review. Animals 2023, 13, 317. https://doi.org/10.3390/ani13020317
Balakrishnan KN, Ramiah SK, Zulkifli I. Heat Shock Protein Response to Stress in Poultry: A Review. Animals. 2023; 13(2):317. https://doi.org/10.3390/ani13020317
Chicago/Turabian StyleBalakrishnan, Krishnan Nair, Suriya Kumari Ramiah, and Idrus Zulkifli. 2023. "Heat Shock Protein Response to Stress in Poultry: A Review" Animals 13, no. 2: 317. https://doi.org/10.3390/ani13020317
APA StyleBalakrishnan, K. N., Ramiah, S. K., & Zulkifli, I. (2023). Heat Shock Protein Response to Stress in Poultry: A Review. Animals, 13(2), 317. https://doi.org/10.3390/ani13020317