

Placental Development and Physiological Changes in Pregnant Ewes in Silvopastoral and Open Pasture Systems during the Summer
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
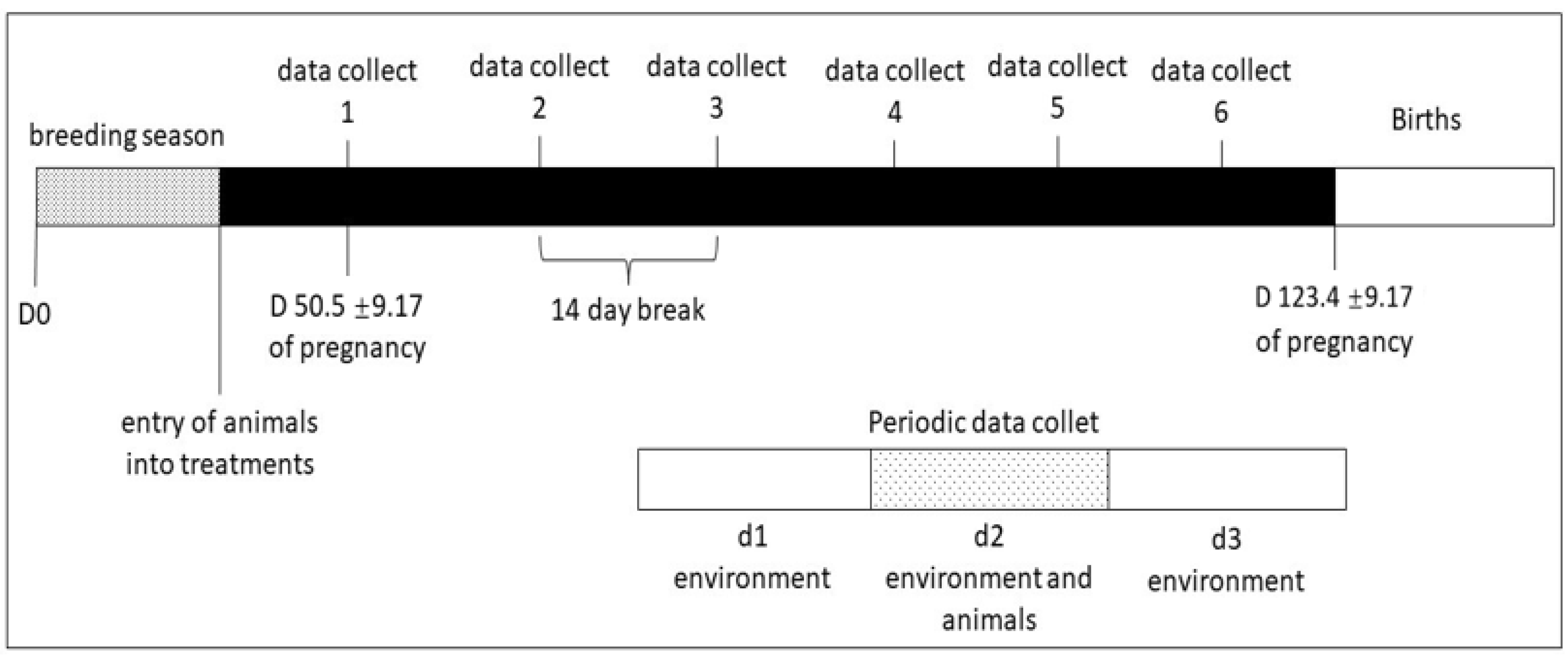
2.1. Study Area, Animals, and Experimental Design
2.2. Microclimatic Variables
2.3. Thermal Comfort Indices
2.4. Evaluation of Physiological Variables
2.5. Peripheral Blood Sampling and Characterization of Leukocytes
2.6. Births and the Collection of Placentas
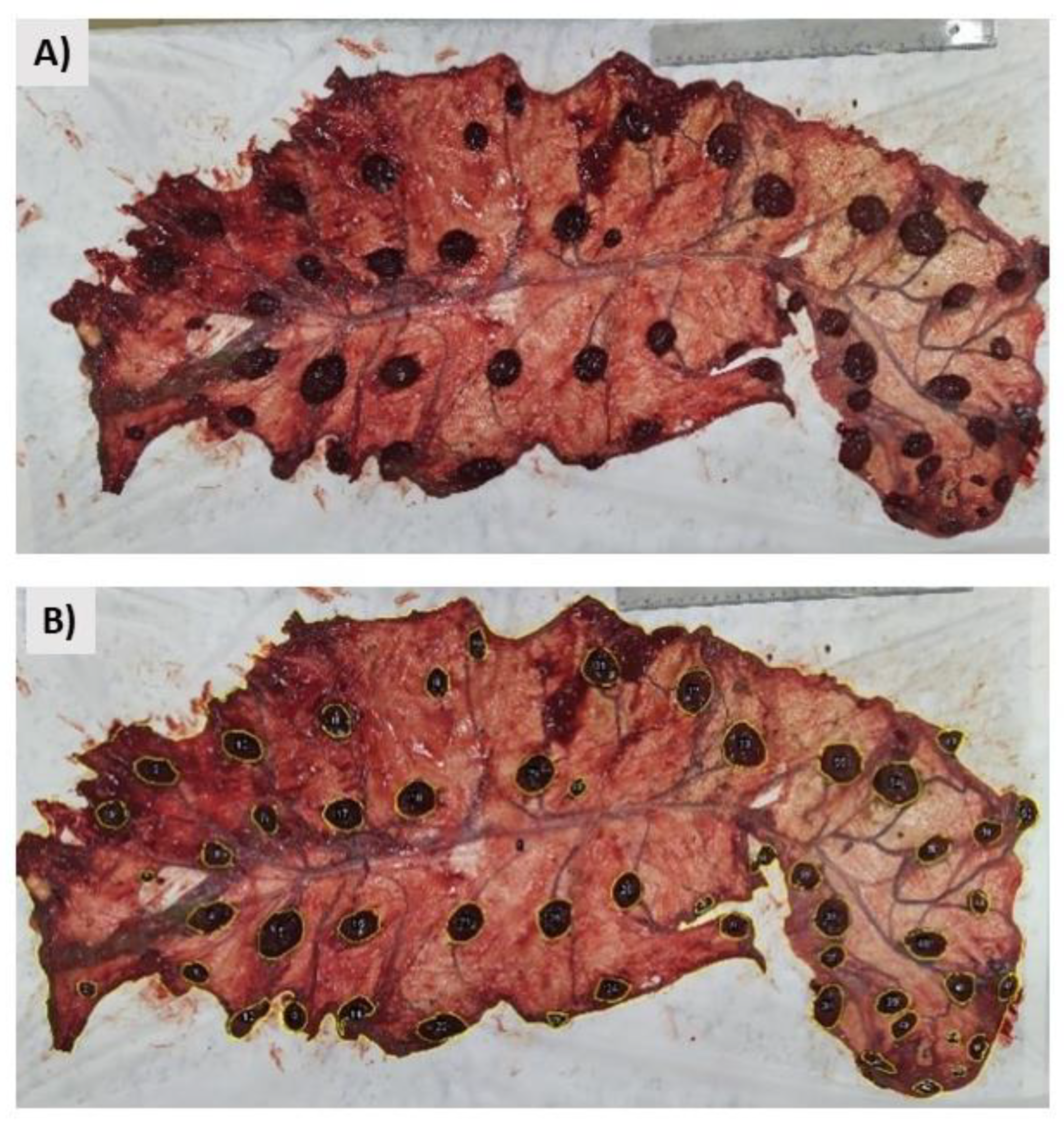
2.7. Placental Biometry
2.8. Statistical Analysis
2.8.1. Environmental and Physiological Variables
2.8.2. Placental Biometry
2.8.3. Leukocyte Characterization
3. Results
3.1. Microclimate Characterization
3.2. Thermoregulatory Variables
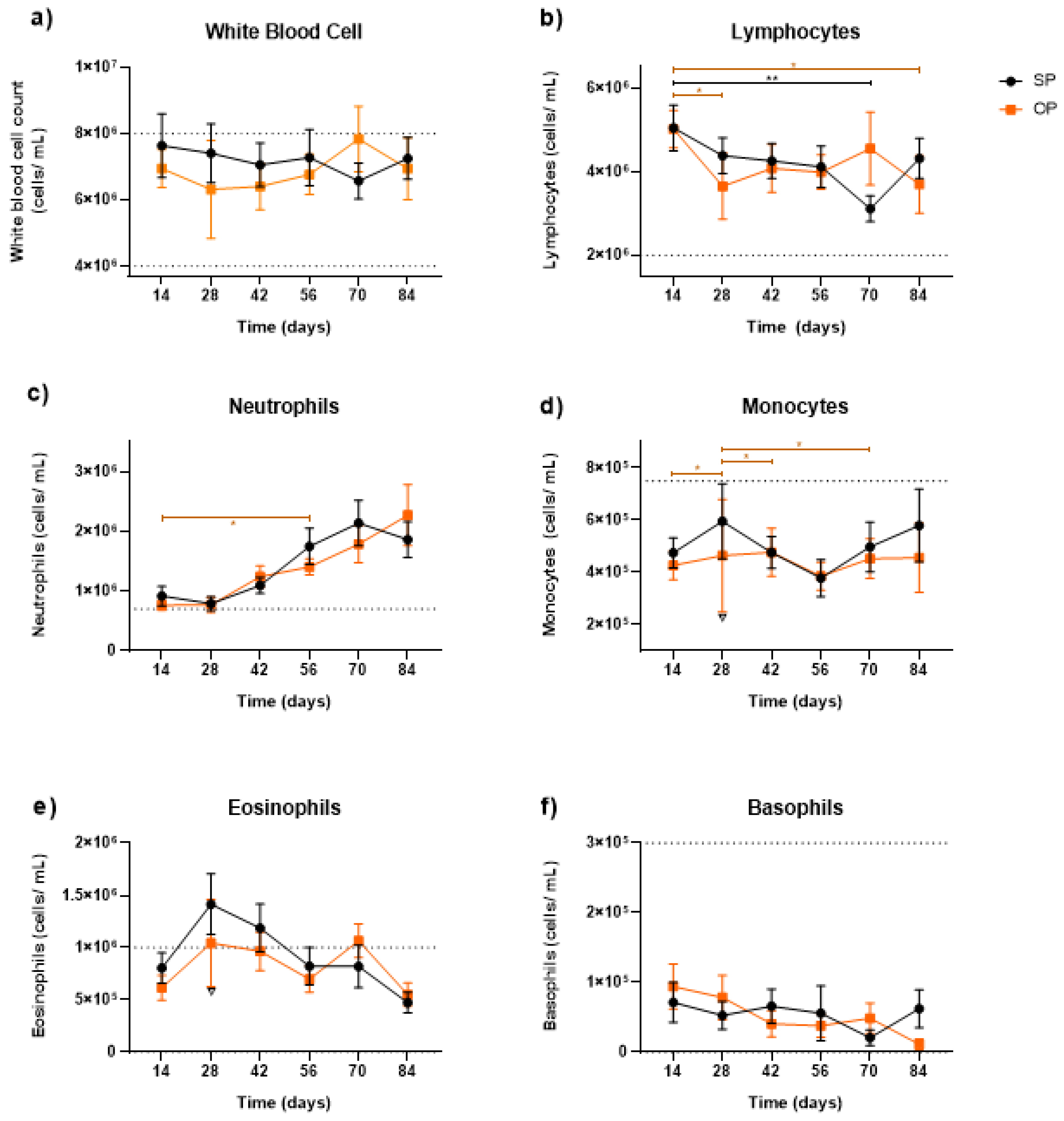
3.3. Leukocyte Characterization
3.4. Reproduction Variables
3.4.1. Gestation and Lambs
3.4.2. Placental Biometry
4. Discussion
4.1. Microclimate Characterization
4.2. Physiological Variables
4.3. Leukocytes
4.4. Gestation and Lambs
4.5. Placental Biometry
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ALVES, F.; de Almeida, R.G. Diretrizes Técnicas Para Produção de Carne Com Baixa Emissão de Carbono Certificada Em Pastagens Tropicais: Carne Baixo Carbono (CBC). 2020. Available online: http://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/1120985 (accessed on 24 November 2022).
- Perez-Garcia, V.; Turco, M.Y. Keep Calm and the Placenta Will Carry On. Dev. Cell 2020, 54, 295–296. [Google Scholar] [CrossRef] [PubMed]
- da Silva, E.M.N.; de Souza, B.B.; de Silva, G.A.; Cezar, M.F.; de Souza, W.H.; Benício, T.M.A.; Freitas, M.M.S. Avaliação Da Adaptabilidade de Caprinos Exóticos e Nativos No Semi-Árido Paraibano. Ciência E Agrotecnologia 2006, 30, 516–521. [Google Scholar] [CrossRef]
- Coelho, L.A.; Sasa, A.; Bicudo, S.D.; Balieiro, J.C.C. Concentrações Plasmáticas de Testosterona, Triiodotironina (T3) e Tiroxina (T4) Em Bodes Submetidos Ao Estresse Calórico. Arq. Bras. Med. Vet. Zootec 2008, 60, 1338–1345. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.L.P.; Dada, J.M.V.; Muniz, P.C.; Nunes-Zotti, M.L.A.; Barros, F.R.O.D.; Vieira, F.M.C. Physiological Responses of Santa Inês x Dorper Ewes and Lambs to Thermal Environment of Silvopasture and Open Pasture Systems. Small Rumin. Res. 2021, 205. [Google Scholar] [CrossRef]
- Rathwa, S.D.; Vasava, A.A.; Pathan, M.M.; Madhira, S.P.; Patel, Y.G.; Pande, A.M. Effect of Season on Physiological, Biochemical, Hormonal, and Oxidative Stress Parameters of Indigenous Sheep. Vet. World 2017, 10, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Dada, J.M.V.; Santos, M.L.P.D.; Muniz, P.C.; Nunes-Zotti, M.L.A.; de Barros, F.R.O.; Vieira, F.M.C. Postpartum Behavioural Response of Santa Inês x Dorper Ewes and Lambs in a Silvopastoral System. Small Rumin. Res. 2021, 203. [Google Scholar] [CrossRef]
- McManus, C.M.; Faria, D.A.; Lucci, C.M.; Louvandini, H.; Pereira, S.A.; Paiva, S.R. Heat Stress Effects on Sheep: Are Hair Sheep More Heat Resistant? Theriogenology 2020, 155, 157–167. [Google Scholar] [CrossRef]
- Limesand, S.W.; Camacho, L.E.; Kelly, A.C.; Antolic, A.T. Impact of Thermal Stress on Placental Function and Fetal Physiology. Anim. Reprod 2018, 15, 886–898. [Google Scholar] [CrossRef]
- Kaufmann, P.; Mayhew, T.M.; Charnock-Jones, D.S. Aspects of Human Fetoplacental Vasculogenesis and Angiogenesis. II. Changes During Normal Pregnancy. Placenta 2004, 25, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Borowicz, P.P.; Vonnahme, K.A.; Johnson, M.L.; Grazul-Bilska, A.T.; Redmer, D.A.; Caton, J.S. Placental Angiogenesis in Sheep Models of Compromised Pregnancy. J. Physiol. 2005, 565, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.E.; Wells, G.D.; Esmay, M.L. Convective Heat Transfer Coefficient for Poultry Excreta. Trans. ASAE 1978, 21, 0534–0536. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: http://www.R-project.org (accessed on 27 December 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Sejian, V.; Bhatta, R.; Gaughan, J.; Malik, P.K.; Naqvi, S.M.K.; Lal, R. Adapting Sheep Production to Climate Change. In Sheep Production Adapting to Climate Change; Springer: Singapore, 2017; pp. 1–29. [Google Scholar]
- Fey, R.; Contro Malavasi, U.; de Matos Malavasi, M. Silvopastoral system: A review regarding the family agriculture. Rev. De Agric. Neotrop. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Alvarado-Canché, A.; Martínez, B.; Castillo, L.E.; Piñeiro-Vázquez, A.; Canul-Soli, J.R. Comportamiento Productivo y Alimenticio de Ovinos En Pastoreo En Sistemas Silvopastoriles Con Leucaena Leucocephala y Cynodon Plectostachyus. Rev. Bio Cienc. 2017, 4, 1–11. [Google Scholar]
- Karvatte, N.; Klosowski, E.S.; de Almeida, R.G.; Mesquita, E.E.; de Oliveira, C.C.; Alves, F.V. Shading Effect on Microclimate and Thermal Comfort Indexes in Integrated Crop-Livestock-Forest Systems in the Brazilian Midwest. Int. J. Biometeorol. 2016, 60, 1933–1941. [Google Scholar] [CrossRef] [Green Version]
- Pezzopane, J.R.M.; Nicodemo, M.L.F.; Bosi, C.; Garcia, A.R.; Lulu, J. Animal Thermal Comfort Indexes in Silvopastoral Systems with Different Tree Arrangements. J. Biol. 2019, 79, 103–111. [Google Scholar] [CrossRef]
- Vieira, F.M.C.; Pilatti, J.A.; Czekoski, Z.M.W.; Fonsêca, V.F.C.; Herbut, P.; Angrecka, S.; de Souza Vismara, E.; de Paulo Macedo, V.; dos Santos, M.C.R.; Paśmionka, I. Effect of the Silvopastoral System on the Thermal Comfort of Lambs in a Subtropical Climate: A Preliminary Study. Agriculture 2021, 11, 790. [Google Scholar] [CrossRef]
- Baêta, F.C.; Souza, C.F. Ambiência Em Edificações Rurais: Conforto Animal, 2nd ed.; UFV: Viçosa, Brazil, 2010. [Google Scholar]
- Eustáquio Filho, A.; Teodoro, S.M.; Chaves, M.A.; dos Santos, P.E.F.; da Silva, M.W.R.; Murta, R.M.; de Carvalho, G.G.P.; de Souza, L.E.B. Zona de Conforto Térmico de Ovinos Da Raça Santa Inês Com Base Nas Respostas Fisiológicas. Rev. Bras. De Zootec. 2011, 40, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Lopes, L.B.; Eckstein, C.; Pina, D.S.; Carnevalli, R.A. The Influence of Trees on the Thermal Environment and Behaviour of Grazing Heifers in Brazilian Midwest. Trop Anim. Health Prod. 2016, 48, 755–761. [Google Scholar] [CrossRef] [Green Version]
- de Sousa, K.T.; Deniz, M.; do Vale, M.M.; Dittrich, J.R.; Hötzel, M.J. Influence of Microclimate on Dairy Cows’ Behavior in Three Pasture Systems during the Winter in South Brazil. J. Biol. 2021, 97, 102873. [Google Scholar] [CrossRef]
- Deniz, M.; Schmitt Filho, A.L.; Farley, J.; de Quadros, S.F.; Hötzel, M.J. High Biodiversity Silvopastoral System as an Alternative to Improve the Thermal Environment in the Dairy Farms. Int. J. Biometeorol. 2019, 63, 83–92. [Google Scholar] [CrossRef] [PubMed]
- da Silva, W.E.; Leite, J.H.G.M.; de Sousa, J.E.R.; Costa, W.P.; da Silva, W.S.T.; Guilhermino, M.M.; Asensio, L.A.B.; Façanha, D.A.E. Daily Rhythmicity of the Thermoregulatory Responses of Locally Adapted Brazilian Sheep in a Semiarid Environment. Int. J. Biometeorol. 2017, 61, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Pent, G.J.; Greiner, S.P.; Munsell, J.F.; Tracy, B.F.; Fike, J.H. Lamb Performance in Hardwood Silvopastures, II: Animal Behavior in Summer1. Transl. Anim. Sci. 2020, 4, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Hyder, I.; Pasumarti, M.; Reddy, P.R.; Prasad, C.S.; Kumar, K.A.; Sejian, V. Thermotolerance in Domestic Ruminants: A HSP70 Perspective. In Heat Shock Proteins in Veterinary Medicine and Sciences; Springer: Berlin/Heidelberg, Germany, 2017; pp. 3–35. [Google Scholar]
- Ghassemi Nejad, J.; Sung, K.-I. Behavioral and Physiological Changes during Heat Stress in Corriedale Ewes Exposed to Water Deprivation. J. Anim. Sci. Technol. 2017, 59, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pehlivan, E.; Kaliber, M.; Konca, Y.; Dellal, G. Effect of Shearing on Some Physiological and Hormonal Parameters in Akkaraman Sheep. Asian-Australas J. Anim. Sci. 2020, 33, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Dias e Silva, T.P.; da Costa Torreão, J.N.; Torreão Marques, C.A.; de Araújo, M.J.; Bezerra, L.R.; kumar Dhanasekaran, D.; Sejian, V. Effect of Multiple Stress Factors (Thermal, Nutritional and Pregnancy Type) on Adaptive Capability of Native Ewes under Semi-Arid Environment. J. Biol. 2016, 59, 39–46. [Google Scholar] [CrossRef]
- Karthik, D.; Suresh, J.; Reddy, Y.R.; Sharma, G.R.K.; Ramana, J.V.; Gangaraju, G.; Reddy, P.P.R.; Reddy, Y.P.K.; Yasaswini, D.; Adegbeye, M.J.; et al. Adaptive Profiles of Nellore Sheep with Reference to Farming System and Season: Physiological, Hemato-Biochemical, Hormonal, Oxidative-Enzymatic and Reproductive Standpoint. Heliyon 2021, 7, e07117. [Google Scholar] [CrossRef]
- Seixas, L.; de Melo, C.B.; Menezes, A.M.; Ramos, A.F.; Paludo, G.R.; Peripolli, V.; Tanure, C.B.; Costa Junior, J.B.G.; McManus, C. Study on Environmental Indices and Heat Tolerance Tests in Hair Sheep. Trop. Anim. Health Prod. 2017, 49, 975–982. [Google Scholar] [CrossRef]
- Wojtas, K.; Cwynar, P.; Kołacz, R. Effect of Thermal Stress on Physiological and Blood Parameters in Merino Sheep. Bull. Vet. Inst. Pulawy 2014, 58, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Kamel, N.N.; Ahmed, A.M.H.; Mehaisen, G.M.K.; Mashaly, M.M.; Abass, A.O. Depression of Leukocyte Protein Synthesis, Immune Function and Growth Performance Induced by High Environmental Temperature in Broiler Chickens. Int. J. Biometeorol. 2017, 61, 1637–1645. [Google Scholar] [CrossRef]
- Bagath, M.; Krishnan, G.; Devaraj, C.; Rashamol, V.P.; Pragna, P.; Lees, A.M.; Sejian, V. The Impact of Heat Stress on the Immune System in Dairy Cattle: A Review. Res. Vet. Sci. 2019, 126, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Gharibi, V.; Khanjani, N.; Heidari, H.; Ebrahimi, M.H.; Hosseinabadi, M.B. The Effect of Heat Stress on Hematological Parameters and Oxidative Stress among Bakery Workers. Toxicol. Ind. Health 2020, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.; Day, R.; Slocombe, R.; Benkendorff, K.; Handlinger, J.; Goulias, J. Effects of Severe Heat Stress on Immune Function, Biochemistry and Histopathology in Farmed Australian Abalone (Hybrid Haliotis Laevigata × Haliotis Rubra). Aquaculture 2014, 432, 26–37. [Google Scholar] [CrossRef]
- Caroprese, M.; Ciliberti, M.G.; Annicchiarico, G.; Albenzio, M.; Muscio, A.; Sevi, A. Hypothalamic-Pituitary-Adrenal Axis Activation and Immune Regulation in Heat-Stressed Sheep after Supplementation with Polyunsaturated Fatty Acids. J. Dairy Sci. 2014, 97, 4247–4258. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.M.; Tait, R.G.; Galles, B.M.; Duffy, E.M.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Heat Stress-Induced Deficits in Growth, Metabolic Efficiency, and Cardiovascular Function Coincided with Chronic Systemic Inflammation and Hypercatecholaminemia in Ractopamine-Supplemented Feedlot Lambs. J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef]
- Liu, H.; Li, K.; Mingbin, L.; Zhao, J.; Xiong, B. Effects of Chestnut Tannins on the Meat Quality, Welfare, and Antioxidant Status of Heat-Stressed Lambs. Meat Sci. 2016, 116, 236–242. [Google Scholar] [CrossRef]
- Seiler, A.; Fagundes, C.P.; Christian, L.M. The Impact of Everyday Stressors on the Immune System and Health. In Stress Challenges and Immunity in Space; Springer International Publishing: Cham, Germany, 2020; pp. 71–92. [Google Scholar]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological Traits as Affected by Heat Stress in Sheep—A Review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Romo-Barron, C.B.; Diaz, D.; Portillo-Loera, J.J.; Romo-Rubio, J.A.; Jimenez-Trejo, F.; Montero-Pardo, A. Impact of Heat Stress on the Reproductive Performance and Physiology of Ewes: A Systematic Review and Meta-Analyses. Int. J. Biometeorol. 2019. [Google Scholar] [CrossRef]
- van Wettere, W.H.E.J.; Kind, K.L.; Gatford, K.L.; Swinbourne, A.M.; Leu, S.T.; Hayman, P.T.; Kelly, J.M.; Weaver, A.C.; Kleemann, D.O.; Walker, S.K. Review of the Impact of Heat Stress on Reproductive Performance of Sheep. J. Anim. Sci. Biotechnol. 2021, 12, 26. [Google Scholar] [CrossRef]
- Silva, P.S.; Hooper, H.B.; Manica, E.; Merighe, G.K.F.; Oliveira, S.A.; Traldi, A.S.; Negrão, J.A. Heat Stress Affects the Expression of Key Genes in the Placenta, Placental Characteristics, and Efficiency of Saanen Goats and the Survival and Growth of Their Kids. J. Dairy Sci. 2021, 104, 4970–4979. [Google Scholar] [CrossRef]
- Casamassima, D.; Sevi, A.; Palazzo, M.; Ramacciato, R.; Colella, G.E.; Bellitti, A. Effects of Two Different Housing Systems on Behavior, Physiology and Milk Yield of Comisana Ewes. Small Rumin. Res. 2001, 41, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Dahl, G.E. Invited Review: Heat Stress Effects during Late Gestation on Dry Cows and Their Calves. J. Dairy Sci. 2013, 96, 4079–4093. [Google Scholar] [CrossRef] [PubMed]
- Coloma-García, W.; Mehaba, N.; Llonch, P.; Caja, G.; Such, X.; Salama, A.A.K. Prenatal Heat Stress Effects on Gestation and Postnatal Behavior in Kid Goats. PLoS ONE 2020, 15, e0220221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, C.M.; Calvert, S.K.; Farish, M.; Donbavand, J.; Pickup, H.E. Breed, Litter and Parity Effects on Placental Weight and Placentome Number, and Consequences for the Neonatal Behaviour of the Lamb. Theriogenology 2005, 63, 1092–1110. [Google Scholar] [CrossRef] [PubMed]
- Özyürek, S.; Türkyilmaz, D. Determination of Relationships between Placental Characteristics and Birth Weight in Morkaraman Sheep. Arch. Anim. Breed 2020, 63, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Lipa, M.; Wielgos, M.; Cohen, D.; Middeldorp, J.M.; Oepkes, D.; Lopriore, E. Comparison between Monochorionic and Dichorionic Placentas with Special Attention to Vascular Anastomoses and Placental Share. Twin Res. Hum. Genet. 2016, 19, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Brzozowska, A.; Wojtasiak, N.; Błaszczyk, B.; Stankiewicz, T.; Udała, J.; Wieczorek-Dąbrowska, M. The effects of non-genetic factors on the morphometric parameters of sheep placenta and the birth weight of lambs. Large Anim. Rev. 2020, 26, 119–126. [Google Scholar]
- Wilson, M.E.; Ford, S.P. Comparative Aspects of Placental Efficiency. Reprod Suppl. 2001, 58, 223–232. [Google Scholar] [CrossRef]
- Mellor, D.J. Nutritional and Placental Determinants of Foetal Growth Rate in Sheep and Consequences for the Newborn Lamb. Br. Vet. J. 1983, 139, 307–324. [Google Scholar] [CrossRef]
- Alexander, G. Studies on the placenta of the sheep (Ovis aries L.). Reproduction 1964, 7, 307–322. [Google Scholar] [CrossRef]
- Ocak, S.; Ogun, S.; Onder, H. Relationship between Placental Traits and Maternal Intrinsic Factors in Sheep. Anim. Reprod. Sci. 2013, 139, 31–37. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | Open Pasture (OP) | Silvopastoral System (SP) | p Value | |
|---|---|---|---|---|
| Air temperature (AT, °C) | Mean ± SD | 26.9 ± 0.41 A | 26.0 ± 0.38 B | 0.0288 |
| Minimum | 17.0 | 17.4 | ||
| Median | 26.2 | 26.2 | ||
| Maximum | 34.3 | 33.3 | ||
| Relative humidity (RH, %) | Mean ± SD | 67.2 ± 3.42 A | 68.4 ± 3.37 A | |
| Minimum | 28.1 | 28.3 | 0.4011 | |
| Median | 69.5 | 69.5 | ||
| Maximum | 93.7 | 93.9 | ||
| Wind speed (WS, m s−1) | Mean ± SD | 1.25 ± 0.20 A | 1.02 ± 0.13 A | 0.0939 |
| Minimum | 0 | 0 | ||
| Median | 0.86 | 0.86 | ||
| Maximum | 4.04 | 5.69 | ||
| Grass temperature (GT, °C) | Mean ± SD | 25.6 ± 0.44 A | 23.4 ± 0.37 B | 0.0043 |
| Minimum | 11.0 | 9.6 | ||
| Median | 23.8 | 23.8 | ||
| Maximum | 37.6 | 31.4 | ||
| Radiant heat load (RHL, W m−2) | Mean ± SD | 610 ± 12.3 A | 576 ± 10.9 B | 0.288 |
| Minimum | ||||
| Median | ||||
| Maximum |
| Variables | Open Pasture (OP) | Silvopastoral System (SP) | p Value | |
|---|---|---|---|---|
| Heart rate (HR, beats min.−1) | Mean ± SD | 116 ± 4.85 A | 100 ± 4.84 B | <0.001 |
| Minimum | 64 | 48 | ||
| Median | 117 | 97 | ||
| Maximum | 178 | 166 | ||
| Respiratory rate (RR, mov. min.−1) | Mean ± SD | 104.1 ± 10.0 A | 76.4 ± 10.1 B | |
| Minimum | 24 | 36 | <0.001 | |
| Median | 102 | 68 | ||
| Maximum | 198 | 182 | ||
| Rectal temperature (RT, °C) | Mean ± SD | 39.35 ± 0.06 A | 39.33 ± 0.06 A | 0.06742 |
| Minimum | 38.4 | 38.4 | ||
| Median | 39.4 | 39.3 | ||
| Maximum | 40.0 | 40.4 | ||
| Mean surface temperature (MST, °C) | Mean ± SD | 32.8 ± 0.76 A | 30.0 ± 0.76 B | <0.001 |
| Minimum | 26.0 | 25.3 | ||
| Median | 33.4 | 29.7 | ||
| Maximum | 44.4 | 38.5 |
| Variables | Open Pasture (OP) | Silvopastoral System (SP) | p Value |
|---|---|---|---|
| Duration of pregnancy (days) | 138.6 ± 12.79 | 137.6 ± 16.39 | 0.7954 |
| Birth weight (lambs) (kg) | 3.937 ± 0.4506 | 3.921 ± 0.5943 | 0.9441 |
| Ten days weight (lambs) (kg) | 5.788 ± 0.8883 | 6.124 ± 1.1257 | 0.5218 |
| Weight gain (lambs) (kg) | 1.774 ± 1.010 | 2.186 ± 0.7481 | 0.3079 |
| Variables | Open Pasture (OP) | Silvopastoral System (SP) | p Value |
|---|---|---|---|
| Membrane area (simple pregnancy) (cm2) | 2296 ± 473.6 | 2298 ± 490.4 | 0.9578 |
| Cotyledon area (simple pregnancy) (cm2) | 234.7 ± 69.59 | 226.4 ± 68.62 | 0.8456 |
| Number of cotyledons | 60.22 ± 22.06 | 70.75 ± 12.61 | 0.2541 |
| Placental mass (kg) | 0.3423 ± 0.08057 | 0.3370 ± 0.1084 | 0.9092 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dada, J.M.V.; Santos, M.L.P.d.; Dani, A.P.S.; Dammann, C.P.J.; Pinto, L.; Vieira, F.M.C.; Barros, F.R.O.d. Placental Development and Physiological Changes in Pregnant Ewes in Silvopastoral and Open Pasture Systems during the Summer. Animals 2023, 13, 478. https://doi.org/10.3390/ani13030478
Dada JMV, Santos MLPd, Dani APS, Dammann CPJ, Pinto L, Vieira FMC, Barros FROd. Placental Development and Physiological Changes in Pregnant Ewes in Silvopastoral and Open Pasture Systems during the Summer. Animals. 2023; 13(3):478. https://doi.org/10.3390/ani13030478
Chicago/Turabian StyleDada, Julia Morgana Vieira, Matheus Luquirini Penteado dos Santos, Ana Paula Schneiders Dani, Cecília Paulina Johann Dammann, Letícia Pinto, Frederico Márcio Corrêa Vieira, and Flávia Regina Oliveira de Barros. 2023. "Placental Development and Physiological Changes in Pregnant Ewes in Silvopastoral and Open Pasture Systems during the Summer" Animals 13, no. 3: 478. https://doi.org/10.3390/ani13030478
APA StyleDada, J. M. V., Santos, M. L. P. d., Dani, A. P. S., Dammann, C. P. J., Pinto, L., Vieira, F. M. C., & Barros, F. R. O. d. (2023). Placental Development and Physiological Changes in Pregnant Ewes in Silvopastoral and Open Pasture Systems during the Summer. Animals, 13(3), 478. https://doi.org/10.3390/ani13030478