
Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicus



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Animals and Rearing Environment
2.2. Neuropeptide Synthesis
2.3. Locomotion Behavior Video Capture
2.4. Data Analysis of Locomotion Behavior
2.5. Muscle Metabolomics
3. Results
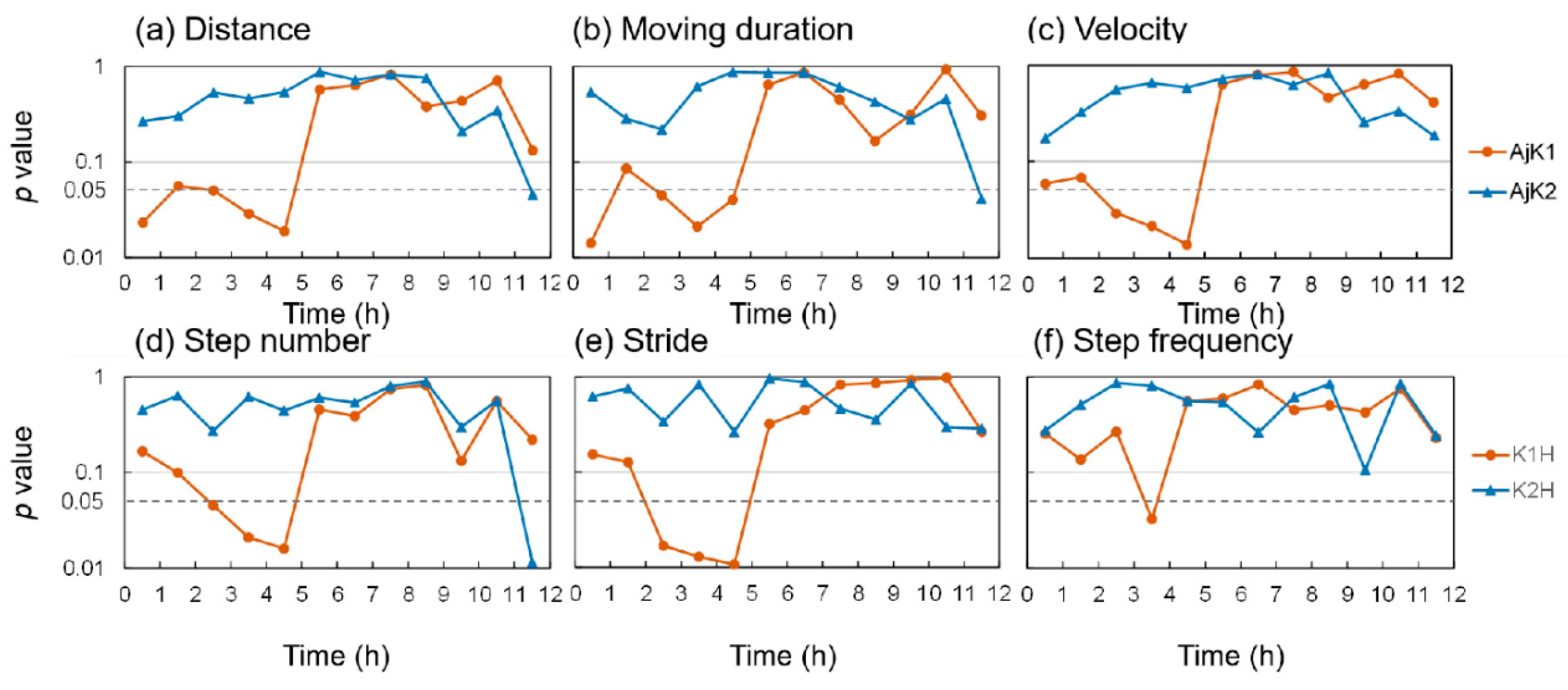
3.1. Changes in Locomotion of A. japonicus after KPs Injection
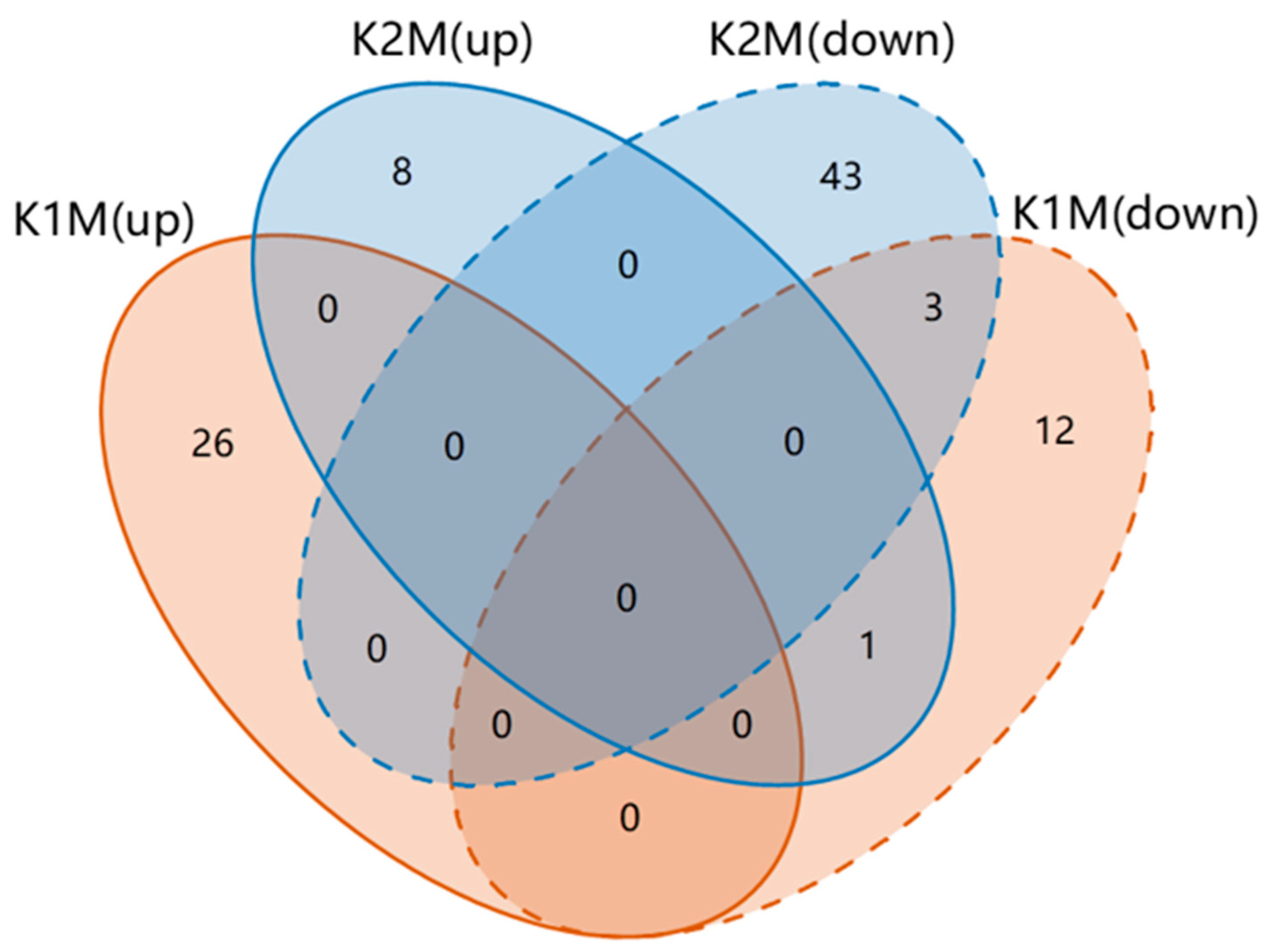
3.2. KPs Induced Changes in Muscle Metabolism
3.3. Differential Metabolites and Metabolic Pathways under KPs Treatment
4. Discussion
4.1. KPs More Likely to Inhibit Locomotion of A. japonicus
4.2. Mechanisms by Which AjK1 Inhibits Locomotor Behavior of A. japonicus
4.3. Possible Functions of the Neuropeptide AjK2
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, K.; Zhang, L.; Zhang, T.; Yang, H.; Brinkman, R. The effect of melatonin on locomotor behavior and muscle physiology in the sea cucumber Apostichopus japonicus. Front. Physiol. 2019, 10, 221. [Google Scholar] [CrossRef] [Green Version]
- Suwansa-Ard, S.; Chaiyamoon, A.; Talarovicova, A.; Tinikul, R.; Tinikul, Y.; Poomtong, T.; Elphick, M.R.; Cummins, S.F.; Sobhon, P. Transcriptomic discovery and comparative analysis of neuropeptide precursors in sea cucumbers (Holothuroidea). Peptides 2018, 99, 231–240. [Google Scholar] [CrossRef]
- Semmens, D.C.; Mirabeau, O.; Moghul, I.; Pancholi, M.R.; Wurm, Y.; Elphick, M.R. Transcriptomic identification of starfish neuropeptide precursors yields new insights into neuropeptide evolution. Open Biol. 2016, 6, 150224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Süudhof, T.C. Neurotransmitter Release. Handb. Exp. Pharmacol. 2008, 184, 1–21. [Google Scholar] [CrossRef]
- Hökfelt, T.; Xu, Z.-Q.D.; Sergeyev, V.; Ubink, R.; Diez, M. Neuropeptides—An overview. Neuropharmacology 2000, 39, 1337–1356. [Google Scholar] [CrossRef] [PubMed]
- Maurice, R.; Elphick; Olivier; Mirabeau. The Evolution and Variety of RFamide-Type Neuropeptides: Insights from Deuterostomian Invertebrates. Front. Endocrinol. 2014, 5, 93. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Cao, Z.; Shen, Z.; Yang, J.; Chen, X.; Yang, Z.; Xu, K.; Xiang, X.; Yu, Q.; Song, Y.; et al. Existence and functions of a kisspeptin neuropeptide signaling system in a non-chordate deuterostome species. elife 2020, 9, e53370. [Google Scholar] [CrossRef]
- Suzuki, S.; Kadokawa, H.; Hashizume, T. Direct kisspeptin-10 stimulation on luteinizing hormone secretion from bovine and porcine anterior pituitary cells. Anim. Reprod. Sci. 2008, 103, 360–365. [Google Scholar] [CrossRef]
- Ohtaki, T.; Shintani, Y.; Honda, S.; Matsumoto, H.; Hori, A.; Kanehashi, K.; Terao, Y.; Kumano, S.; Takatsu, Y.; Masuda, Y.; et al. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001, 411, 613–617. [Google Scholar] [CrossRef]
- Roa, J.; Aguilar, E.; Dieguez, C.; Pinilla, L.; Tena-Sempere, M. New frontiers in kisspeptin/GPR54 physiology as fundamental gatekeepers of reproductive function. Front. Neuroendocrinol. 2008, 29, 48–69. [Google Scholar] [CrossRef]
- Hudson, A.D.; Kauffman, A.S. Metabolic actions of kisspeptin signaling: Effects on body weight, energy expenditure, and feeding. Pharmacol. Ther. 2022, 231, 107974. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Kim, C.H.; Go, H.J.; Egertova, M.; Zampronio, C.G.; Jones, A.M.; Park, N.G.; Elphick, M.R. Biochemical, Anatomical, and Pharmacological Characterization of Calcitonin-Type Neuropeptides in Starfish: Discovery of an Ancient Role as Muscle Relaxants. Front. Neurosci. 2018, 12, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Hamel, J.F.; Mercier, A. The Sea Cucumber Apostichopus Japonicus: History, Biology and Aquaculture; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Chen, M.; Talarovicova, A.; Zheng, Y.; Storey, K.B.; Elphick, M.R. Neuropeptide precursors and neuropeptides in the sea cucumber Apostichopus japonicus: A genomic, transcriptomic and proteomic analysis. Sci. Rep. 2019, 9, 8829. [Google Scholar] [CrossRef] [Green Version]
- Domenici, P. The scaling of locomotor performance in predator–prey encounters: From fish to killer whales. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 131, 169–182. [Google Scholar] [CrossRef]
- Zhou, M.; Dorland, R.D. Aggregation and vertical migration behavior of Euphausia superba. Deep Sea Res. Part II Top. Stud. Oceanogr. 2004, 51, 2119–2137. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, L.; Fan, X.; Guo, X.; Yang, H. The effect of pedal peptide-type neuropeptide on locomotor behavior and muscle physiology in the sea cucumber Apostichopus japonicus. Front. Physiol. 2020, 11, 559348. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, L.B.; Lin, C.G.; Sun, J.M.; Kan, R.T.; Yang, H.S. Influence of flow velocity on motor behavior of sea cucumber Apostichopus japonicus. Physiol. Behav. 2015, 144, 52–59. [Google Scholar] [CrossRef]
- Rowe, M.L.; Elphick, M.R. The neuropeptide transcriptome of a model echinoderm, the sea urchin Strongylocentrotus purpuratus. Gen. Comp. Endocrinol. 2012, 179, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Xu, R.N.X.; Fan, L.M.; Rieser, M.J.; El-Shourbagy, T.A. Recent advances in high-throughput quantitative bioanalysis by LC-MS/MS. J. Pharm. Biomed. Anal. 2007, 44, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Ru, X.; Zhang, L.; Liu, S.; Yang, H. Reproduction affects locomotor behaviour and muscle physiology in the sea cucumber, Apostichopus japonicus. Anim. Behav. 2017, 133, 223–228. [Google Scholar] [CrossRef]
- Kato, S.; Tsurumaru, S.; Taga, M.; Yamane, T.; Shibata, Y.; Ohno, K.; Fujiwara, A.; Yamano, K.; Yoshikuni, M. Neuronal peptides induce oocyte maturation and gamete spawning of sea cucumber, Apostichopus japonicus. Dev. Biol. 2009, 326, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B. Pharmacology, 8th ed.; People’s Medical Publishing House: Beijing, China, 2013; p. 484. [Google Scholar]
- Li, L.; Chen, M.Y.; Storey, K.B. Metabolic response of longitudinal muscles to acute hypoxia in sea cucumber Apostichopus japonicus (Selenka): A metabolome integrated analysis. Comp. Biochem. Physiol. D-Genom. Proteom. 2019, 29, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Pan, Y. Quantitative Research on Motor Behavioral Rhythms of Sea Cucumber Apostichopus japonicus (Selenka). Ph.D. Thesis, University of Chinese Academy of Sciences (Institute of Oceanology, Cas), Qingdao, China, 2015. [Google Scholar]
- Padilla, S.L.; Perez, J.G.; Ben-Hamo, M.; Johnson, C.W.; Sanchez, R.E.A.; Bussi, I.L.; Palmiter, R.D.; de la Iglesia, H.O. Kisspeptin neurons in the arcuate nucleus of the hypothalamus orchestrate circadian rhythms and metabolism. Curr. Biol. 2019, 29, 592–604. [Google Scholar] [CrossRef] [Green Version]
- Csabafi, K.; Jaszberenyi, M.; Bagosi, Z.; Liptak, N.; Telegdy, G. Effects of kisspeptin-13 on the hypothalamic-pituitary-adrenal axis, thermoregulation, anxiety and locomotor activity in rats. Behav. Brain Res. 2013, 241, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Ibos, K.E.; Bodnar, E.; Bagosi, Z.; Bozso, Z.; Toth, G.; Szabo, G.; Csabafi, K. Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats. Biomedicines 2021, 9, 112. [Google Scholar] [CrossRef]
- Gibula-Bruzda, E.; Marszalek-Grabska, M.; Gawel, K.; Trzcinska, R.; Silberring, J.; Kotlinska, J.H. The new kisspeptin derivative—kissorphin (KSO)—attenuates acute hyperlocomotion and sensitization induced by ethanol and morphine in mice. Alcohol 2017, 64, 45–53. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, L.; Fan, X.; Zhuo, P.; Feng, Q.; Zhang, S.; Guo, X.; Liu, X. Influence of an L-type SALMFamide neuropeptide on locomotory performance and muscle physiology in the sea cucumber Apostichopus japonicus. J. Exp. Biol. 2021, 224, jeb242566. [Google Scholar] [CrossRef]
- Kahsai, L.; Martin, J.R.; Winther, A.M.E. Neuropeptides in the Drosophila central complex in modulation of locomotor behavior. J. Exp. Biol. 2010, 213, 2256–2265. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Zhao, Z. Structure and function of neuropeptide F in insects. J. Integr. Agric. 2020, 19, 1429–1438. [Google Scholar] [CrossRef]
- Hou, L.; Yang, P.; Jiang, F.; Liu, Q.; Wang, X.; Kang, L. The neuropeptide F/nitric oxide pathway is essential for shaping locomotor plasticity underlying locust phase transition. Elife 2017, 6, e22526. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.M.; Schlichting, M.; Abruzzi, K.C.; Long, X.; Rosbash, M. Allatostatin-C/AstC-R2 is a novel pathway to modulate the circadian activity pattern in Drosophila. Curr. Biol. 2019, 29, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, B.; Szlanka, T.; Zvara, A.; Zurovec, M.; Sery, M.; Kakas, S.; Ramasz, B.; Hegedus, Z.; Lukacsovich, T.; Puskas, L.; et al. Selective elimination/RNAi silencing of FMRF-related peptides and their receptors decreases the locomotor activity in Drosophila melanogaster. Gen. Comp. Endocrinol. 2013, 191, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Romanowski, A.; Meelkop, E.; Caldart, C.S.; Schoofs, L.; Golombek, D.A. Pigment-dispersing factor signaling in the circadian system of Caenorhabditis elegans. Genes Brain Behav. 2015, 14, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Conzelmann, M.; Offenburger, S.-L.; Asadulina, A.; Keller, T.; Muench, T.A.; Jekely, G. Neuropeptides regulate swimming depth of Platynereis larvae. Proc. Natl. Acad. Sci. USA 2011, 108, E1174–E1183. [Google Scholar] [CrossRef] [Green Version]
- Nakamachi, T.; Matsuda, K.; Maruyama, K.; Miura, T.; Uchiyama, M.; Funahashi, H.; Sakurai, T.; Shioda, S. Regulation by orexin of feeding behaviour and locomotor activity in the goldfish. J. Neuroendocrinol. 2006, 18, 290–297. [Google Scholar] [CrossRef]
- Abbott, M.; Volkoff, H. Thyrotropin Releasing Hormone (TRH) in goldfish (Carassius auratus): Role in the regulation of feeding and locomotor behaviors and interactions with the orexin system and cocaine- and amphetamine regulated transcript (CART). Horm. Behav. 2011, 59, 236–245. [Google Scholar] [CrossRef]
- Matsuda, K. Regulation of feeding behavior and psychomotor activity by corticotropin-releasing hormone (CRH) in fish. Front. Neurosci. 2013, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Fujiki, N.; Yoshida, Y.; Ripley, B.; Mignot, E.; Nishino, S. Effects of IV and ICV hypocretin-1 (Orexin A) in hypocretin receptor-2 gene mutated narcoleptic dogs and IV hypocretin-1 replacement therapy in a hypocretin-ligand-deficient narcoleptic dog. Sleep 2003, 26, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, A.; Tsujino, N.; Funahashi, H.; Honda, K.; Guan, J.L.; Wang, Q.P.; Tominaga, M.; Goto, K.; Shioda, S.; Sakurai, T. Orexins activate histaminergic neurons via the orexin 2 receptor. Biochem. Biophys. Res. Commun. 2002, 290, 1237–1245. [Google Scholar] [CrossRef]
- Nakamura, T.; Uramura, K.; Nambu, T.; Yada, T.; Goto, K.; Yanagisawa, M.; Sakurai, T. Orexin-induced hyperlocomotion and stereotypy are mediated by the dopaminergic system. Brain Res. 2000, 873, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Yokogawa, T.; Marin, W.; Faraco, J.; Pezeron, G.; Appelbaum, L.; Zhang, J.; Rosa, F.; Mourrain, P.; Mignot, E. Characterization of sleep in zebrafish and insomnia in hypocretin receptor mutants. PLoS Biol. 2007, 5, 2379–2397. [Google Scholar] [CrossRef] [PubMed]
- Paneda, C.; Huitron-Resendiz, S.; Frago, L.M.; Chowen, J.A.; Picetti, R.; de Lecea, L.; Roberts, A.J. Neuropeptide S reinstates cocaine-seeking behavior and increases locomotor activity through corticotropin-releasing factor receptor 1 in mice. J. Neurosci. 2009, 29, 4155–4161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibellini, F.; Smith, T.K. The Kennedy pathway—De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life 2010, 62, 414–428. [Google Scholar] [CrossRef]
- Lee, S.; Norheim, F.; Gulseth, H.L.; Langleite, T.M.; Aker, A.; Gundersen, T.E.; Holen, T.; Birkeland, K.I.; Drevon, C.A. Skeletal muscle phosphatidylcholine and phosphatidylethanolamine respond to exercise and influence insulin sensitivity in men. Sci. Rep. 2018, 8, 6531. [Google Scholar] [CrossRef] [Green Version]
- Takagi, A. Lipid comosition of arcoplasmic reticulum of human skeletal muscle. Biochim. Et Biophys. Acta (BBA)-Lipids Lipid Metab. 1971, 248, 12–20. [Google Scholar] [CrossRef]
- Funai, K.; Lodhi, I.J.; Spears, L.D.; Yin, L.; Song, H.; Klein, S.; Semenkovich, C.F. Skeletal Muscle Phospholipid Metabolism Regulates Insulin Sensitivity and Contractile Function. Diabetes: A J. Am. Diabetes Assoc. 2015, 65, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Funai, K.; Song, H.; Yin, L.; Lodhi, I.J.; Wei, X.; Yoshino, J.; Coleman, T.; Semenkovich, C.F. Muscle lipogenesis balances insulin sensitivity and strength through calcium signaling. J. Clin. Investig. 2013, 123, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Selathurai, A.; Kowalski, G.M.; Burch, M.L.; Sepulveda, P.; Risis, S.; Lee-Young, R.S.; Lamon, S.; Meikle, P.J.; Genders, A.J.; McGee, S.L. The CDP-ethanolamine pathway regulates skeletal muscle diacylglycerol content and mitochondrial biogenesis without altering insulin sensitivity. Cell Metab. 2015, 21, 718–730. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Wu, Z.; Wang, X.; Yang, P.; Yu, D.; Zhao, C.; Xu, G.; Kang, L. Metabolomic analysis reveals that carnitines are key regulatory metabolites in phase transition of the locusts. Proc. Natl. Acad. Sci. USA 2012, 109, 3259–3263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Q.-L.; Meng, X.; Fu, X.; Chen, B.; Yang, J.; Yang, H.; Zhou, Q. Intermediate metabolites of the pyrimidine metabolism pathway extend the lifespan of C. elegans through regulating reproductive signals. Aging (Albany NY) 2019, 11, 3993–4010. [Google Scholar] [CrossRef] [PubMed]
- Che, X.; Liu, P.; Wu, C.; Song, W.; An, N.; Yu, L.; Bai, Y.; Xing, Z.; Cai, J.; Wang, X.; et al. Potential role of the ecto-5’-nucleotidase in morphine-induced uridine release and neurobehavioral changes. Neuropharmacology 2018, 141, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Groenendijk, M.; De Wilde, M.C.; Hageman, R.J.J.; Kamphuis, P.J.G.; Hendrikus, K.P.J. Use of a Composition for Supporting Activities in Daily Living and Prolonging Independent Living, Comprises Docosahexaenoic Acid and/or Eicosapentaenoic Acid, and Uridine. WO 2009/002146 Al, 31 December 2008. [Google Scholar]
- Honda, K.; Komoda, Y.; Nishida, S.; Nagasaki, H.; Higashi, A.; Uchizono, K.; Inoue, S. Uridine as an active component of sleep-promoting substance: Its effects on nocturnal sleep in rats. Neurosci. Res. 1984, 1, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.S.; Fisher, H.; Wagner, G.C. Uridine reduces rotation induced by l-dopa and methamphetamine in 6-ohda-treated rats. Pharmacol. Biochem. Behav. 1995, 52, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wu, C.; Song, W.; Yu, L.; Yang, X.; Xiang, R.; Wang, F.; Yang, J. Uridine decreases morphine-induced behavioral sensitization by decreasing dorsal striatal dopamine release possibly via agonistic effects at GABA(A) receptors. Eur. Neuropsychopharmacol. 2014, 24, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.-H.; Coloff, J.L. The Diverse Functions of Non-Essential Amino Acids in Cancer. Cancers 2019, 11, 675. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P. The glycinergic inhibitory synapse. Cell. Mol. Life Sci. 2001, 58, 760–793. [Google Scholar] [CrossRef]
- Vinay, L.; Jean-Xavier, C. Plasticity of spinal cord locomotor networks and contribution of cation-chloride cotransporters. Brain Res. Rev. 2008, 57, 103–110. [Google Scholar] [CrossRef]
- Hinckley, C.; Seebach, B.; Ziskind-Conhaim, L. Distinct roles of glycinergic and GABAergic inhibition in coordinating locomotor-like rhythms in the neonatal mouse spinal cord. Neuroscience 2005, 131, 745–758. [Google Scholar] [CrossRef]
- Pflieger, J.; Clarac, F.; Vinay, L. Effects of bicuculline on spinal networks and motorneurons in the neonatal rats. Eur. J. Neurosci. 2000, 12, 154. [Google Scholar]
- Kudo, N.; Nishimaru, H. Reorganization of locomotor activity during development in the prenatal rat. Ann. N. Y. Acad. Sci. 2006, 860, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Fidelin, K.; Wyart, C. Inhibition and motor control in the developing zebrafish spinal cord. Curr. Opin. Neurobiol. 2014, 26, 103–109. [Google Scholar] [CrossRef] [PubMed]
- De Koning, T.J.; Snell, K.; Duran, M.; Berger, R.; Poll-the, B.-T.; Surtees, R. l-Serine in disease and development. Biochem. J. 2003, 371, 653–661. [Google Scholar] [CrossRef]
- Ito, Y.; Takahashi, S.; Shen, M.; Yamaguchi, K.; Satoh, M. Effects of L-serine ingestion on human sleep. SpringerPlus 2014, 3, 456. [Google Scholar] [CrossRef] [Green Version]
- Nagamachi, S.; Nishigawa, T.; Takakura, M.; Ikeda, H.; Kodaira, M.; Yamaguchi, T.; Chowdhury, V.S.; Yasuo, S.; Furuse, M. Dietary L-serine modifies free amino acid composition of maternal milk and lowers the body weight of the offspring in mice. J. Vet. Med. Sci. 2017, 80, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Shigemi, K.; Tsuneyoshi, Y.; Yamada, S.; Kabuki, Y.; Furuse, M. Oral administration of L-serine reduces the locomotor activity of socially isolated rats. Neurosci. Lett. 2010, 468, 75–79. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Rivera, L.V.; Goto, J.J. A locomotor activity study of the effects of l-serine to neurodegeneration impacts of beta-methylamino-l-alanine in Drosoph. Melanogaster FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- D’Aniello, A.; Di Fiore, M.M.; Fisher, G.H.; Milone, A.; Seleni, A.; D’Aniello, S.; Perna, A.F.; Ingrosso, D. Occurrence of D-aspartic acid and N-methyl-D-aspartic acid in rat neuroendocrine tissues and their role in the modulation of luteinizing hormone and growth hormone release. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2000, 14, 699–714. [Google Scholar] [CrossRef]
- Roshanzamir, F.; Safavi, S.M. The putative effects of D-Aspartic acid on blood testosterone levels: A systematic review. Int. J. Reprod. Biomed. 2017, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Di Fiore, M.M.; Boni, R.; Santillo, A.; Falvo, S.; Gallo, A.; Esposito, S.; Baccari, G.C. D-Aspartic acid in vertebrate reproduction: Animal models and experimental designs. Biomolecules 2019, 9, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations | Control Group | AjK1 Treatment Groups | AjK2 Treatment Groups | ||||
|---|---|---|---|---|---|---|---|
| Seawater | 0.1 μM | 1 μM | 10 μM | 0.1 μM | 1 μM | 10 μM | |
| Groups | S | K1L | K1M | K1H | K2L | K2M | K2H |
| Moving Parameters | Kinetic Constants | S | AjK1 | AjK2 | ||||
|---|---|---|---|---|---|---|---|---|
| H | M | L | H | M | L | |||
| Distance | Maximum reduction (cm) | n.a. | 328 ± 34 | 302 ± 101 | n.a. | n.a. | n.a. | n.a. |
| Decay rate constant (h-1) | n.a. | 0.26 ± 0.04 | 0.58 ± 0.26 | n.a. | n.a. | n.a. | n.a. | |
| Half-life period (h) | n.a. | 2.68 ± 0.40 | 1.20 ± 0.55 | n.a. | n.a. | n.a. | n.a. | |
| Moving duration | Maximum reduction (min) | n.a. | 43.5 ± 6.0 | 46.8 ± 16.3 | n.a. | n.a. | n.a. | n.a. |
| Decay rate constant (h-1) | n.a. | 0.24 ± 0.05 | 0.61 ± 0.29 | n.a. | n.a. | n.a. | n.a. | |
| Half-life period (h) | n.a. | 2.85 ± 0.57 | 1.13 ± 0.53 | n.a. | n.a. | n.a. | n.a. | |
| Velocity | Maximum reduction (cm/min) | n.a. | 4.85 ± 0.63 | 5.07 ± 1.70 | n.a. | n.a. | n.a. | n.a. |
| Decay rate constant (h-1) | n.a. | 0.25 ± 0.05 | 0.58 ± 0.27 | n.a. | n.a. | n.a. | n.a. | |
| Half-life period (h) | n.a. | 2.83 ± 0.54 | 1.19 ± 0.55 | n.a. | n.a. | n.a. | n.a. | |
| Step number | Maximum reduction | n.a. | 49.1 ± 6.1 | - | - | n.a. | - | - |
| Decay rate constant (h-1) | n.a. | 0.24 ± 0.04 | - | - | n.a. | - | - | |
| Half-life period (h) | n.a. | 2.84 ± 0.51 | - | - | n.a. | - | - | |
| Stride | Maximum reduction (cm) | n.a. | 4.45 ± 1.06 | - | - | 2.57 ± 1.02 | - | - |
| Decay rate constant (h-1) | n.a. | 0.28 ± 0.10 | - | - | 0.75 ± 0.40 | - | - | |
| Half-life period (h) | n.a. | 2.46 ± 0.84 | - | - | 0.92 ± 0.49 | - | - | |
| Step frequency | Maximum reduction (step/min) | n.a. | n.a. | - | - | n.a. | - | - |
| Decay rate constant (h-1) | n.a. | n.a. | - | - | n.a. | - | - | |
| Half-life period (h) | n.a. | n.a. | - | - | n.a. | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Zhang, L.; Xiao, K. Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicus. Animals 2023, 13, 705. https://doi.org/10.3390/ani13040705
Guo X, Zhang L, Xiao K. Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicus. Animals. 2023; 13(4):705. https://doi.org/10.3390/ani13040705
Chicago/Turabian StyleGuo, Xueying, Libin Zhang, and Kang Xiao. 2023. "Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicus" Animals 13, no. 4: 705. https://doi.org/10.3390/ani13040705
APA StyleGuo, X., Zhang, L., & Xiao, K. (2023). Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicus. Animals, 13(4), 705. https://doi.org/10.3390/ani13040705