
Formation, Application, and Significance of Chicken Primordial Germ Cells: A Review
 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. An Overview of Avian PGCs

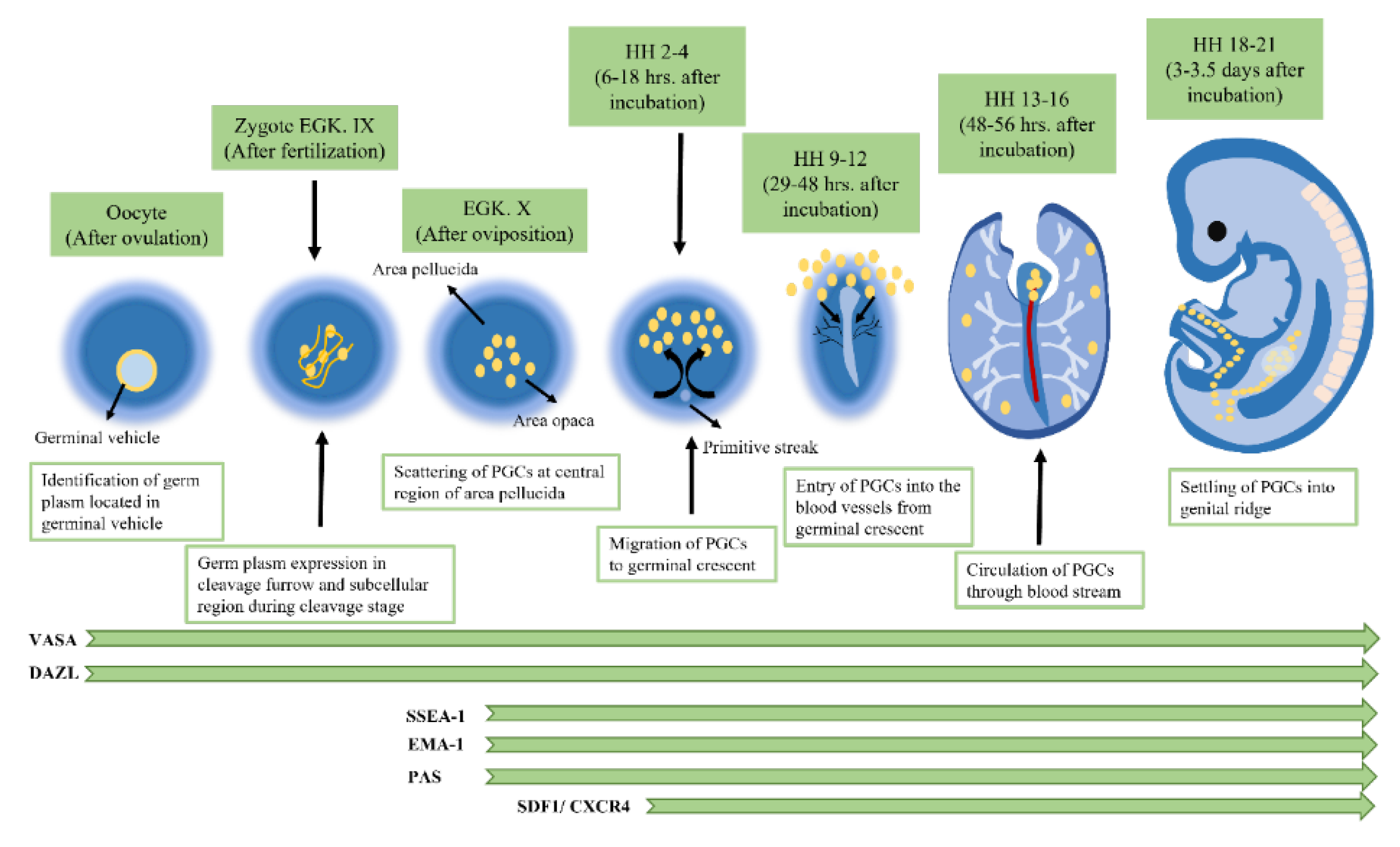
1.1.1. Origin of Chicken PGCs
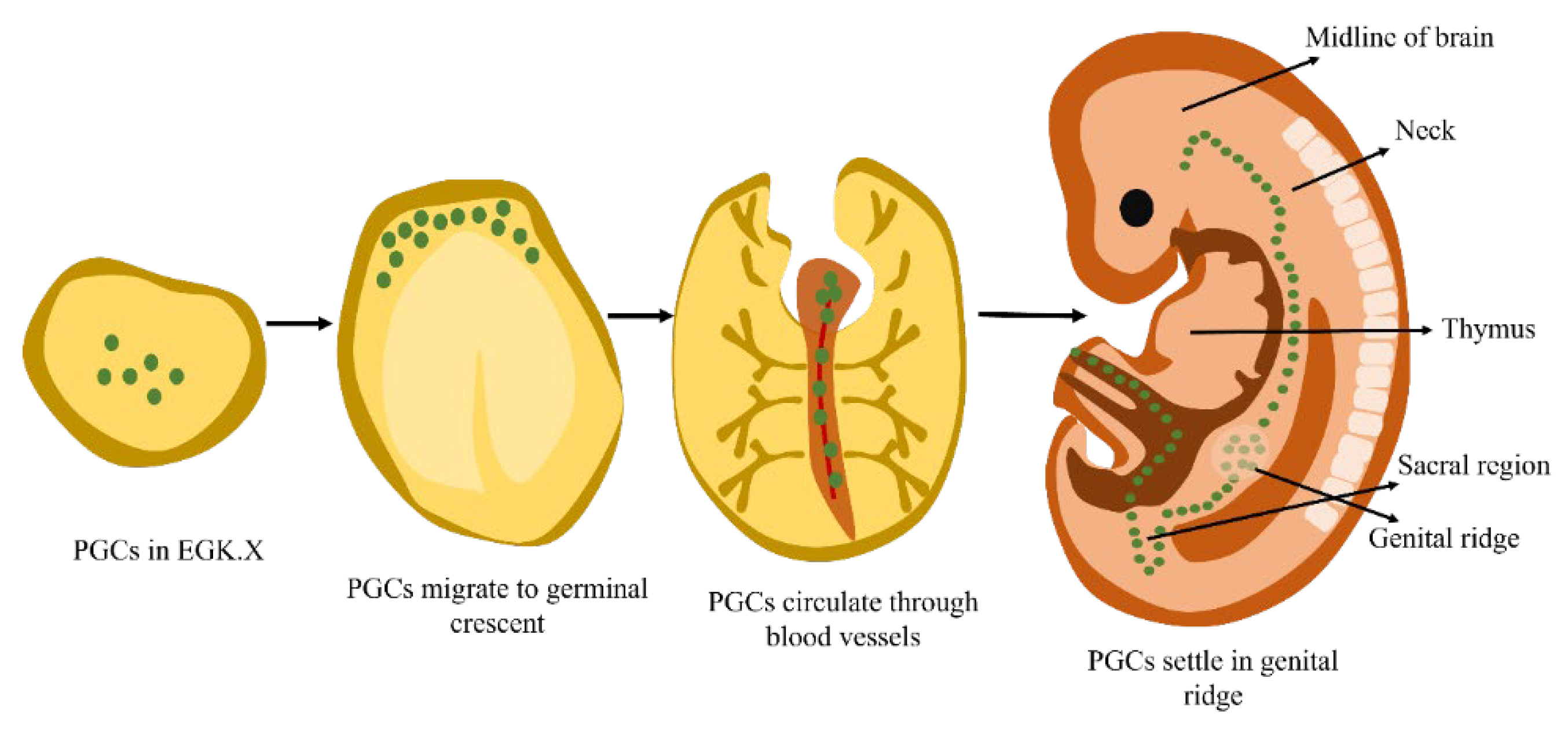
1.1.2. Migration of PGCs

1.2. Research Progress on PGC Gene Markers and Identification Methods
1.2.1. Derivation of PGCs Using an Indirect Co-Culture Method
1.2.2. Identification of PGCs
1.3. Research Progress on In Vitro Culture Line Formation and Differentiation in PGCs
Propagation of Cultured Primordial Germ Cells In Vitro
1.4. Chicken Cell Line Formation
1.4.1. Epigenesis of PGCs
1.4.2. Germ Cell Gametogenesis in Chicken
2. The Application of PGCs
2.1. Cryopreservation of PGCs—A Frozen Storage Technique
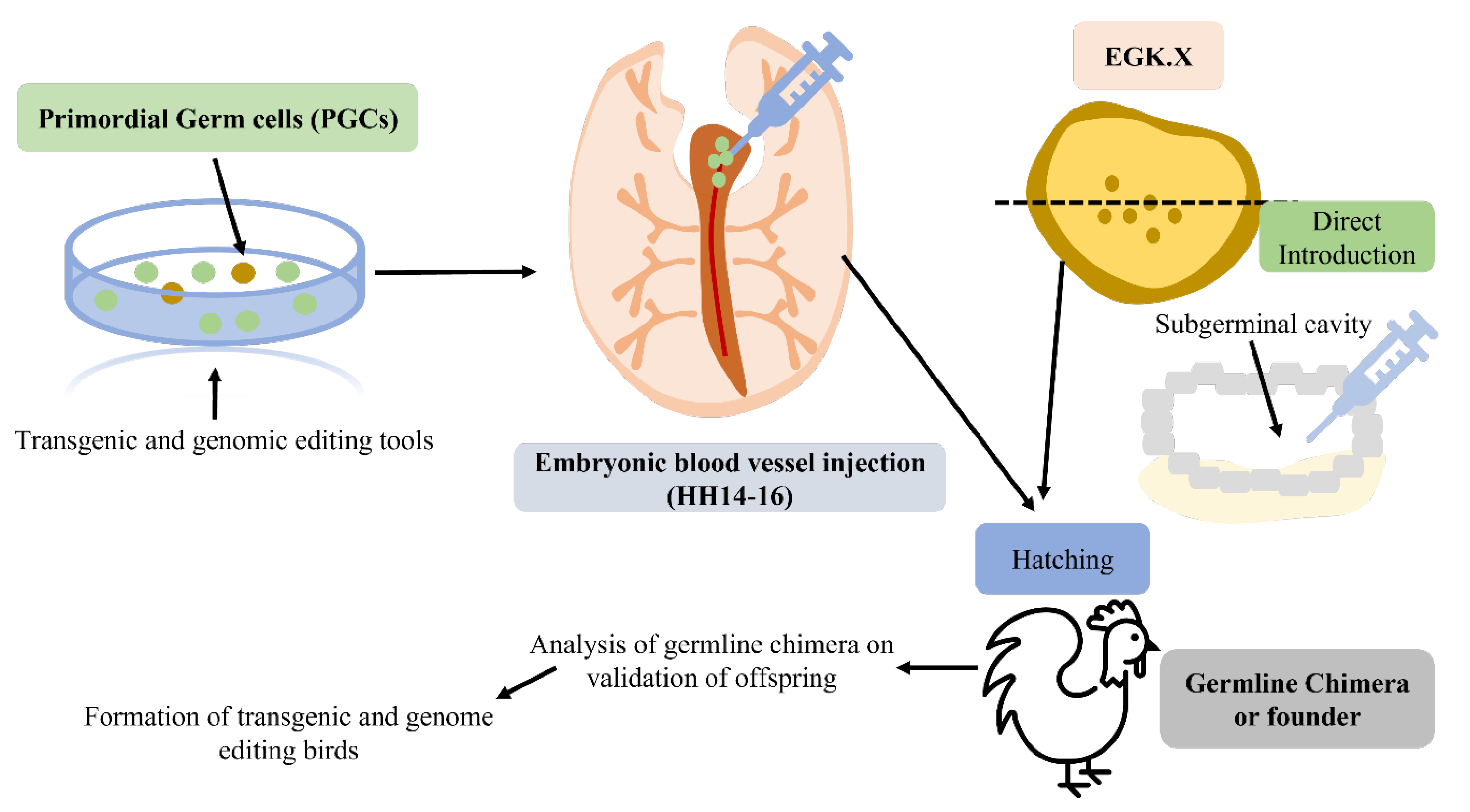
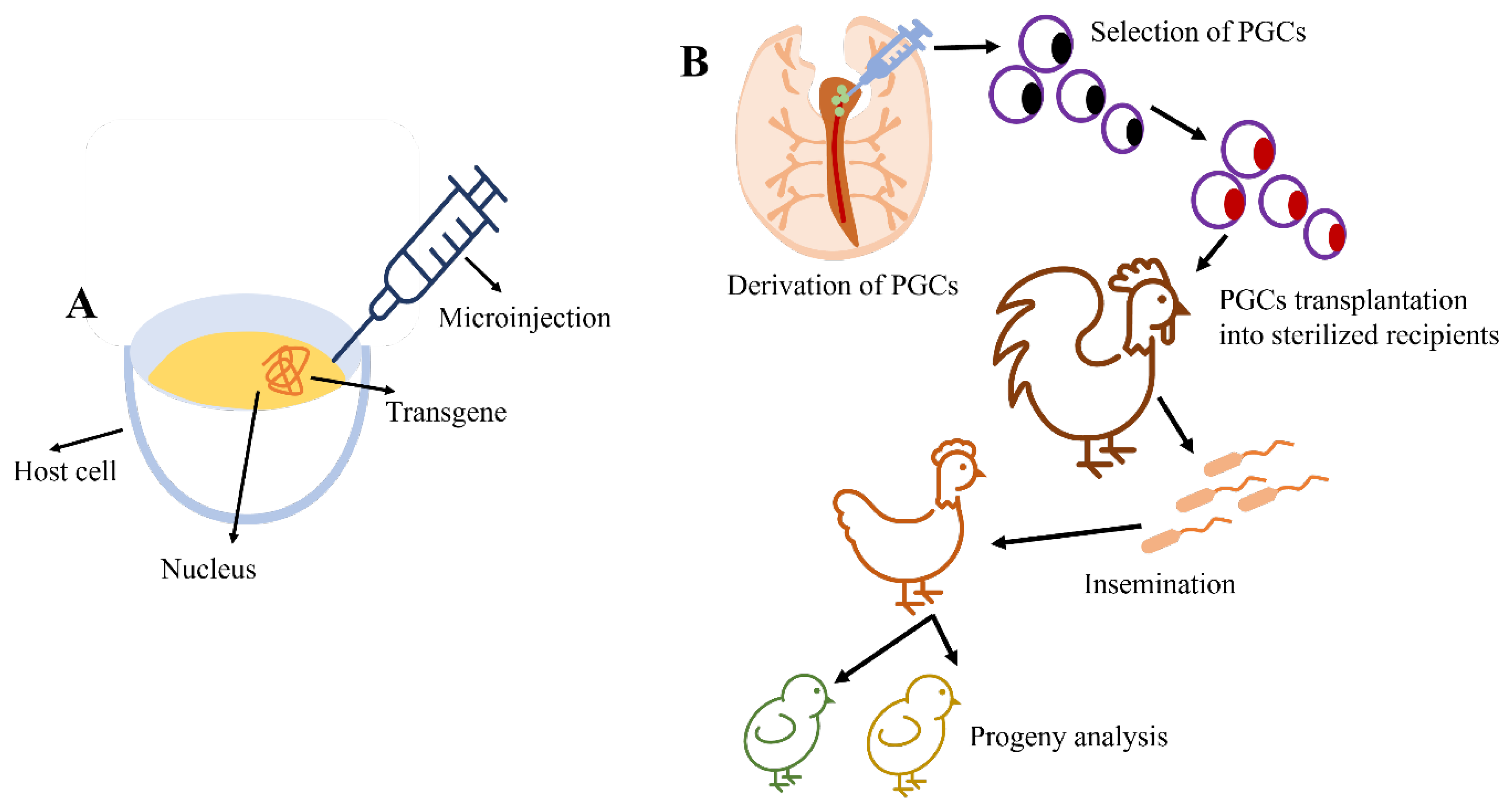
2.2. Research Progress of Primordial Germ Cells (PGCs)—A Cell-Based Gene Transfer Method
3. Future Aspects and Considerations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Himmel, L.E.; Wilson, K.L.; Santagostino, S.F.; Bolon, B. Chapter 23—Genetically Engineered Animal Models in Toxicologic Research. In Haschek and Rousseaux’s Handbook of Toxicologic Pathology, 4th ed.; Haschek, W.M., Rousseaux, C.G., Wallig, M.A., Bolon, B., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 859–924. [Google Scholar] [CrossRef]
- Doran, T.J.; Cooper, C.A.; Jenkins, K.A.; Tizard, M.L. Advances in genetic engineering of the avian genome: “Realising the promise”. Transgenic Res. 2016, 25, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Tagami, T.; Miyahara, D.; Nakamura, Y. Avian Primordial Germ Cells. Adv. Exp. Med. Biol. 2017, 1001, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Lu, Z.; Chen, D.; Yang, M.; Liao, Y.; Mao, W.; Mo, L.; Sun, J.; Yang, W.; Xu, H.; et al. Derivation of chicken primordial germ cells using an indirect Co-culture system. Theriogenology 2019, 123, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.E.; Lehmann, R. Mechanisms guiding primordial germ cell migration: Strategies from different organisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Wang, S.; Jiang, H.; Hua, Y.; Yin, B.; Huang, X.; Man, Q.; Wang, H.; Zhu, G. Oct4 dependent chromatin activation is required for chicken primordial germ cell migration. Stem Cell Rev. Rep. 2022, 18, 2535–2546. [Google Scholar] [CrossRef]
- Zuo, Q.; Jing, J.; Zhou, J.; Zhang, Y.; Wei, W.; Chen, G.; Li, B. Dual regulatory actions of LncBMP4 on BMP4 promote chicken primordial germ cell formation. EMBO Rep. 2022, 23, e52491. [Google Scholar] [CrossRef]
- Andersson, L.; Georges, M. Domestic-animal genomics: Deciphering the genetics of complex traits. Nat. Rev. Genet. 2004, 5, 202–212. [Google Scholar] [CrossRef]
- Han, J.Y.; Park, Y.H. Primordial germ cell-mediated transgenesis and genome editing in birds. J. Anim. Sci. Biotechnol. 2018, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Lee, H.C.; Han, J.Y. Germline Modification and Engineering in Avian Species. Mol. Cells 2015, 38, 743–749. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.W.; Scangos, G.A.; Plotkin, D.J.; Barbosa, J.A.; Ruddle, F.H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 7380–7384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, R.E.; Pursel, V.G.; Rexroad, C.E., Jr.; Wall, R.J.; Bolt, D.J.; Ebert, K.M.; Palmiter, R.D.; Brinster, R.L. Production of transgenic rabbits, sheep and pigs by microinjection. Nature 1985, 315, 680–683. [Google Scholar] [CrossRef] [Green Version]
- Thomas, K.R.; Capecchi, M.R. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 1987, 51, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y. Germ cells and transgenesis in chickens. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Choi, H.J.; Park, T.S.; Lee, S.I.; Kim, Y.M.; Rengaraj, D.; Nagai, H.; Sheng, G.; Lim, J.M.; Han, J.Y. Cleavage events and sperm dynamics in chick intrauterine embryos. PLoS ONE 2013, 8, e80631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herron, L.R.; Pridans, C.; Turnbull, M.L.; Smith, N.; Lillico, S.; Sherman, A.; Gilhooley, H.J.; Wear, M.; Kurian, D.; Papadakos, G.; et al. A chicken bioreactor for efficient production of functional cytokines. BMC Biotechnol. 2018, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Hassani, S.-N.; Mozdziak, P.; Baharvand, H. Avian embryos and related cell lines: A convenient platform for recombinant proteins and vaccine production. Biotechnol. J. 2017, 12, 1600598. [Google Scholar] [CrossRef]
- Pal, A. Transgenesis and Biopharming. In Protocols in Advanced Genomics and Allied Techniques; Springer: New York, NY, USA, 2022; pp. 483–503. [Google Scholar] [CrossRef]
- Love, J.; Gribbin, C.; Mather, C.; Sang, H. Transgenic birds by DNA microinjection. Bio/Technology 1994, 12, 60–63. [Google Scholar] [CrossRef]
- Eyal-Giladi, H.; Kochav, S. From cleavage to primitive streak formation: A complementary normal table and a new look at the first stages of the development of the chick. I. General morphology. Dev. Biol. 1976, 49, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; van de Lavoir, M.C.; Albanese, J.; Beenhouwer, D.O.; Cardarelli, P.M.; Cuison, S.; Deng, D.F.; Deshpande, S.; Diamond, J.H.; Green, L.; et al. Production of human monoclonal antibody in eggs of chimeric chickens. Nat. Biotechnol. 2005, 23, 1159–1169. [Google Scholar] [CrossRef]
- Capecchi, M.R. Gene targeting in mice: Functional analysis of the mammalian genome for the twenty-first century. Nat. Rev. Genet. 2005, 6, 507–512. [Google Scholar] [CrossRef]
- Waldeyer, W. Eierstock und Ei: Ein Beitrag zur Anatomie und Entwicklungsgeschichte des Sexualorgane; Hansebooks: Leipzig, Germany, 1870. [Google Scholar]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. Dev. Dyn. 1992, 195, 231–272. [Google Scholar] [CrossRef]
- Eyal-Giladi, H.; Ginsburg, M.; Farbarov, A. Avian primordial germ cells are of epiblastic origin. J. Embryol. Exp. Morphol. 1981, 65, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Tsang, T.E.; Khoo, P.L.; Jamieson, R.V.; Zhou, S.X.; Ang, S.L.; Behringer, R.; Tam, P.P. The allocation and differentiation of mouse primordial germ cells. Int. J. Dev. Biol. 2001, 45, 549–555. [Google Scholar]
- Tsunekawa, N.; Naito, M.; Sakai, Y.; Nishida, T.; Noce, T. Isolation of chicken vasa homolog gene and tracing the origin of primordial germ cells. Development 2000, 127, 2741–2750. [Google Scholar] [CrossRef]
- Lee, H.C.; Choi, H.J.; Lee, H.G.; Lim, J.M.; Ono, T.; Han, J.Y. DAZL Expression Explains Origin and Central Formation of Primordial Germ Cells in Chickens. Stem Cells Dev. 2016, 25, 68–79. [Google Scholar] [CrossRef]
- Mozdziak, P.E.; Petitte, J.N. Status of transgenic chicken models for developmental biology. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 229, 414–421. [Google Scholar] [CrossRef]
- Ginsburg, M.; Eyal-Giladi, H. Primordial germ cells of the young chick blastoderm originate from the central zone of the area pellucida irrespective of the embryo-forming process. Development 1987, 101, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Urven, L.E.; Erickson, C.A.; Abbott, U.K.; McCarrey, J.R. Analysis of germ line development in the chick embryo using an anti-mouse EC cell antibody. Development 1988, 103, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Nussubaum, M. Zur Differenzierung des Geschlechts im Tierreich. Arch. Mikrosk. Anat. 1880, 18, 1–121. [Google Scholar] [CrossRef] [Green Version]
- Swift, C.H. Origin and Early History of the Primordial Germ-Cells in the Chick; University of Chicago: Chicago, IL, USA, 1914. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, J.B. The history of the germ cells in the domestic fowl. J. Morphol. 1928, 46, 275–315. [Google Scholar] [CrossRef]
- Karagenç, L.; Cinnamon, Y.; Ginsburg, M.; Petitte, J.N. Origin of primordial germ cells in the prestreak chick embryo. Dev. Genet. 1996, 19, 290–301. [Google Scholar] [CrossRef]
- Motono, M.; Ohashi, T.; Nishijima, K.; Iijima, S. Analysis of chicken primordial germ cells. Cytotechnology 2008, 57, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Mozdziak, P.E.; Wysocki, R.; Angerman-Stewart, J.; Pardue, S.L.; Petitte, J.N. Production of chick germline chimeras from fluorescence-activated cell-sorted gonocytes. Poult. Sci. 2006, 85, 1764–1768. [Google Scholar] [CrossRef]
- Ancelin, K.; Lange, U.C.; Hajkova, P.; Schneider, R.; Bannister, A.J.; Kouzarides, T.; Surani, M.A. Blimp1 associates with Prmt5 and directs histone arginine methylation in mouse germ cells. Nat. Cell Biol. 2006, 8, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, P.S.; Siekhaus, D.E.; Lehmann, R. In vivo migration: A germ cell perspective. Annu. Rev. Cell Dev. Biol. 2006, 22, 237–265. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.; McGrew, M. Primordial Germ Cell Technologies for Avian Germplasm Cryopreservation and Investigating Germ Cell Development. J. Poult. Sci. 2012, 49, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.S.; Lee, H.C.; Kim, H.J.; Lee, H.G.; Kim, Y.M.; Lee, H.J.; Park, Y.H.; Yang, S.Y.; Rengaraj, D.; Park, T.S.; et al. Spatial and temporal action of chicken primordial germ cells during initial migration. Reproduction 2015, 149, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Han, J.Y. The early development of germ cells in chicken. Int. J. Dev. Biol. 2018, 62, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Molyneaux, K.A.; Zinszner, H.; Kunwar, P.S.; Schaible, K.; Stebler, J.; Sunshine, M.J.; O’Brien, W.; Raz, E.; Littman, D.; Wylie, C.; et al. The chemokine SDF1/CXCL12 and its receptor CXCR4 regulate mouse germ cell migration and survival. Development 2003, 130, 4279–4286. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Usui, F.; Miyahara, D.; Mori, T.; Ono, T.; Takeda, K.; Nirasawa, K.; Kagami, H.; Tagami, T. Efficient system for preservation and regeneration of genetic resources in chicken: Concurrent storage of primordial germ cells and live animals from early embryos of a rare indigenous fowl (Gifujidori). Reprod. Fertil. Dev. 2010, 22, 1237–1246. [Google Scholar] [CrossRef]
- van de Lavoir, M.C.; Diamond, J.H.; Leighton, P.A.; Mather-Love, C.; Heyer, B.S.; Bradshaw, R.; Kerchner, A.; Hooi, L.T.; Gessaro, T.M.; Swanberg, S.E.; et al. Germline transmission of genetically modified primordial germ cells. Nature 2006, 441, 766–769. [Google Scholar] [CrossRef]
- Kerr, C.L.; Gearhart, J.D.; Elliott, A.M.; Donovan, P.J. Embryonic germ cells: When germ cells become stem cells. Semin. Reprod. Med. 2006, 24, 304–313. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kagami, H.; Tagami, T. Development, differentiation and manipulation of chicken germ cells. Dev. Growth Differ. 2013, 55, 20–40. [Google Scholar] [CrossRef]
- Park, T.S.; Han, J.Y. Derivation and characterization of pluripotent embryonic germ cells in chicken. Mol. Reprod. Dev. 2000, 56, 475–482. [Google Scholar] [CrossRef]
- Park, T.S.; Jeong, D.K.; Kim, J.N.; Song, G.H.; Hong, Y.H.; Lim, J.M.; Han, J.Y. Improved germline transmission in chicken chimeras produced by transplantation of gonadal primordial germ cells into recipient embryos. Biol. Reprod. 2003, 68, 1657–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.J.; Seo, H.W.; Lee, B.R.; Yoo, M.; Womack, J.E.; Han, J.Y. Gene expression and DNA methylation status of chicken primordial germ cells. Mol. Biotechnol. 2013, 54, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Lee, B.R.; Hwang, Y.S.; Lee, H.C.; Rengaraj, D.; Song, G.; Park, T.S.; Han, J.Y. MicroRNA-mediated posttranscriptional regulation is required for maintaining undifferentiated properties of blastoderm and primordial germ cells in chickens. Proc. Natl. Acad. Sci. USA 2011, 108, 10426–10431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Extavour, C.G.; Akam, M. Mechanisms of germ cell specification across the metazoans: Epigenesis and preformation. Development 2003, 130, 5869–5884. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Yamamoto, Y.; Usui, F.; Mushika, T.; Ono, T.; Setioko, A.R.; Takeda, K.; Nirasawa, K.; Kagami, H.; Tagami, T. Migration and proliferation of primordial germ cells in the early chicken embryo. Poult. Sci. 2007, 86, 2182–2193. [Google Scholar] [CrossRef]
- Stebler, J.; Spieler, D.; Slanchev, K.; Molyneaux, K.A.; Richter, U.; Cojocaru, V.; Tarabykin, V.; Wylie, C.; Kessel, M.; Raz, E. Primordial germ cell migration in the chick and mouse embryo: The role of the chemokine SDF-1/CXCL12. Dev. Biol. 2004, 272, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Ukeshima, A. Germ cell death in the degenerating right ovary of the chick embryo. Zool. Sci. 1996, 13, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D.; Lee, J.H.; Song, S.; Kim, S.W.; Han, J.S.; Shin, S.P.; Park, B.C.; Park, T.S. Generation of myostatin-knockout chickens mediated by D10A-Cas9 nickase. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 5688–5696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koslová, A.; Trefil, P.; Mucksová, J.; Reinišová, M.; Plachý, J.; Kalina, J.; Kučerová, D.; Geryk, J.; Krchlíková, V.; Lejčková, B.; et al. Precise CRISPR/Cas9 editing of the NHE1 gene renders chickens resistant to the J subgroup of avian leukosis virus. Proc. Natl. Acad. Sci. USA 2020, 117, 2108–2112. [Google Scholar] [CrossRef]
- Park, T.S.; Park, J.; Lee, J.H.; Park, J.W.; Park, B.C. Disruption of G(0)/G(1) switch gene 2 (G0S2) reduced abdominal fat deposition and altered fatty acid composition in chicken. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Yoon, J.W.; Jung, K.M.; Kim, Y.M.; Park, J.S.; Lee, K.Y.; Park, K.J.; Hwang, Y.S.; Park, Y.H.; Rengaraj, D.; et al. Targeted gene insertion into Z chromosome of chicken primordial germ cells for avian sexing model development. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 8519–8529. [Google Scholar] [CrossRef]
- Tonus, C.; Connan, D.; Waroux, O.; Vandenhove, B.; Wayet, J.; Gillet, L.; Desmecht, D.; Antoine, N.; Ectors, F.J.; Grobet, L. Cryopreservation of chicken primordial germ cells by vitrification and slow freezing: A comparative study. Theriogenology 2017, 88, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Lavitrano, M.; Forni, M.; Bacci, M.L.; Di Stefano, C.; Varzi, V.; Wang, H.; Seren, E. Sperm mediated gene transfer in pig: Selection of donor boars and optimization of DNA uptake. Mol. Reprod. Dev. 2003, 64, 284–291. [Google Scholar] [CrossRef]
- Bahrami, S.; Amiri-Yekta, A.; Daneshipour, A.; Jazayeri, S.H.; Mozdziak, P.E.; Sanati, M.H.; Gourabi, H. Designing A Transgenic Chicken: Applying New Approaches toward A Promising Bioreactor. Cell J. 2020, 22, 133–139. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
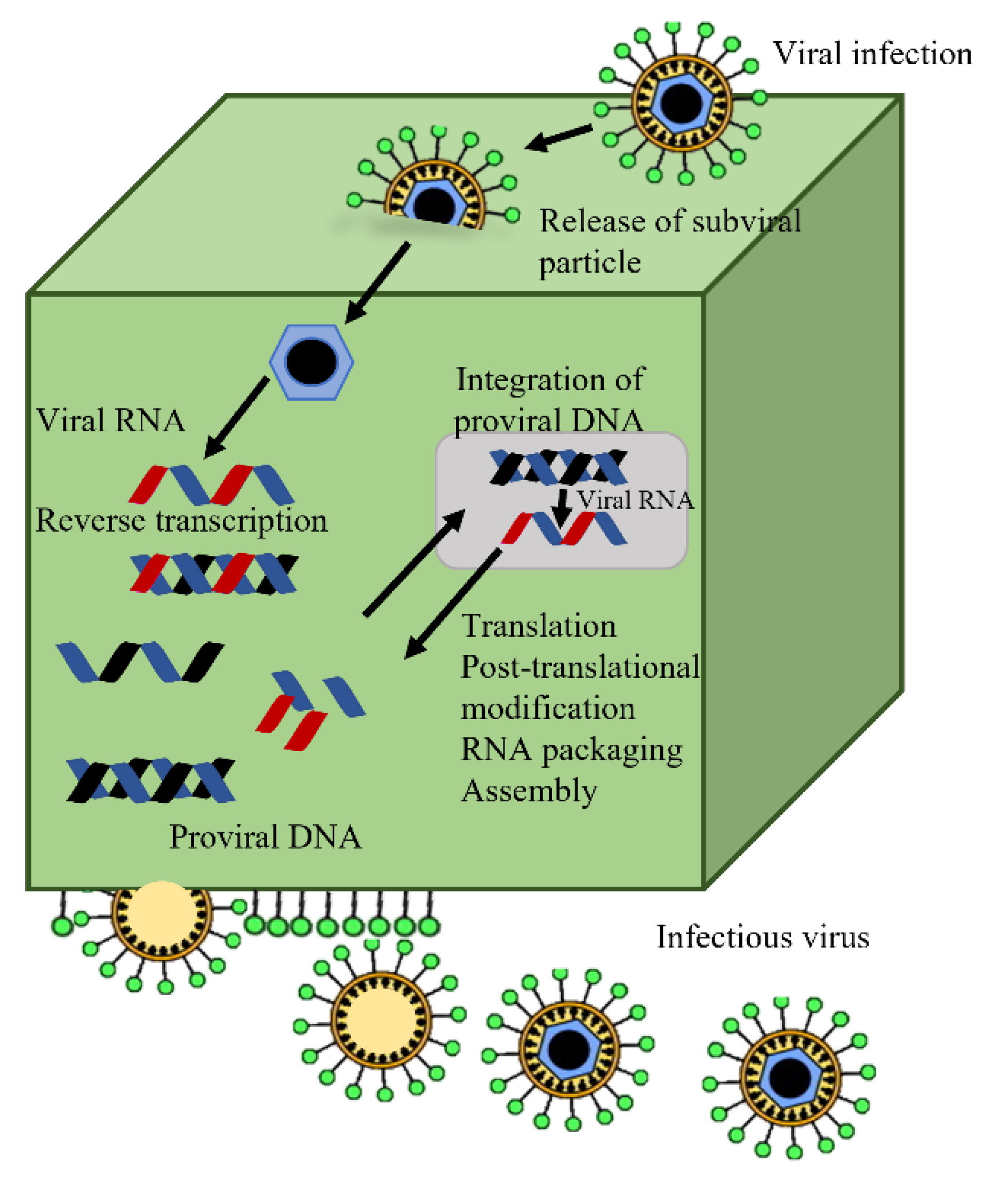{kind=link}
| Year of Study | Name of Study | References |
|---|---|---|
| 1870 | Eirstock und Ei. Eine Beitrag zur Anatomie und Entwicklungsgeschichte der Sexualorgane. | [20] |
| 1880 | Zur Differenzierung des Geschlechts im Tierreich | [32] |
| 1914 | Origin and early history of the primordial germ-cells in the chick | [33] |
| 1928 | The history of germ cells in the domestic fowl | [34] |
| 1996 | Origin of primordial germ cells in the prestreak chick embryos | [35] |
| 2006, 2008 |
| [36,37] |
| 2006 |
| [38,39] |
| Species | Mediator | Main Research | Purpose | References |
|---|---|---|---|---|
| Chicken | PGCs | A technical platform mediated by Cas9n was applied to the PGCs, and via the germline transmission system, the myostatin-knockout chickens were generated. | This mutation helps to achieve a hypermuscular phenotype in chickens. This will help promote the growth factor and lead to double-muscle development—a major economic trait in the livestock industry | [56] |
| Chicken | PGCs | In this study, an amino acid was simply deleted from the gene encoding to infect the chicken cells. The deleted amino acid acts as a receptor for avian leukosis virus subgroup J | The generated mutation is beneficial to produce resistance to avian leukosis virus subgroup J. This virus is an important pathogen in the poultry industry | [57] |
| Chicken | PGCs | PGC-mediated germline transmission was performed using the CRISPR/Cas9 tool to knockout G0/G1 and switch it by Gene 2 | This mutation helps to drastically reduce the abdominal fat in chickens, thereby improving their body fat ratio without altering their other traits | [58] |
| Chicken | PGCs | GFP was knocked into the z chromosome through the CRISPR/Cas9 tool. | GFP expressing progenies for sexing. Sexing model development was performed; this can help to detect sex during embryogenesis | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathan; Zaib, G.; Jin, K.; Zuo, Q.; Habib, M.; Zhang, Y.; Li, B. Formation, Application, and Significance of Chicken Primordial Germ Cells: A Review. Animals 2023, 13, 1096. https://doi.org/10.3390/ani13061096
Mathan, Zaib G, Jin K, Zuo Q, Habib M, Zhang Y, Li B. Formation, Application, and Significance of Chicken Primordial Germ Cells: A Review. Animals. 2023; 13(6):1096. https://doi.org/10.3390/ani13061096
Chicago/Turabian StyleMathan, Gul Zaib, Kai Jin, Qisheng Zuo, Maham Habib, Yani Zhang, and Bichun Li. 2023. "Formation, Application, and Significance of Chicken Primordial Germ Cells: A Review" Animals 13, no. 6: 1096. https://doi.org/10.3390/ani13061096
APA StyleMathan, Zaib, G., Jin, K., Zuo, Q., Habib, M., Zhang, Y., & Li, B. (2023). Formation, Application, and Significance of Chicken Primordial Germ Cells: A Review. Animals, 13(6), 1096. https://doi.org/10.3390/ani13061096