
Detection and Molecular Characterization of Giardia and Cryptosporidium spp. Circulating in Wild Small Mammals from Portugal
 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fecal Sample Collection
2.2. Nucleic Acid Extraction
2.3. General Procedure for PCR Amplifications
2.4. Giardia spp. DNA Screening
2.5. Cryptosporidium spp. DNA Screening
2.6. Sequencing and Sequence Analysis
2.7. Mapping and Statistical Analysis
3. Results
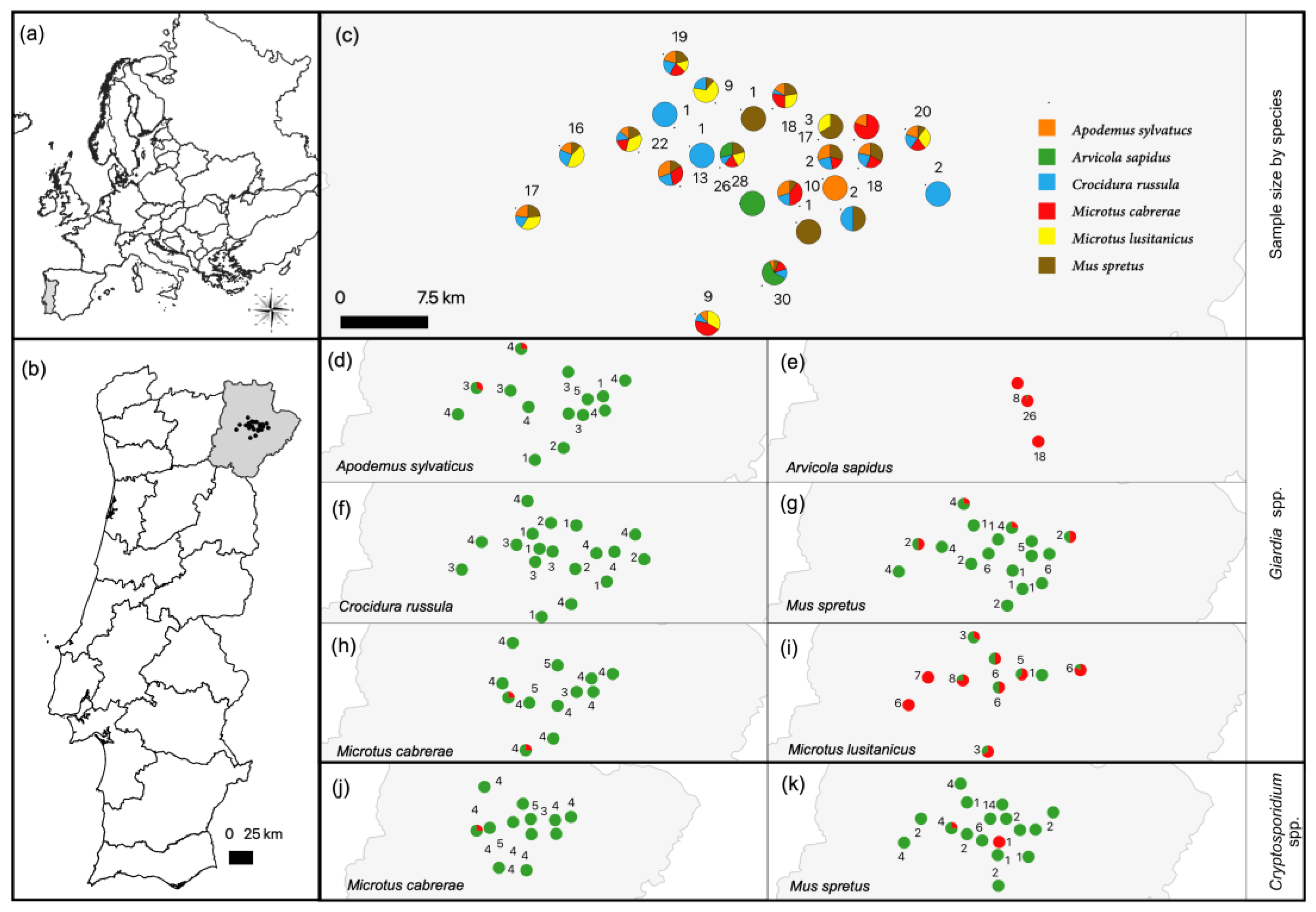
3.1. Occurrence of Giardia spp. and Cryptosporidium spp.
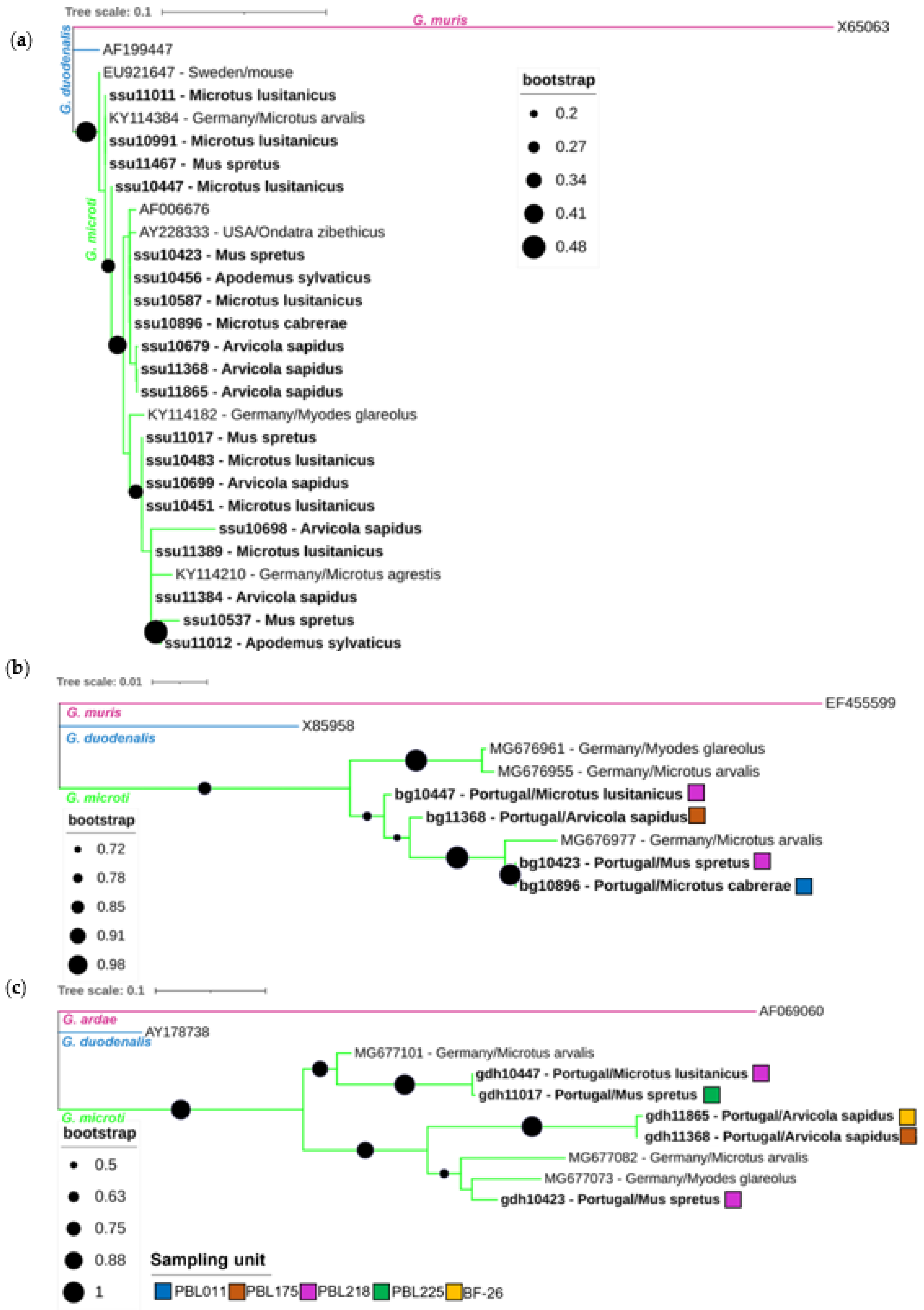
3.2. Characterization of Giardia spp.
3.3. Characterization of Cryptosporidium spp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendoza, H.; Rubio, A.V.; García-Peña, G.E.; Suzán, G.; Simonetti, J.A. Does land-use change increase the abundance of zoonotic reservoirs? Rodents say yes. Eur. J. Wildl. Res. 2020, 66, 6. [Google Scholar] [CrossRef]
- Kivistö, R.; Kämäräinen, S.; Huitu, O.; Niemimaa, J.; Henttonen, H. Zoonotic Cryptosporidium spp. in Wild Rodents and Shrews. Microorganisms 2021, 9, 2242. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B.G.; Singleton, G.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Galán-Puchades, M.; Trelis, M.; Sáez-Durán, S.; Cifre, S.; Gosálvez, C.; Sanxis-Furió, J.; Pascual, J.; Bueno-Marí, R.; Franco, S.; Peracho, V.; et al. One Health Approach to Zoonotic Parasites: Molecular Detection of Intestinal Protozoans in an Urban Population of Norway Rats, Rattus norvegicus, in Barcelona, Spain. Pathogens 2021, 10, 311. [Google Scholar] [CrossRef] [PubMed]
- Riebold, D.; Russow, K.; Schlegel, M.; Wollny, T.; Thiel, J.; Freise, J.; Hüppop, O.; Eccard, J.A.; Plenge-Bönig, A.; Loebermann, M.; et al. Occurrence of Gastrointestinal Parasites in Small Mammals from Germany. Vector-Borne Zoonotic Dis. 2020, 20, 125–133. [Google Scholar] [CrossRef]
- Hunter, P.R.; Thompson, R.A. The zoonotic transmission of Giardia and Cryptosporidium. Int. J. Parasitol. 2005, 35, 1181–1190. [Google Scholar] [CrossRef]
- Krawczyk, A.I.; van Leeuwen, A.D.; Jacobs-Reitsma, W.; Wijnands, L.M.; Bouw, E.; Jahfari, S.; van Hoek, A.H.A.M.; van der Giessen, J.W.B.; Roelfsema, J.H.; Kroes, M.; et al. Presence of zoonotic agents in engorged ticks and hedgehog faeces from Erinaceus europaeus in (sub) urban areas. Parasites Vectors 2015, 8, 210. [Google Scholar] [CrossRef]
- Torres, J.; Gracenea, M.; Gómez, M.; Arrizabalaga, A.; González-Moreno, O. The occurrence of Cryptosporidium parvum and C. muris in wild rodents and insectivores in Spain. Vet.-Parasitol. 2000, 92, 253–260. [Google Scholar] [CrossRef]
- Sprong, H.; Cacciò, S.M.; van der Giessen, J.W.B.; on behalf of the ZOOPNET network and partners. Identification of Zoonotic Genotypes of Giardia duodenalis. PLoS Negl. Trop. Dis. 2009, 3, e558. [Google Scholar] [CrossRef]
- Heyworth, M.F. Giardia duodenalis genetic assemblages and hosts. Parasite 2016, 23, 13. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Ryan, U.; Xiao, L.; Feng, Y. Zoonotic giardiasis: An update. Parasitol. Res. 2021, 120, 4199–4218. [Google Scholar] [CrossRef] [PubMed]
- Cacciò, S.M.; Lalle, M.; Svärd, S.G. Host specificity in the Giardia duodenalis species complex. Infect. Genet. Evol. 2018, 66, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.A.; Monis, P. Giardia—From Genome to Proteome. Adv. Parasitol. 2012, 78, 57–95. [Google Scholar] [CrossRef]
- Monis, P.T.; Andrews, R.H.; Mayrhofer, G.; Ey, P.L. Genetic diversity within the morphological species Giardia intestinalis and its relationship to host origin. Infect. Genet. Evol. 2003, 3, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Kunstýř, I.; Schoeneberg, U.; Friedhoff, K.T. Host specificity ofGiardia muris isolates from mouse and golden hamster. Parasitol. Res. 1992, 78, 621–622. [Google Scholar] [CrossRef]
- Van Keulen, H.; Feely, D.E.; Macechko, P.T.; Jarroll, E.L.; Erlandsen, S.L. The sequence of Giardia small subunit rRNA shows that voles and muskrats are parasitized by a unique species Giardia microti. J. Parasitol. 1998, 84, 294. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Spierling, N.G.; Schmidt, S.; Rosenfeld, U.M.; Reil, D.; Imholt, C.; Jacob, J.; Ulrich, R.G.; Aebischer, T.; Klotz, C. Occurrence and distribution of Giardia species in wild rodents in Germany. Parasites Vectors 2018, 11, 213. [Google Scholar] [CrossRef]
- Lecová, L.; Hammerbauerová, I.; Tůmová, P.; Nohýnková, E. Companion animals as a potential source of Giardia intestinalis infection in humans in the Czech Republic—A pilot study. Vet. Parasitol. Reg. Stud. Rep. 2020, 21, 100431. [Google Scholar] [CrossRef]
- Perec-Matysiak, A.; Buńkowska-Gawlik, K.; Zaleśny, G.; Hildebrand, J. Small rodents as reservoirs of Cryptosporidium spp. and Giardia spp. in south-western Poland. Ann. Agric. Environ. Med. 2015, 22, 1–5. [Google Scholar] [CrossRef]
- Tsui, C.K.-M.; Miller, R.; Uyaguari-Diaz, M.; Tang, P.; Chauve, C.; Hsiao, W.; Isaac-Renton, J.; Prystajecky, N. Beaver Fever: Whole-Genome Characterization of Waterborne Outbreak and Sporadic Isolates to Study the Zoonotic Transmission of Giardiasis. Msphere 2018, 3, e00090-18. [Google Scholar] [CrossRef] [Green Version]
- Fayer, R.; Santín, M.; Trout, J.M.; DeStefano, S.; Koenen, K.; Kaur, T. Prevalence of Microsporidia, Cryptosporidium spp., and Giardia spp. in Beavers (CASTOR CANADENSIS) in Massachusetts. J. Zoo Wildl. Med. 2006, 37, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Erlandsen, S.L.; Sherlock, L.A.; Bemrick, W.J.; Ghobrial, H.; Jakubowski, W. Prevalence of Giardia spp. in beaver and muskrat populations in northeastern states and Minnesota: Detection of intestinal trophozoites at necropsy provides greater sensitivity than detection of cysts in fecal samples. Appl. Environ. Microbiol. 1990, 56, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Gherman, C.M.; Kalmár, Z.; Györke, A.; Mircean, V. Occurrence of Giardia duodenalis assemblages in farmed long-tailed chinchillas Chinchilla lanigera (Rodentia) from Romania. Parasites Vectors 2018, 11, 86. [Google Scholar] [CrossRef]
- Levecke, B.; Meulemans, L.; Dalemans, T.; Casaert, S.; Claerebout, E.; Geurden, T. Mixed Giardia duodenalis assemblage A, B, C and E infections in pet chinchillas (Chinchilla lanigera) in Flanders (Belgium). Vet.-Parasitol. 2011, 177, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Klotz, C.; Radam, E.; Rausch, S.; Gosten-Heinrich, P.; Aebischer, T. Real-Time PCR for Molecular Detection of Zoonotic and Non-Zoonotic Giardia spp. in Wild Rodents. Microorganisms 2021, 9, 1610. [Google Scholar] [CrossRef]
- Asghari, A.; Motazedian, M.H.; Asgari, Q.; Shamsi, L.; Sarkari, B.; Shahabi, S.; Mohammadi-Ghalehbin, B. Occurrence, genetic characterization, and zoonotic importance of Giardia duodenalis in various species of rodents (Mus musculus, Rattus norvegicus, and Rattus rattus). Comp. Immunol. Microbiol. Infect. Dis. 2022, 85, 101812. [Google Scholar] [CrossRef] [PubMed]
- De Liberato, C.; Di Filippo, M.M.; Sagrafoli, D.; Ferraro, D.; Procesi, I.G.; Berrilli, F. Giardia microti in pet Microtus guentheri: Evidence of a parasite never detected in Italy. Parasitol. Int. 2021, 80, 102207. [Google Scholar] [CrossRef]
- Fernández-Álvarez, Á.; Martín-Alonso, A.; Abreu-Acosta, N.; Feliu, C.; Hugot, J.-P.; Valladares, B.; Foronda, P. Identification of a novel assemblage G subgenotype and a zoonotic assemblage B in rodent isolates of Giardia duodenalis in the Canary Islands, Spain. Parasitology 2014, 141, 206–215. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, R.; Zhao, W.; Qi, M.; Zhao, J.; Zhang, L.; Li, J.; Liu, A. Genotyping and subtyping of Giardia and Cryptosporidium isolates from commensal rodents in China. Parasitology 2015, 142, 800–806. [Google Scholar] [CrossRef]
- Ryan, U.M.; Feng, Y.; Fayer, R.; Xiao, L. Taxonomy and molecular epidemiology of Cryptosporidium and Giardia—A 50 year perspective (1971–2021). Int. J. Parasitol. 2021, 51, 1099–1119. [Google Scholar] [CrossRef]
- Khan, A.; Shaik, J.S.; Grigg, M.E. Genomics and molecular epidemiology of Cryptosporidium species. Acta Trop. 2018, 184, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Gerace, E.; Lo Presti, V.D.M.; Biondo, C. Cryptosporidium infection: Epidemiology, pathogenesis, and differential diagnosis. Eur. J. Microbiol. Immunol. 2019, 9, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.J.; Vinayak, S. Cryptosporidium: Host-Parasite Interactions and Pathogenesis. Curr. Clin. Microbiol. Rep. 2021, 8, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Zahedi, A.; Feng, Y.; Xiao, L. An Update on Zoonotic Cryptosporidium Species and Genotypes in Humans. Animals 2021, 11, 3307. [Google Scholar] [CrossRef]
- Feng, Y.; Ryan, U.M.; Xiao, L. Genetic Diversity and Population Structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef]
- Xiao, L.; Feng, Y. Molecular epidemiologic tools for waterborne pathogens Cryptosporidium spp. and Giardia duodenalis. Food Waterborne Parasitol. 2017, 8–9, 14–32. [Google Scholar] [CrossRef]
- Danišová, O.; Valenčáková, A.; Stanko, M.; Luptáková, L.; Hatalová, E.; Čanády, A. Rodents as a reservoir of infection caused by multiple zoonotic species/genotypes of C. parvum, C. hominis, C. suis, C. scrofarum, and the first evidence of C. muskrat genotypes I and II of rodents in Europe. Acta Trop. 2017, 172, 29–35. [Google Scholar] [CrossRef]
- Song, J.; Kim, C.-Y.; Chang, S.-N.; Abdelkader, T.S.; Han, J.; Kim, T.-H.; Oh, H.; Lee, J.M.; Kim, D.-S.; Kim, J.-T.; et al. Detection and Molecular Characterization of Cryptosporidium spp. from Wild Rodents and Insectivores in South Korea. Korean J. Parasitol. 2015, 53, 737–743. [Google Scholar] [CrossRef]
- Vioque, F.; Dashti, A.; Santín, M.; Ruiz-Fons, F.; Köster, P.C.; Hernández-Castro, C.; García, J.T.; Bailo, B.; Ortega, S.; Olea, P.P.; et al. Wild micromammal host spectrum of zoonotic eukaryotic parasites in Spain. Occurrence and genetic characterisation. Transbound. Emerg. Dis. 2022, 69, e2926–e2942. [Google Scholar] [CrossRef]
- Zhang, K.; Fu, Y.; Li, J.; Zhang, L. Public health and ecological significance of rodents in Cryptosporidium infections. One Health 2022, 14, 100364. [Google Scholar] [CrossRef] [PubMed]
- Taghipour, A.; Olfatifar, M.; Foroutan, M.; Bahadory, S.; Malih, N.; Norouzi, M. Global prevalence of Cryptosporidium infection in rodents: A systematic review and meta-analysis. Prev. Vet.-Med. 2020, 182, 105119. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Moreira, M.J.; Soares, S.; Delgado, M.D.L.; Figueiredo, J.; Silva, E.; Castro, A.; Da Cosa, J.M.C. Presence of Cryptosporidium spp. and Giardia duodenalis in Drinking Water Samples in the North of Portugal. Korean J. Parasitol. 2010, 48, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Lobo, M.; Xiao, L.; Antunes, F.; Matos, O. Occurrence of Cryptosporidium and Giardia genotypes and subtypes in raw and treated water in Portugal. Lett. Appl. Microbiol. 2009, 48, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Júlio, C.; Sá, C.; Ferreira, I.; Martins, S.; Oleastro, M.; Ângelo, H.; Guerreiro, J.; Tenreiro, R. Waterborne transmission of Giardia and Cryptosporidium at river beaches in Southern Europe (Portugal). J. Water Health 2012, 10, 484–496. [Google Scholar] [CrossRef]
- Almeida, A.A.; Delgado, M.L.; Soares, S.C.; Castro, A.O.; Moreira, M.J.; Mendonça, C.M.; Canada, N.B.; Da Costa, J.M.C.; Coelho, H.G. Genetic Characterization of Cryptosporidium Isolates from Humans in Northern Portugal. J. Eukaryot. Microbiol. 2006, 53, S26–S27. [Google Scholar] [CrossRef]
- Júlio, C.; Vilares, A.; Oleastro, M.; Ferreira, I.; Gomes, S.; Monteiro, L.; Nunes, B.; Tenreiro, R.; Ângelo, H. Prevalence and risk factors for Giardia duodenalis infection among children: A case study in Portugal. Parasites Vectors 2012, 5, 22–28. [Google Scholar] [CrossRef]
- Barão, I.; Queirós, J.; Vale-Gonçalves, H.; Paupério, J.; Pita, R. Landscape Characteristics Affecting Small Mammal Occurrence in Heterogeneous Olive Grove Agro-Ecosystems. Conservation 2022, 2, 51–67. [Google Scholar] [CrossRef]
- Instituto da Conservação da Natureza e das Florestas (ICNF). Parque Natural de Montesinho. Available online: http://www2.icnf.pt/portal/ap/p-nat/pnm (accessed on 1 September 2021).
- Sutherland, G.D.; Harestad, A.S.; Price, K.; Lertzman, K.P. Scaling of Natal Dispersal Distances in Terrestrial Birds and Mammals. Conserv. Ecol. 2000, 4, 16. [Google Scholar] [CrossRef]
- Verweij, J.J.; Schinkel, J.; Laeijendecker, D.; van Rooyen, M.A.; van Lieshout, L.; Polderman, A.M. Real-time PCR for the detection of Giardia lamblia. Mol. Cell Probes 2003, 17, 223–225. [Google Scholar] [CrossRef]
- Capewell, P.; Krumrie, S.; Katzer, F.; Alexander, C.L.; Weir, W. Molecular Epidemiology of Giardia Infections in the Genomic Era. Trends Parasitol. 2020, 37, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Read, C.M.; Monis, P.T.; Thompson, R.A. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic heterogeneity at the β-giardin locus among human and animal isolates of Giardiaduodenalis and identification of potentially zoonotic subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef]
- Tiangtip, R.; Jongwutiwes, S. Molecular analysis of Cryptosporidium species isolated from HIV-infected patients in Thailand. Trop. Med. Int. Health 2002, 7, 357–364. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System; QGIS Association: Grüt, Switzerland, 2022. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Lebbad, M.; Mattsson, J.G.; Christensson, B.; Ljungström, B.; Backhans, A.; Andersson, J.O.; Svärd, S.G. From mouse to moose: Multilocus genotyping of Giardia isolates from various animal species. Vet. Parasitol. 2010, 168, 231–239. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Fayer, R.; Bern, C.; Gilman, R.H.; Trout, J.M.; Schantz, P.M.; Das, P.; Lal, A.A.; Xiao, L. Triosephosphate Isomerase Gene Characterization and Potential Zoonotic Transmission of Giardia duodenalis. Emerg. Infect. Dis. 2003, 9, 1444–1452. [Google Scholar] [CrossRef]
- Backhans, A.; Jacobson, M.; Hansson, I.; Lebbad, M.; Lambertz, S.T.; Gammelgård, E.; Saager, M.; Akande, O.; Fellström, C. Occurrence of pathogens in wild rodents caught on Swedish pig and chicken farms. Epidemiol. Infect. 2012, 141, 1885–1891. [Google Scholar] [CrossRef]
- Cacciò, S.M.; Beck, R.; Almeida, A.; Bajer, A.; Pozio, E. Identification of Giardia species and Giardia duodenalis assemblages by sequence analysis of the 5.8S rDNA gene and internal transcribed spacers. Parasitology 2010, 137, 919–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U.; Zahedi, A. Molecular epidemiology of giardiasis from a veterinary perspective. Adv. Parasitol. 2019, 106, 209–254. [Google Scholar] [CrossRef]
- Dixon, B.R. Giardia duodenalis in humans and animals—Transmission and disease. Res. Vet.-Sci. 2021, 135, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.; Rivero-Juárez, A.; Santín, M.; George, N.S.; Köster, P.C.; López-López, P.; Risalde, M.A.; García-Bocanegra, I.; Gómez-Villamandos, J.C.; Caballero-Gómez, J.; et al. Diarrhoea-causing enteric protist species in intensively and extensively raised pigs (Sus scrofa domesticus) in Southern Spain. Part I: Prevalence and genetic diversity. Transbound. Emerg. Dis. 2021, 69, e1051–e1064. [Google Scholar] [CrossRef]
- Thompson, R.C.A.; Ash, A. Molecular epidemiology of Giardia and Cryptosporidium infections. Infect. Genet. Evol. 2016, 40, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Wielinga, C.M.; Thompson, R.C.A. Comparative evaluation of Giardia duodenalis sequence data. Parasitology 2007, 134, 1795–1821. [Google Scholar] [CrossRef]
- Tan, T.K.; van Low, V.; Ng, W.H.; Ibrahim, J.; Wang, D.; Tan, C.H.; Chellappan, S.; Lim, Y.A.L. Occurrence of zoonotic Cryptosporidium and Giardia duodenalis species/genotypes in urban rodents. Parasitol. Int. 2019, 69, 110–113. [Google Scholar] [CrossRef]
- Leonhard, S.; Pfister, K.; Beelitz, P.; Wielinga, C.; Thompson, R. The molecular characterisation of Giardia from dogs in southern Germany. Vet.-Parasitol. 2007, 150, 33–38. [Google Scholar] [CrossRef]
- Thompson, R.; Monis, P. Variation in Giardia: Implications for Taxonomy and Epidemiology. Adv. Parasitol. 2004, 58, 69–137. [Google Scholar] [CrossRef]
- Seifollahi, Z.; Sarkari, B.; Motazedian, M.H.; Asgari, Q.; Ranjbar, M.J.; Khabisi, S.A. Protozoan Parasites of Rodents and Their Zoonotic Significance in Boyer-Ahmad District, Southwestern Iran. Vet. Med. Int. 2016, 2016, 1–5. [Google Scholar] [CrossRef]
- Bajer, A. Between-year variation and spatial dynamics of Cryptosporidium spp. and Giardia spp. infections in naturally infected rodent populations. Parasitology 2008, 135, 1629–1649. [Google Scholar] [CrossRef] [PubMed]
- Pita, R.; Mira, A.; Beja, P. Influence of Land Mosaic Composition and Structure on Patchy Populations: The Case of the Water Vole (Arvicola sapidus) in Mediterranean Farmland. PLoS ONE 2013, 8, e69976. [Google Scholar] [CrossRef]
- Bencatel, J.; Sabino-Marques, H.; Álvares, F.; Moura, A.E.; Barbosa, A.M. Atlas de Mamíferos de Portugal, 2nd ed.; Universidade de Évora: Évora, Portugal, 2019. [Google Scholar]
- Čondlová, Š.; Horčičková, M.; Havrdová, N.; Sak, B.; Hlásková, L.; Perec-Matysiak, A.; Kicia, M.; McEvoy, J.; Kváč, M. Diversity of Cryptosporidium spp. in Apodemus spp. in Europe. Eur. J. Protistol. 2019, 69, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bajer, A.; Bednarska, M.; Pawełczyk, A.; Behnke, J.M.; Gilbert, F.S.; Sinski, E. Prevalence and abundance of Cryptosporidium parvum and Giardia spp. in wild rural rodents from the Mazury Lake District region of Poland. Parasitology 2002, 125, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Horčičková, M.; Čondlová, Š.; Holubová, N.; Sak, B.; Květoňová, D.; Hlásková, L.; Konečný, R.; Sedláček, F.; Clark, M.; Giddings, C.; et al. Diversity ofCryptosporidiumin common voles and description ofCryptosporidium alticolissp. n. andCryptosporidium microtisp. n. (Apicomplexa: Cryptosporidiidae). Parasitology 2019, 146, 220–233. [Google Scholar] [CrossRef]
- Myšková, E.; Brož, M.; Fuglei, E.; Kvičerová, J.; Mácová, A.; Sak, B.; Kváč, M.; Ditrich, O. Gastrointestinal parasites of arctic foxes (Vulpes lagopus) and sibling voles (Microtus levis) in Spitsbergen, Svalbard. Parasitol. Res. 2019, 118, 3409–3418. [Google Scholar] [CrossRef]
- Stenger, B.L.S.; Horčičková, M.; Clark, M.E.; Kváč, M.; Čondlová, Š.; Khan, E.; Widmer, G.; Xiao, L.; Giddings, C.W.; Pennil, C.; et al. Cryptosporidium infecting wild cricetid rodents from the subfamilies Arvicolinae and Neotominae. Parasitology 2017, 145, 326–334. [Google Scholar] [CrossRef] [PubMed]
- García-Livia, K.; Martín-Alonso, A.; Foronda, P. Diversity of Cryptosporidium spp. in wild rodents from the Canary Islands, Spain. Parasit. Vectors 2020, 13, 445. [Google Scholar] [CrossRef]
- Ježková, J.; Prediger, J.; Holubová, N.; Sak, B.; Konečný, R.; Feng, Y.; Xiao, L.; Rost, M.; McEvoy, J.; Kváč, M. Cryptosporidium rattin. sp. (Apicomplexa: Cryptosporidiidae) and genetic diversity of Cryptosporidium spp. in brown rats (Rattus norvegicus) in the Czech Republic. Parasitology 2020, 148, 84–97. [Google Scholar] [CrossRef]
- Köster, P.; Dashti, A.; Bailo, B.; Muadica, A.; Maloney, J.; Santín, M.; Chicharro, C.; Migueláñez, S.; Nieto, F.; Cano-Terriza, D.; et al. Occurrence and Genetic Diversity of Protist Parasites in Captive Non-Human Primates, Zookeepers, and Free-Living Sympatric Rats in the Córdoba Zoo Conservation Centre, Southern Spain. Animals 2021, 11, 700. [Google Scholar] [CrossRef]
- Hancke, D.; Suárez, O.V. A review of the diversity of Cryptosporidium in Rattus norvegicus, R. rattus and Mus musculus: What we know and challenges for the future. Acta Trop. 2021, 226, 106244. [Google Scholar] [CrossRef]
- Iseki, M.; Maekawa, T.; Moriya, K.; Uni, S.; Takada, S. Infectivity ofCryptosporidium muris (strain RN 66) in various laboratory animals. Parasitol. Res. 1989, 75, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Kváč, M.; Kestřánová, M.; Květoňová, D.; Kotková, M.; Ortega, Y.; McEvoy, J.; Sak, B. Cryptosporidium tyzzeri and Cryptosporidium muris originated from wild West-European house mice (Mus musculus domesticus) and East-European house mice (Mus musculus musculus) are non-infectious for pigs. Exp. Parasitol. 2012, 131, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Escalante, L.; Yang, C.; Sulaiman, I.; Escalante, A.A.; Montali, R.J.; Fayer, R.; Lal, A.A. Phylogenetic Analysis of Cryptosporidium Parasites Based on the Small-Subunit rRNA Gene Locus. Appl. Environ. Microbiol. 1999, 65, 1578–1583. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yang, W.; Ryan, U.; Zhang, L.; Kvácč, M.; Koudela, B.; Modrý, D.; Li, N.; Fayer, R.; Xiao, L. Development of a Multilocus Sequence Tool for Typing Cryptosporidium muris and Cryptosporidium andersoni. J. Clin. Microbiol. 2011, 49, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Katsumata, T.; Hosea, D.; Uga, S.; Kohno, S.; Ranuh, I.G.; Yanagi, T. Short report: Possible Cryptosporidium muris infection in humans. Am. J. Trop. Med. Hyg. 2000, 62, 70–72. [Google Scholar] [CrossRef]
- Gatei, W.; Ashford, R.W.; Beeching, N.J.; Kamwati, S.K.; Greensill, J.; Hart, C.A. Cryptosporidium muris Infection in an HIV-Infected Adult, Kenya. Emerg. Infect. Dis. 2002, 8, 204–206. [Google Scholar] [CrossRef]
- Palmer, C.J.; Xiao, L.; Terashima, A.; Guerra, H.; Gotuzzo, E.; Saldías, G.; Bonilla, J.A.; Zhou, L.; Lindquist, A.; Upton, S.J. Cryptosporidium muris, a Rodent Pathogen, Recovered from a Human in Perú. Emerg. Infect. Dis. 2003, 9, 1174–1176. [Google Scholar] [CrossRef]
- Muthusamy, D.; Rao, S.S.; Ramani, S.; Monica, B.; Banerjee, I.; Abraham, O.C.; Mathai, D.C.; Primrose, B.; Muliyil, J.; Wanke, C.A.; et al. Multilocus Genotyping of Cryptosporidium sp. Isolates from Human Immunodeficiency Virus-Infected Individuals in South India. J. Clin. Microbiol. 2006, 44, 632–634. [Google Scholar] [CrossRef]
- Guyot, K.; Follet-Dumoulin, A.; Lelièvre, E.; Sarfati, C.; Rabodonirina, M.; Nevez, G.; Cailliez, J.C.; Camus, D.; Dei-Cas, E. Molecular Characterization of Cryptosporidium Isolates Obtained from Humans in France. J. Clin. Microbiol. 2001, 39, 3472–3480. [Google Scholar] [CrossRef]
- Chappell, C.L.; Okhuysen, P.; Tzipori, S.; Widmer, G.; Lupo, P.; Langer-Curry, R.C. Cryptosporidium muris: Infectivity and Illness in Healthy Adult Volunteers. Am. J. Trop. Med. Hyg. 2015, 92, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kváč, M.; Hofmannová, L.; Hlásková, L.; Květoňová, D.; Vítovec, J.; McEvoy, J.; Sak, B. Cryptosporidium erinacei n. sp. (Apicomplexa: Cryptosporidiidae) in hedgehogs. Vet. Parasitol. 2014, 201, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidiumspecies in humans and animals: Current understanding and research needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Animals | Results of Screening No. of positive samples/No. of analyzed samples (Occurrence in % and 95% CI) | Results of Genotyping of Selected Giardia spp.-Positive Samples No. of positive samples/No. of analyzed samples | ||||
| Family | Species (common name) | Cryptosporidium spp. (SSU rRNA PCR) | Giardia spp. (SSU rRNA qPCR) | G. microti (SSU rRNA PCR) | G. microti (bg PCR) | G. microti (gdh PCR) |
| Muridae | Apodemus sylvaticus (wood mouse) | 0/43 | 2/43 (4.65%; 0.01–15.81) | 2/2 | 0/2 | 0/2 |
| Mus spretus (Algerian mouse) | 2/48 (4.17%; 0.00–9.82) | 4/48 (8.33%; 2.32–19.98) | 4/4 | 1/4 | 2/4 | |
| Total | 2/91 (2.20%; 0.27–7.71) | 6/91 (6.60%; 2.46–13.80) | 6/6 | 1/6 | 2/6 | |
| Cricetidae | Arvicola sapidus (southwestern water vole) | 0/52 | 51/52 (98.08%; 89.74–99.95) | 6/8 | 1/8 | 2/8 |
| Microtus cabrerae (Cabrera’s vole) | 1/49 (2.04%; 0.00–10.85) | 2/49 (4.08%; 0.01–13.98) | 1/2 | 1/2 | 0/2 | |
| Microtus lusitanicus (Lusitanian pine vole) | 0/51 | 36/51 (70.59%; 56.17–82.51) | 7/8 | 1/8 | 1/8 | |
| Total | 1/152 (0.66%; 0.00–3.61) | 89/152 (58.55%; 50.29–66.48) | 14/18 | 3/18 | 3/18 | |
| Soricidae | Crocidura russula (greater white-toothed shrew) | 0/47 | 0/47 | - | - | - |
| Total | 3/290 (1.03%; 0.21–2.99) | 95/290 (32.76%; 27.39–38.49) | 20/24 | 4/24 | 5/24 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lux, L.; Ulrich, R.G.; Santos-Silva, S.; Queirós, J.; Imholt, C.; Klotz, C.; Paupério, J.; Pita, R.; Vale-Gonçalves, H.; Alves, P.C.; et al. Detection and Molecular Characterization of Giardia and Cryptosporidium spp. Circulating in Wild Small Mammals from Portugal. Animals 2023, 13, 515. https://doi.org/10.3390/ani13030515
Lux L, Ulrich RG, Santos-Silva S, Queirós J, Imholt C, Klotz C, Paupério J, Pita R, Vale-Gonçalves H, Alves PC, et al. Detection and Molecular Characterization of Giardia and Cryptosporidium spp. Circulating in Wild Small Mammals from Portugal. Animals. 2023; 13(3):515. https://doi.org/10.3390/ani13030515
Chicago/Turabian StyleLux, Laura, Rainer G. Ulrich, Sérgio Santos-Silva, João Queirós, Christian Imholt, Christian Klotz, Joana Paupério, Ricardo Pita, Hélia Vale-Gonçalves, Paulo Célio Alves, and et al. 2023. "Detection and Molecular Characterization of Giardia and Cryptosporidium spp. Circulating in Wild Small Mammals from Portugal" Animals 13, no. 3: 515. https://doi.org/10.3390/ani13030515
APA StyleLux, L., Ulrich, R. G., Santos-Silva, S., Queirós, J., Imholt, C., Klotz, C., Paupério, J., Pita, R., Vale-Gonçalves, H., Alves, P. C., & Mesquita, J. R. (2023). Detection and Molecular Characterization of Giardia and Cryptosporidium spp. Circulating in Wild Small Mammals from Portugal. Animals, 13(3), 515. https://doi.org/10.3390/ani13030515