
Executive Functions as Measured by the Dog Executive Function Scale (DEFS) over the Lifespan of Dogs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Confirmatory Factor Analysis
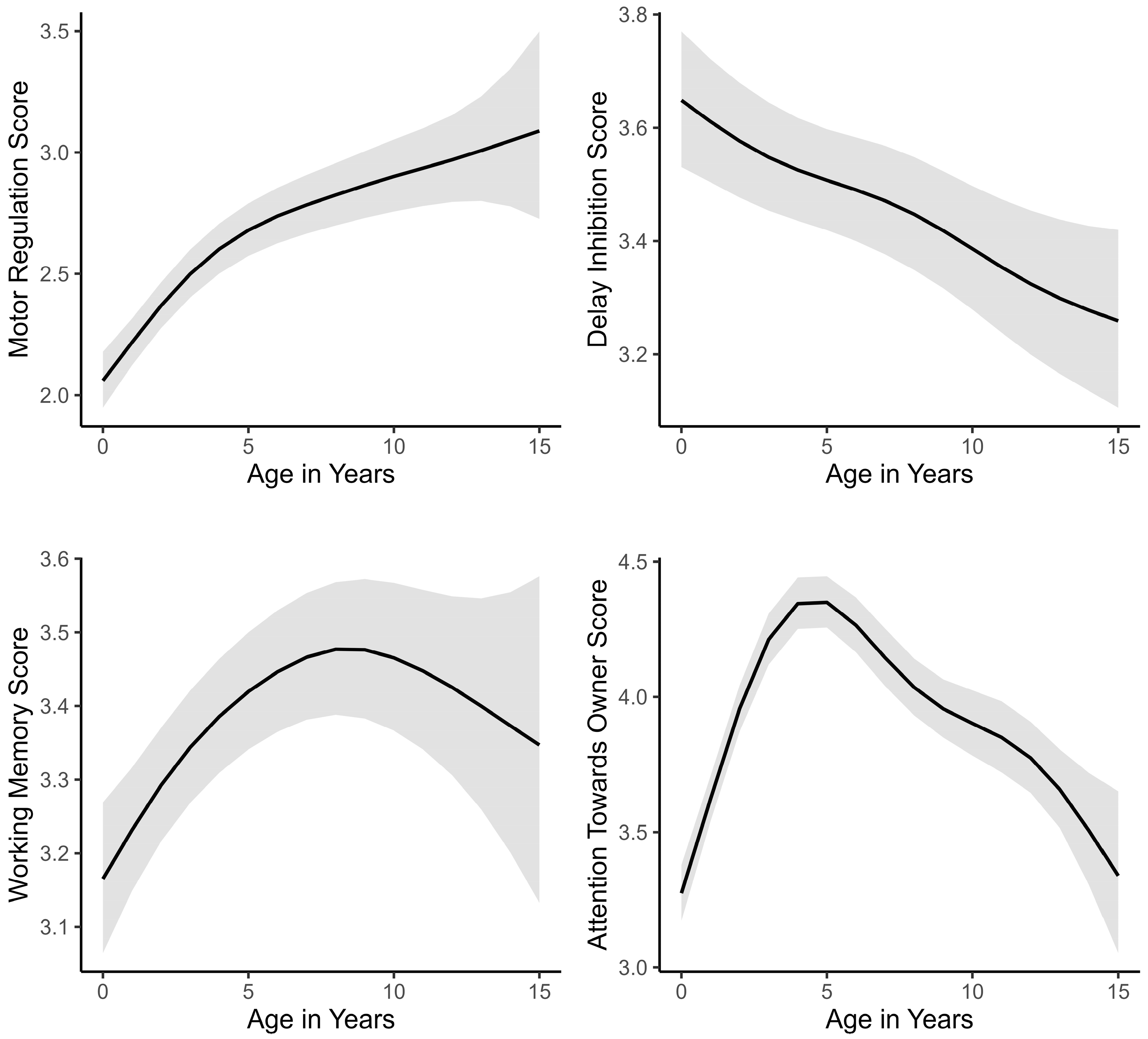
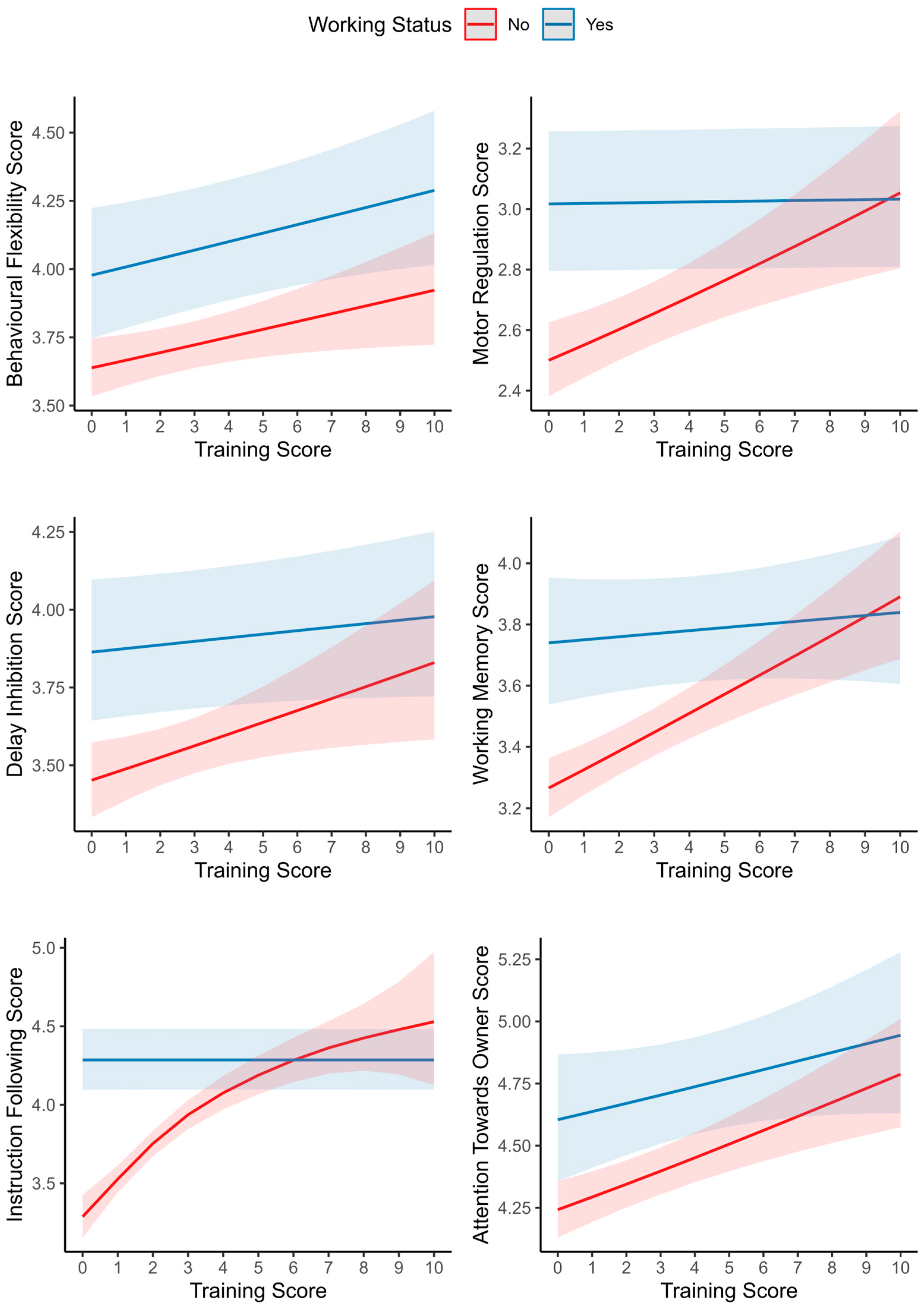
3.2. Life Span Trajectories of Subscale Scores and Other Predictors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diamond, A. Executive functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef] [PubMed]
- Pecora, G.; Zoratto, F.; Paoletti, M.; Bellagamba, F.; Paglieri, F.; Addessi, E. Executive Function. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–5. [Google Scholar]
- Hare, B.; Ferrans, M. Is cognition the secret to working dog success? Anim. Cogn. 2021, 24, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.; Cracknell, N.; Zulch, H.; Mills, D.S. Factors associated with long-term success in working police dogs. Appl. Anim. Behav. Sci. 2018, 207, 67–72. [Google Scholar] [CrossRef]
- Bray, E.E.; MacLean, E.L.; Hare, B.A. Context specificity of inhibitory control in dogs. Anim. Cogn. 2014, 17, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Bray, E.E.; Otto, C.M.; Udell, M.A.; Hall, N.J.; Johnston, A.M.; MacLean, E.L. Enhancing the Selection and Performance of Working Dogs. Front. Vet. Sci. 2021, 8, 430. [Google Scholar] [CrossRef]
- Cohen, E.E.B. A longitudinal study of maternal style, young adult temperament and cognition, and program outcome in guide dogs. Ph.D. Thesis, University of Pennsylvania, Philadelphia, PA, USA, 2018. [Google Scholar]
- Lazarowski, L.; Waggoner, P.; Katz, J.S. The future of detector dog research. Comp. Cogn. Behav. Rev. 2019, 14, 77–80. [Google Scholar] [CrossRef]
- Lazarowski, L.; Strassberg, L.R.; Waggoner, L.; Katz, J.S. Persistence and human-directed behavior in detection dogs: Ontogenetic development and relationships to working dog success. Appl. Anim. Behav. Sci. 2019, 220, 104860. [Google Scholar] [CrossRef]
- Tiira, K.; Tikkanen, A.; Vainio, O. Inhibitory control–Important trait for explosive detection performance in police dogs? Appl. Anim. Behav. Sci. 2020, 224, 104942. [Google Scholar] [CrossRef]
- Troisi, C.A.; Mills, D.S.; Wilkinson, A.; Zulch, H.E. Behavioral and cognitive factors that affect the success of scent detection dogs. Comp. Cogn. Behav. Rev. 2019, 14, 51–76. [Google Scholar] [CrossRef]
- Friedman, N.P.; Miyake, A. Unity and diversity of executive functions: Individual differences as a window on cognitive structure. Cortex 2017, 86, 186–204. [Google Scholar] [CrossRef] [Green Version]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Garon, N.; Bryson, S.E.; Smith, I.M. Executive function in preschoolers: A review using an integrative framework. Psychol. Bull. 2008, 134, 31–60. [Google Scholar] [CrossRef] [PubMed]
- Brydges, C.R.; Fox, A.M.; Reid, C.L.; Anderson, M. The differentiation of executive functions in middle and late childhood: A longitudinal latent-variable analysis. Intelligence 2014, 47, 34–43. [Google Scholar] [CrossRef]
- Gioa, G.; Isquith, P.; Guy, S.; Kenworthy, L. Behavior rating inventory of executive function. Psychol. Assess. Resour. 2000. [Google Scholar]
- Sherman, E.M.; Brooks, B.L. Behavior rating inventory of executive function–preschool version (BRIEF-P): Test review and clinical guidelines for use. Child Neuropsychol. 2010, 16, 503–519. [Google Scholar] [CrossRef]
- Roth, R.M.; Gioia, G.A.; Isquith, P.K. BRIEF-A: Behavior Rating Inventory of Executive Function—Adult Version; Psychological Assessment Resources: 2005. Available online: https://www.parinc.com/Products/Pkey/23 (accessed on 17 January 2023).
- Cotman, C.W.; Head, E. The canine (dog) model of human aging and disease: Dietary, environmental and immunotherapy approaches. J. Alzheimer’s Dis. 2008, 15, 685–707. [Google Scholar] [CrossRef]
- Topál, J.; Miklósi, Á.; Gácsi, M.; Dóka, A.; Pongrácz, P.; Kubinyi, E.; Viranyi, Z.; Csanyi, V. The dog as a model for understanding human social behavior. Adv. Study Behav. 2009, 39, 71–116. [Google Scholar]
- Roth, R.M.; Lance, C.E.; Isquith, P.K.; Fischer, A.S.; Giancola, P.R. Confirmatory factor analysis of the behavior rating inventory of executive function-adult version in healthy adults and application to attention-deficit/hyperactivity disorder. Arch. Clin. Neuropsychol. 2013, 28, 425–434. [Google Scholar] [CrossRef]
- Craik, F.I.; Bialystok, E. Cognition through the lifespan: Mechanisms of change. Trends Cogn. Sci. 2006, 10, 131–138. [Google Scholar] [CrossRef]
- Harada, C.N.; Love, M.C.N.; Triebel, K.L. Normal cognitive aging. Clin. Geriatr. Med. 2013, 29, 737–752. [Google Scholar] [CrossRef]
- Rey-Mermet, A.; Gade, M. Inhibition in aging: What is preserved? What declines? A meta-analysis. Psychon. Bull. Rev. 2018, 25, 1695–1716. [Google Scholar] [CrossRef] [PubMed]
- Veríssimo, J.; Verhaeghen, P.; Goldman, N.; Weinstein, M.; Ullman, M.T. Evidence that ageing yields improvements as well as declines across attention and executive functions. Nat. Hum. Behav. 2022, 6, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Bray, E.E.; Gruen, M.E.; Gnanadesikan, G.E.; Horschler, D.J.; Levy, K.M.; Kennedy, B.S.; Hare, B.A.; MacLean, E.L. Dog cognitive development: A longitudinal study across the first 2 years of life. Anim. Cogn. 2020, 14, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Lazarowski, L.; Krichbaum, S.; Waggoner, L.P.; Katz, J.S. The development of problem-solving abilities in a population of candidate detection dogs (Canis familiaris). Anim. Cogn. 2020, 23, 755–768. [Google Scholar] [CrossRef]
- Adams, B.; Chan, A.; Callahan, H.; Milgram, N.W. The canine as a model of human cognitive aging: Recent developments. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2000, 24, 675–692. [Google Scholar] [CrossRef]
- Adams, B.; Chan, A.; Callahan, H.; Siwak, C.; Tapp, D.; Ikeda-Douglas, C.; Atkinson, P.; Head, E.; Cotman, C.W.; Milgram, N.W. Use of a delayed non-matching to position task to model age-dependent cognitive decline in the dog. Behav. Brain Res. 2000, 108, 47–56. [Google Scholar] [CrossRef]
- Chapagain, D.; Viranyi, Z.; Wallis, L.J.; Huber, L.; Serra, J.; Range, F. Aging of attentiveness in border collies and other pet dog breeds: The protective benefits of lifelong training. Front. Aging Neurosci. 2017, 9, 100. [Google Scholar] [CrossRef]
- Chapagain, D.; Range, F.; Huber, L.; Viranyi, Z. Cognitive aging in dogs. Gerontology 2018, 64, 165–171. [Google Scholar] [CrossRef]
- Chapagain, D.; Viranyi, Z.; Huber, L.; Serra, J.; Schoesswender, J.; Range, F. Effect of age and dietary intervention on discrimination learning in pet dogs. Front. Psychol. 2018, 9, 2217. [Google Scholar] [CrossRef]
- Davis, P.R.; Giannini, G.; Rudolph, K.; Calloway, N.; Royer, C.M.; Beckett, T.L.; Murphy, M.; Bresch, F.; Pagani, D.; Platt, T.; et al. Abeta vaccination in combination with behavioral enrichment in aged beagles: Effects on cognition, Abeta, and microhemorrhages. Neurobiol. Aging 2017, 49, 86–99. [Google Scholar] [CrossRef]
- Milgram, N.; Head, E.; Zicker, S.; Ikeda-Douglas, C.; Murphey, H.; Muggenburg, B.; Siwak, C.; Tapp, D.; Cotman, C. Learning ability in aged beagle dogs is preserved by behavioral enrichment and dietary fortification: A two-year longitudinal study. Neurobiol. Aging 2005, 26, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Tapp, P.; Siwak, C.T.; Estrada, J.; Head, E.; Muggenburg, B.A.; Cotman, C.W.; Milgram, N.W. Size and reversal learning in the beagle dog as a measure of executive function and inhibitory control in aging. Learn. Mem. 2003, 10, 64–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watowich, M.M.; MacLean, E.L.; Hare, B.; Call, J.; Kaminski, J.; Miklósi, Á.; Snyder-Mackler, N. Age influences domestic dog cognitive performance independent of average breed lifespan. Anim. Cogn. 2020, 23, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Holst, Y.; Thorell, L.B. Adult executive functioning inventory (ADEXI): Validity, reliability, and relations to ADHD. Int. J. Methods Psychiatr. Res. 2018, 27, e1567. [Google Scholar] [CrossRef]
- Isquith, P.K.; Roth, R.M.; Gioia, G. Contribution of rating scales to the assessment of executive functions. Appl. Neuropsychol. Child 2013, 2, 125–132. [Google Scholar] [CrossRef]
- Thorell, L.B.; Nyberg, L. The Childhood Executive Functioning Inventory (CHEXI): A new rating instrument for parents and teachers. Dev. Neuropsychol. 2008, 33, 536–552. [Google Scholar] [CrossRef]
- Foraita, M.; Howell, T.; Bennett, P. Development of the dog executive function scale (DEFS) for adult dogs. Animal Cognition 2022, 25, 1479–1491. [Google Scholar] [CrossRef]
- Wright, H.F.; Mills, D.S.; Pollux, P.M. Development and validation of a psychometric tool for assessing impulsivity in the domestic dog (Canis familiaris). Int. J. Comp. Psychol. 2011, 24, 210–225. [Google Scholar] [CrossRef]
- Vas, J.; Topal, J.; Pech, E.; Miklosi, A. Measuring attention deficit and activity in dogs: A new application and validation of a human ADHD questionnaire. Appl. Anim. Behav. Sci. 2007, 103, 105–117. [Google Scholar] [CrossRef]
- Holgado–Tello, F.P.; Chacón–Moscoso, S.; Barbero–García, I.; Vila–Abad, E. Polychoric versus Pearson correlations in exploratory and confirmatory factor analysis of ordinal variables. Qual. Quant. 2010, 44, 153–166. [Google Scholar] [CrossRef]
- Hooper, D.; Coughlan, J.; Mullen, M.R. Structural equation modelling: Guidelines for determining model fit. Electron. J. Bus. Res. Methods 2008, 6, 53–60. [Google Scholar]
- Hastie, T.; Tibshirani, R. Generalized additive models: Some applications. J. Am. Stat. Assoc. 1987, 82, 371–386. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Cattell, R.B. Fixing the number of factors: The most practicable psychometric procedures. In The Scientific Use of Factor Analysis in Behavioral and Life Sciences; Springer: Berlin/Heidelberg, Germany, 1978; pp. 72–91. [Google Scholar]
- Gorsuch, R.L. Factor Analysis; Taylor & Francis Group: London, UK, 1983. [Google Scholar]
- Overall, K.L. Manual of Clinical Behavioral Medicine for Dogs and Cats; Saunders: St. Louis, MO, USA, 2013. [Google Scholar]
- Scott, J.; Fuller, J. Genetics and the Social Behavior of the Dog; The University of Chicago Press: Chicago, IL, USA, 1965. [Google Scholar]
- Lazarowski, L.; Thompkins, A.; Krichbaum, S.; Waggoner, L.P.; Deshpande, G.; Katz, J.S. Comparing pet and detection dogs (Canis familiaris) on two aspects of social cognition. Learn. Behav. 2020, 48, 432–443. [Google Scholar] [CrossRef] [PubMed]
- RSPCA: Caring for Your Puppy. Available online: https://www.rspca.org.uk/adviceandwelfare/pets/dogs/health/puppycare (accessed on 16 December 2022).
- Bray, E.E.; Gruen, M.E.; Gnanadesikan, G.E.; Horschler, D.J.; Levy, K.M.; Kennedy, B.S.; Hare, B.A.; MacLean, E.L. Cognitive characteristics of 8-to 10-week-old assistance dog puppies. Anim. Behav. 2020, 166, 193–206. [Google Scholar] [CrossRef]
- Foraita, M.; Howell, T.; Bennett, P. Working memory and inhibition in 7.5-week-old Seeing Eye Dog puppies. Appl. Anim. Behav. Sci. 2021, 241, 105394. [Google Scholar] [CrossRef]
- Alloway, T.P.; Alloway, R.G. Working memory across the lifespan: A cross-sectional approach. J. Cogn. Psychol. 2013, 25, 84–93. [Google Scholar] [CrossRef]
- Wallis, L.J.; Range, F.; Muller, C.A.; Serisier, S.; Huber, L.; Viranyi, Z. Lifespan development of attentiveness in domestic dogs: Drawing parallels with humans. Front. Psychol. 2014, 5, 71. [Google Scholar] [CrossRef]
- Verhaeghen, P. Aging and executive control: Reports of a demise greatly exaggerated. Curr. Dir. Psychol. Sci. 2011, 20, 174–180. [Google Scholar] [CrossRef]
- Hasher, L.; Stoltzfus, E.R.; Zacks, R.T.; Rypma, B. Age and inhibition. J. Exp. Psychol. Learn. Mem. Cogn. 1991, 17, 163. [Google Scholar] [CrossRef]
- Persad, C.C.; Abeles, N.; Zacks, R.T.; Denburg, N.L. Inhibitory changes after age 60 and their relationship to measures of attention and memory. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2002, 57, P223–P232. [Google Scholar] [CrossRef] [PubMed]
- Fong, M.C.-M.; Law, T.S.-T.; Ma, M.K.-H.; Hui, N.Y.; Wang, W.S. Can inhibition deficit hypothesis account for age-related differences in semantic fluency? Converging evidence from Stroop color and word test and an ERP flanker task. Brain Lang. 2021, 218, 104952. [Google Scholar] [CrossRef] [PubMed]
- Zanghi, B.M.; Kerr, W.; de Rivera, C.; Araujo, J.A.; Milgram, N.W. Effect of age and feeding schedule on diurnal rest/activity rhythms in dogs. J. Vet. Behav. 2012, 7, 339–347. [Google Scholar] [CrossRef]
- Bray, E.E.; Sammel, M.D.; Seyfarth, R.M.; Serpell, J.A.; Cheney, D.L. Temperament and problem solving in a population of adolescent guide dogs. Anim. Cogn. 2017, 20, 923–939. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.; Ling, D.S. Conclusions about interventions, programs, and approaches for improving executive functions that appear justified and those that, despite much hype, do not. Dev. Cogn. Neurosci. 2016, 18, 34–48. [Google Scholar] [CrossRef]
- Diamond, A.; Ling, D.S. Review of the evidence on, and fundamental questions about, efforts to improve executive functions, including working memory. In Cognitive and Working Memory Training: Perspectives from Psychology, Neuroscience, and Human Development; Novick, J.M., Bunting, M.F., Dougherty, M.R., Engle, R.W., Eds.; Oxford University Press: Oxford, UK, 2020; pp. 143–431. [Google Scholar]
- Foraita, M.; Howell, T.; Bennett, P. Environmental influences on development of executive functions in dogs. Anim. Cogn. 2021, 24, 655–675. [Google Scholar] [CrossRef] [PubMed]
- Junttila, S.; Huohvanainen, S.; Tiira, K. Effect of sex and reproductive status on inhibitory control and social cognition in the domestic dog (Canis familiaris). Animals 2021, 11, 2448. [Google Scholar] [CrossRef] [PubMed]
- Scandurra, A.; Marinelli, L.; Looke, M.; D’Aniello, B.; Mongillo, P. The effect of age, sex and gonadectomy on dogs’ use of spatial navigation strategies. Appl. Anim. Behav. Sci. 2018, 205, 89–97. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Juvenile Dog Sample (n = 129) | Senior Dog Sample (n = 127) | |||||
|---|---|---|---|---|---|---|
| DEFS | Orthogonal | Single | DEFS | Orthogonal | Single | |
| df | 215.00 | 230.00 | 230.00 | 215.00 | 230.00 | 230.00 |
| χ2 | 238.45 | 1317.31 | 750.64 | 243.09 | 1289.53 | 1035.07 |
| (χ2) p | 0.13 | 0.00 | 0.00 | 0.09 | 0.00 | 0.00 |
| RSMEA | 0.03 | 0.19 | 0.13 | 0.03 | 0.19 | 0.17 |
| CFI | 0.99 | 0.65 | 0.83 | 0.99 | 0.71 | 0.78 |
| TLI | 0.99 | 0.61 | 0.81 | 0.99 | 0.68 | 0.75 |
| SRMR | 0.08 | 0.18 | 0.12 | 0.08 | 0.18 | 0.15 |
| Latent Variable | Indicator | B (Unstandardised Estimates) | β (Standardised Estimates) | SE | Z | ||||
|---|---|---|---|---|---|---|---|---|---|
| Juvenile | Senior | Juvenile | Senior | Juvenile | Senior | Juvenile | Senior | ||
| Behavioural Flexibility | BF1 | 1.000 | 1.00 | 0.510 | 0.422 | 0.000 | 0.000 | ||
| BF2 | 1.651 | 2.128 | 0.842 | 0.897 | 0.292 | 0.501 | 5.645 | 4.246 | |
| BF3 | 1.037 | 1.676 | 0.529 | 0.706 | 0.205 | 0.379 | 5.056 | 4.427 | |
| BF4 | 1.618 | 2.055 | 0.825 | 0.866 | 0.274 | 0.485 | 5.903 | 4.235 | |
| Motor Regulation | MR1 | 1.000 | 1 | 0.498 | 0.387 | 0.000 | 0.000 | ||
| MR2 | 1.818 | 2.399 | 0.906 | 0.928 | 0.281 | 0.507 | 6.471 | 4.729 | |
| MR3 | 1.744 | 2.151 | 0.869 | 0.832 | 0.285 | 0.485 | 6.131 | 4.435 | |
| MR4 | 1.496 | 1.932 | 0.745 | 0.748 | 0.254 | 0.438 | 5.885 | 4.412 | |
| Delay Inhibition | DI1 | 1.000 | 1 | 0.737 | 0.589 | 0.000 | 0.000 | ||
| DI2 | 0.890 | 0.664 | 0.656 | 0.585 | 0.159 | 0.113 | 5.611 | 5.871 | |
| DI3 | 0.655 | 0.660 | 0.483 | 0.685 | 0.136 | 0.092 | 4.817 | 7.179 | |
| DI4 | 1.051 | 0.773 | 0.775 | 0.693 | 0.203 | 0.110 | 5.165 | 7.056 | |
| Working Memory | WM1 | 1.000 | 1.000 | 0.518 | 0.579 | 0.000 | 0.000 | ||
| WM2 | 1.381 | 0.835 | 0.715 | 0.637 | 0.250 | 0.166 | 5.529 | 5.023 | |
| WM3 | 1.152 | 0.919 | 0.597 | 0.016 | 0.213 | 0.168 | 5.404 | 5.483 | |
| WM4 | 0.370 | 0.023 | 0.192 | 0.422 | 0.193 | 0.153 | 1.912 | 0.147 | |
| Instruction Following | IF1 | 1.000 | 1.000 | 0.422 | 0.660 | 0.000 | 0.000 | ||
| IF2 | 1.320 | 1.097 | 0.557 | 0.724 | 0.359 | 0.137 | 3.678 | 7.989 | |
| IF3 | 1.832 | 1.225 | 0.773 | 0.808 | 0.496 | 0.172 | 3.692 | 7.127 | |
| IF4 | 1.033 | 0.790 | 0.436 | 0.522 | 0.351 | 0.136 | 2.944 | 5.810 | |
| Attention Towards Owner | AO1 | 1.000 | 1.000 | 0.746 | 0.918 | 0.000 | 0.000 | ||
| AO2 | 1.118 | 0.893 | 0.834 | 0.819 | 0.112 | 0.079 | 10.010 | 11.323 | |
| AO3 | 1.090 | 0.820 | 0.813 | 0.753 | 0.085 | 0.075 | 12.861 | 10.912 | |
| Behavioural Flexibility | Motor Regulation | Delay Inhibition | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Predictors | Estimates | CI | p | Estimates | CI | p | Estimates | CI | p |
| (Intercept) | 3.99 | 3.82–4.17 | <0.001 | 3.08 | 2.89–3.28 | <0.001 | 3.49 | 3.33–3.66 | <0.001 |
| Working status: No | Reference | ||||||||
| Working status: Yes | 1.09 | 1.04–1.15 | 0.001 | 1.14 | 1.06–1.22 | <0.001 | 1.09 | 1.04–1.16 | 0.001 |
| Sex (Female/intact) | Reference | ||||||||
| Sex (Female/spayed) | 0.96 | 0.91–1.01 | 0.102 | 0.91 | 0.85–0.98 | 0.011 | 1.03 | 0.97–1.08 | 0.344 |
| Sex (Male/desexed) | 0.93 | 0.89–0.98 | 0.005 | 0.84 | 0.78–0.90 | <0.001 | 1.01 | 0.96–1.07 | 0.593 |
| Sex (Male/intact) | 0.98 | 0.93–1.04 | 0.504 | 0.91 | 0.84–0.99 | 0.022 | 1.00 | 0.95–1.07 | 0.874 |
| Smooth term (Years) | 0.672 | <0.001 | <0.001 | ||||||
| Smooth term (Training Score) | 0.011 | 0.486 | 0.487 | ||||||
| Tensor product interaction (Years, Training Score) | 0.519 | 0.162 | 0.122 | ||||||
| Smooth term (Training Score) xWorking status: No | 0.256 | <0.001 | 0.019 | ||||||
| Smooth term (Training Score) × Working status: Yes | 0.820 | 0.916 | 0.519 | ||||||
| Observations | 954 | 954 | 954 | ||||||
| R2 | 0.032 | 0.152 | 0.031 | ||||||
| Deviance | 3.03% | 14.6% | 2.98% | ||||||
| Working Memory | Instruction Following | Attention Towards Owner | |||||||
| Predictors | Estimates | CI | p | Estimates | CI | p | Estimates | CI | p |
| (Intercept) | 3.55 | 3.42–3.69 | <0.001 | 3.71 | 3.57–3.85 | <0.001 | 4.03 | 3.89–4.16 | <0.001 |
| Working status: No | Reference | ||||||||
| Working status: Yes | 1.10 | 1.05–1.14 | <0.001 | 1.11 | 1.07–1.16 | <0.001 | 1.07 | 1.03–1.11 | 0.001 |
| Sex (Female/intact) | Reference | ||||||||
| Sex (Female/spayed) | 0.99 | 0.94–1.03 | 0.506 | 1.04 | 1.00–1.08 | 0.062 | 1.00 | 0.96–1.04 | 0.944 |
| Sex (Male/desexed) | 0.96 | 0.92–1.00 | 0.044 | 1.04 | 1.00–1.08 | 0.083 | 0.98 | 0.95–1.02 | 0.353 |
| Sex (Male/intact) | 1.02 | 0.98–1.07 | 0.317 | 1.00 | 0.95–1.05 | 0.943 | 0.98 | 0.94–1.03 | 0.462 |
| Smooth term (Years) | <0.001 | 0.924 | <0.001 | ||||||
| Smooth term (Training Score) | 0.178 | 0.708 | 0.011 | ||||||
| Tensor product interaction (Years, Training Score) | 0.075 | 0.482 | 0.127 | ||||||
| Smooth term (Training Score) × Working status: No | <0.001 | <0.001 | 0.047 | ||||||
| Smooth term (Training Score) × Working status: Yes | 0.581 | 0.723 | 0.950 | ||||||
| Observations | 954 | 954 | 954 | ||||||
| R2 | 0.082 | 0.157 | 0.246 | ||||||
| Deviance | 8.07% | 13.1% | 24.7% | ||||||
| Behavioural Flexibility | Motor Regulation | Working Memory | |
|---|---|---|---|
| Female spayed/Female intact | 0.0743 | 0.64515 | 0.551 |
| Female spayed/Male desexed | 0.0774 | 0.00046 | 0.137 |
| Female spayed/Male intact | 0.2216 | 0.02176 | 0.386 |
| Female intact/Male desexed | 0.0041 | 0.11774 | 0.496 |
| Female intact/Male intact | 0.3591 | 0.23522 | 0.298 |
| Male desexed/Male intact | 0.0111 | 0.79451 | 0.047 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foraita, M.; Howell, T.; Bennett, P. Executive Functions as Measured by the Dog Executive Function Scale (DEFS) over the Lifespan of Dogs. Animals 2023, 13, 533. https://doi.org/10.3390/ani13030533
Foraita M, Howell T, Bennett P. Executive Functions as Measured by the Dog Executive Function Scale (DEFS) over the Lifespan of Dogs. Animals. 2023; 13(3):533. https://doi.org/10.3390/ani13030533
Chicago/Turabian StyleForaita, Maike, Tiffani Howell, and Pauleen Bennett. 2023. "Executive Functions as Measured by the Dog Executive Function Scale (DEFS) over the Lifespan of Dogs" Animals 13, no. 3: 533. https://doi.org/10.3390/ani13030533
APA StyleForaita, M., Howell, T., & Bennett, P. (2023). Executive Functions as Measured by the Dog Executive Function Scale (DEFS) over the Lifespan of Dogs. Animals, 13(3), 533. https://doi.org/10.3390/ani13030533