


Immunogenicity and Efficacy of a Feed-Based Bivalent Vaccine against Streptococcosis and Motile Aeromonad Septicemia in Red Hybrid Tilapia (Oreochromis sp.)
 ,
,  ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Red Hybrid Tilapia (Oreochromis sp.)
2.2. Bacterial Pathogens and Culture Conditions
2.3. Feed-Based Bivalent Vaccine Preparation
2.4. Feed Quality Analyses
2.4.1. Proximate Analysis
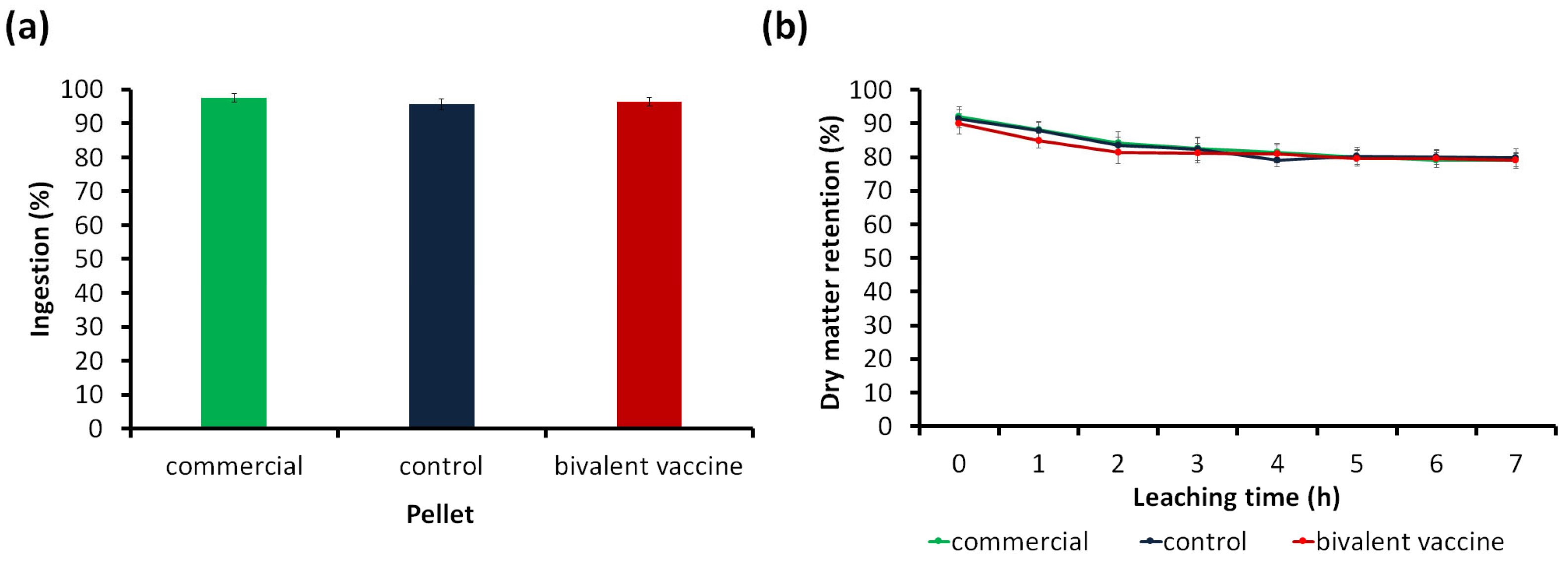
2.4.2. Palatability Test
2.4.3. Physical Stability in Water
2.4.4. Vaccine Safety Test
2.4.5. Growth Performance
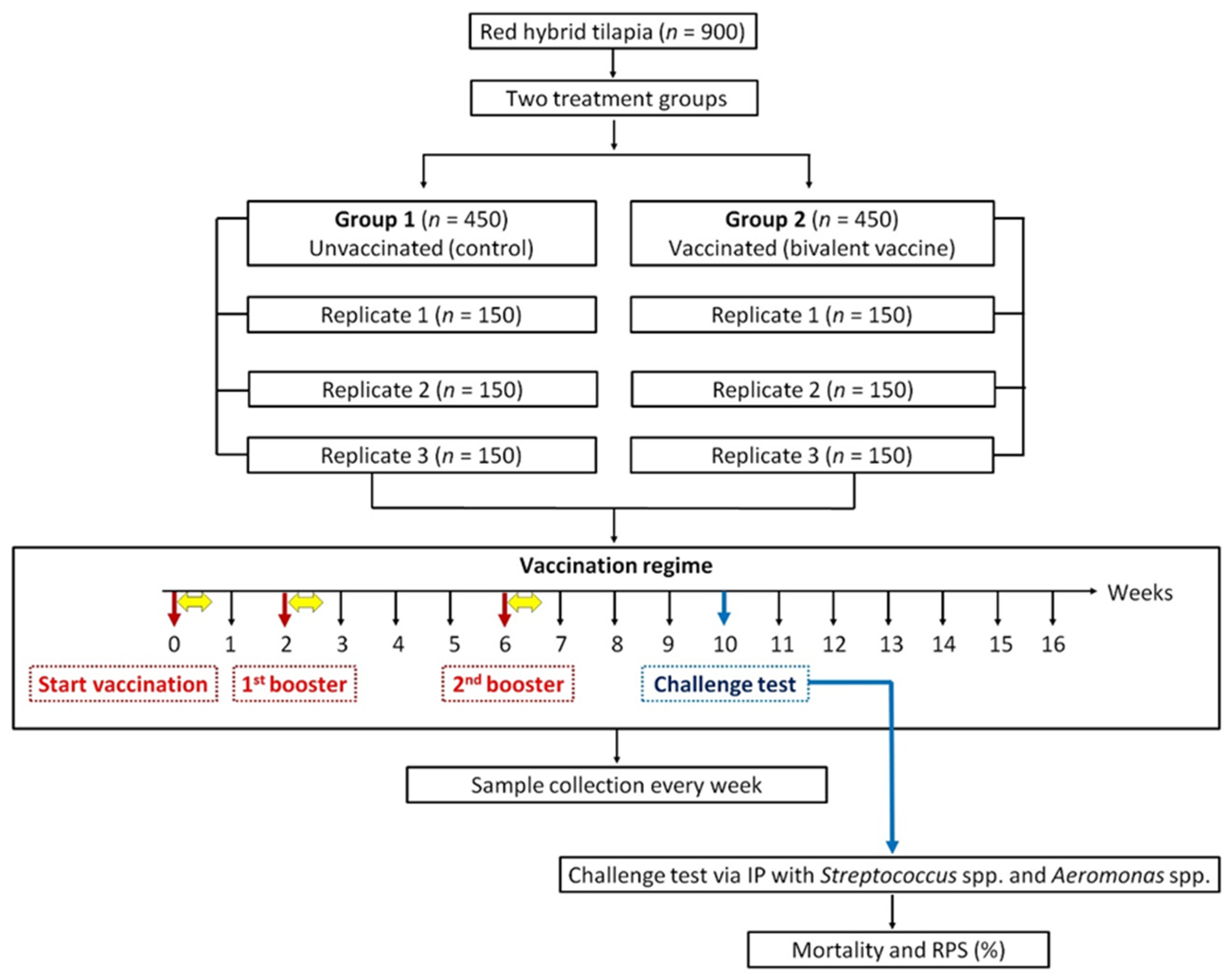
2.5. Vaccination Trial
Experimental Design
2.6. Immunological Analyses
2.6.1. Sample Collection
2.6.2. Lysozyme Assay
2.6.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6.4. Challenge Trial
2.7. Histopathological Assessment
2.8. Statistical Analysis
3. Results
3.1. Feed Quality Analyses
3.1.1. Proximate Analysis
3.1.2. Palatability Test
3.1.3. Physical Stability in Static Water Test
3.1.4. Growth Performance
3.2. Intraperitoneal Challenge Test
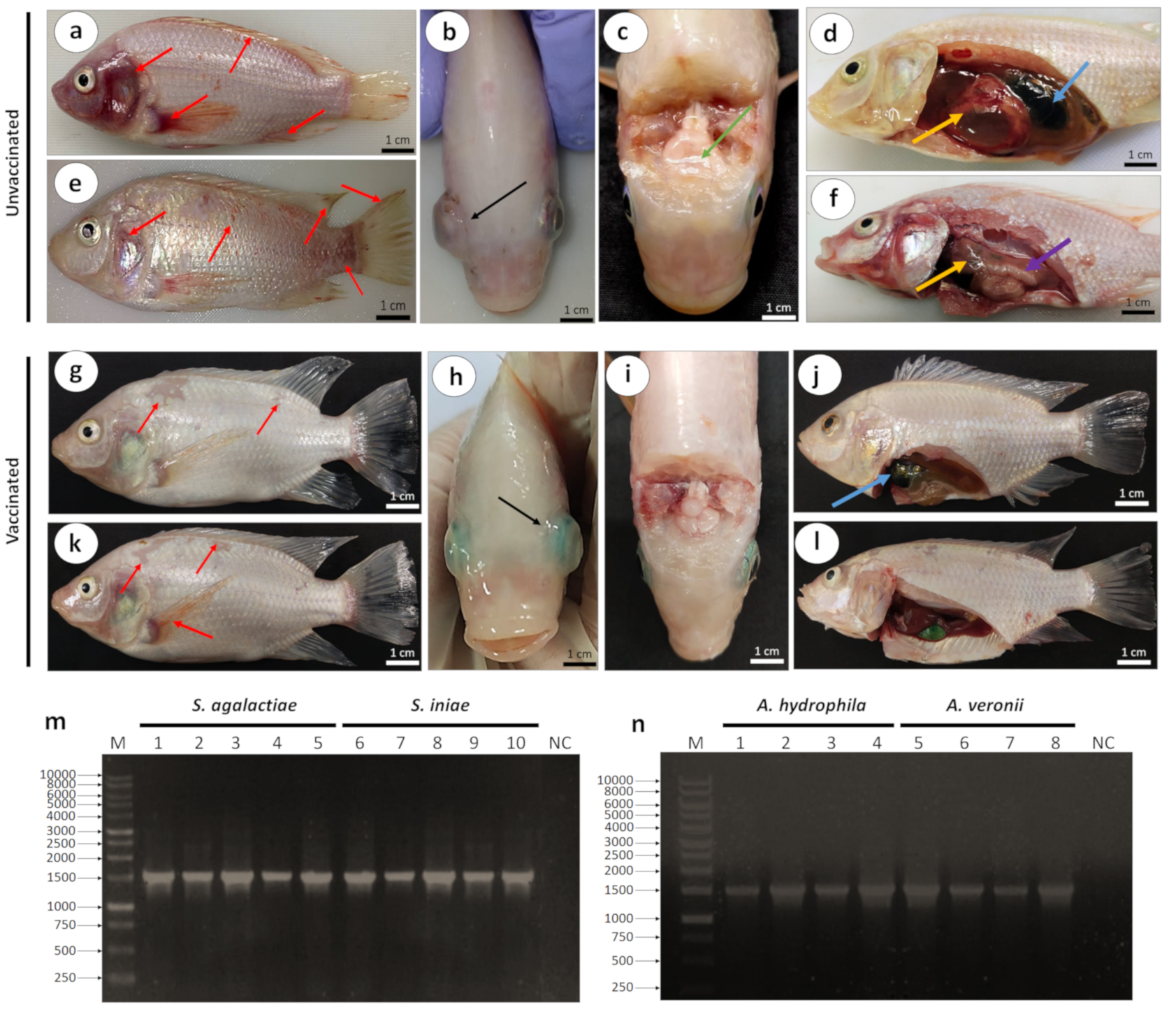
3.2.1. Clinical Signs, Gross Lesions, and Bacteriology
3.2.2. Protective Efficacies of the Feed-Based Bivalent Vaccine
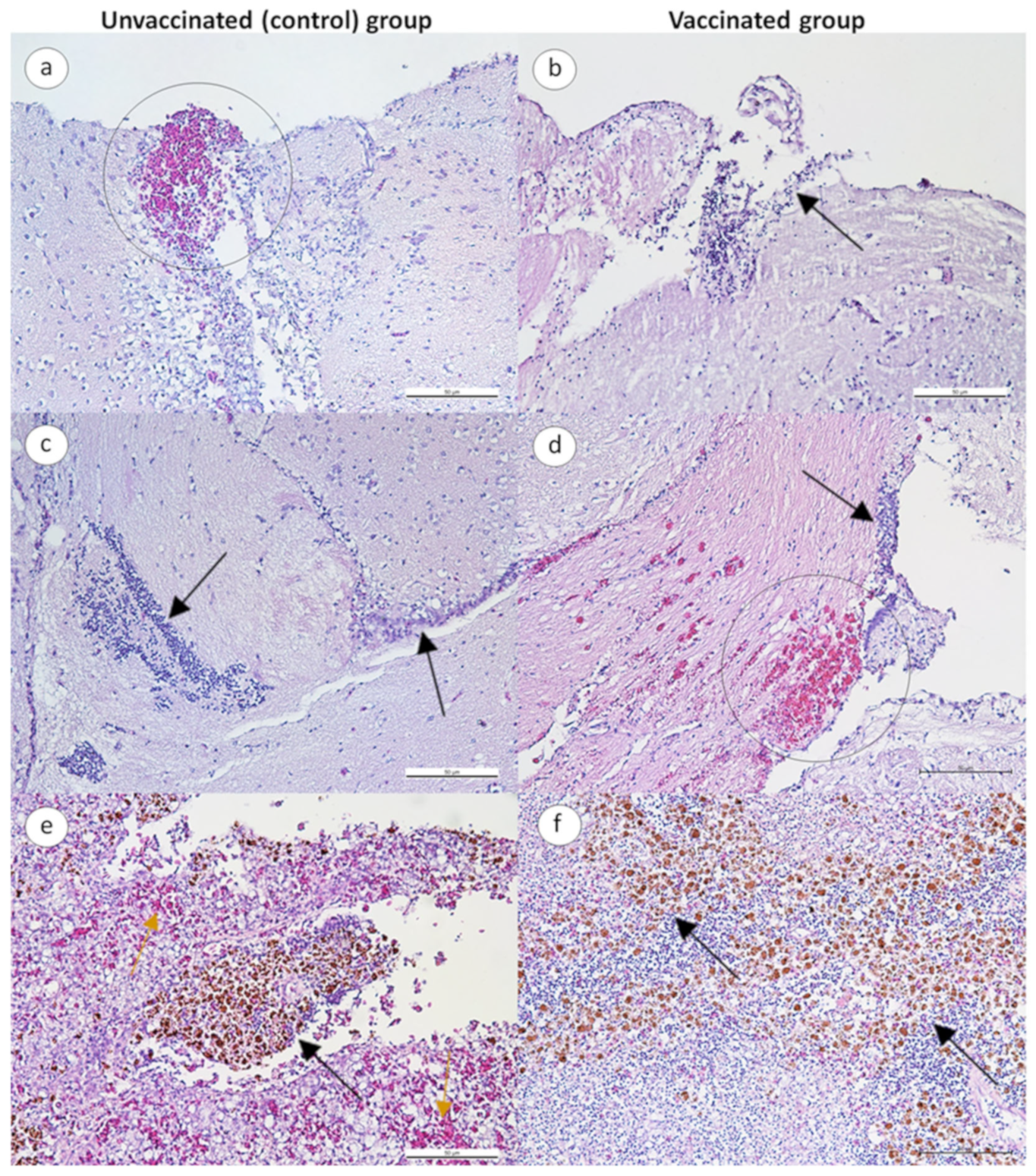
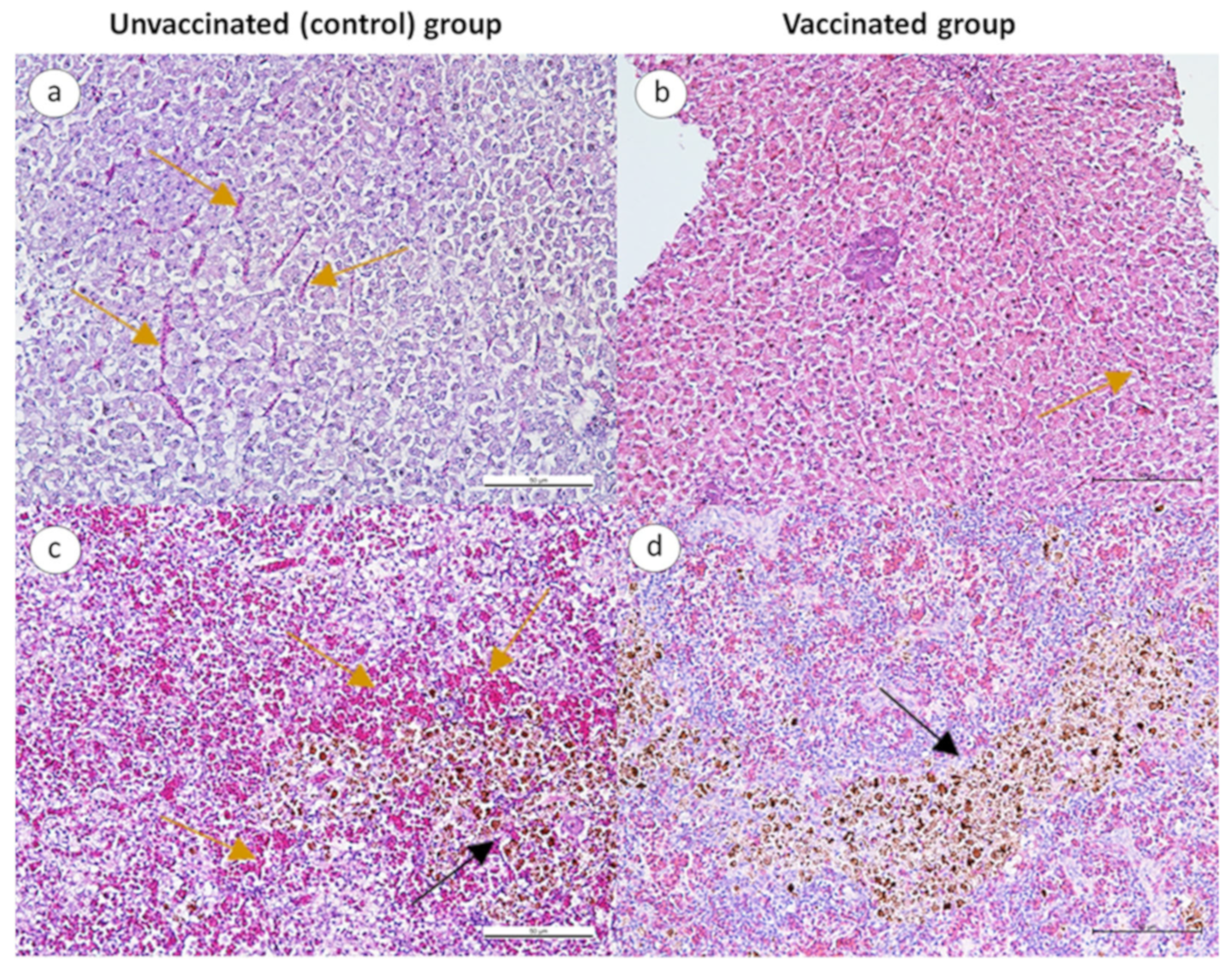
3.2.3. Histopathological Changes in Infected Fish
3.3. Immunological Responses
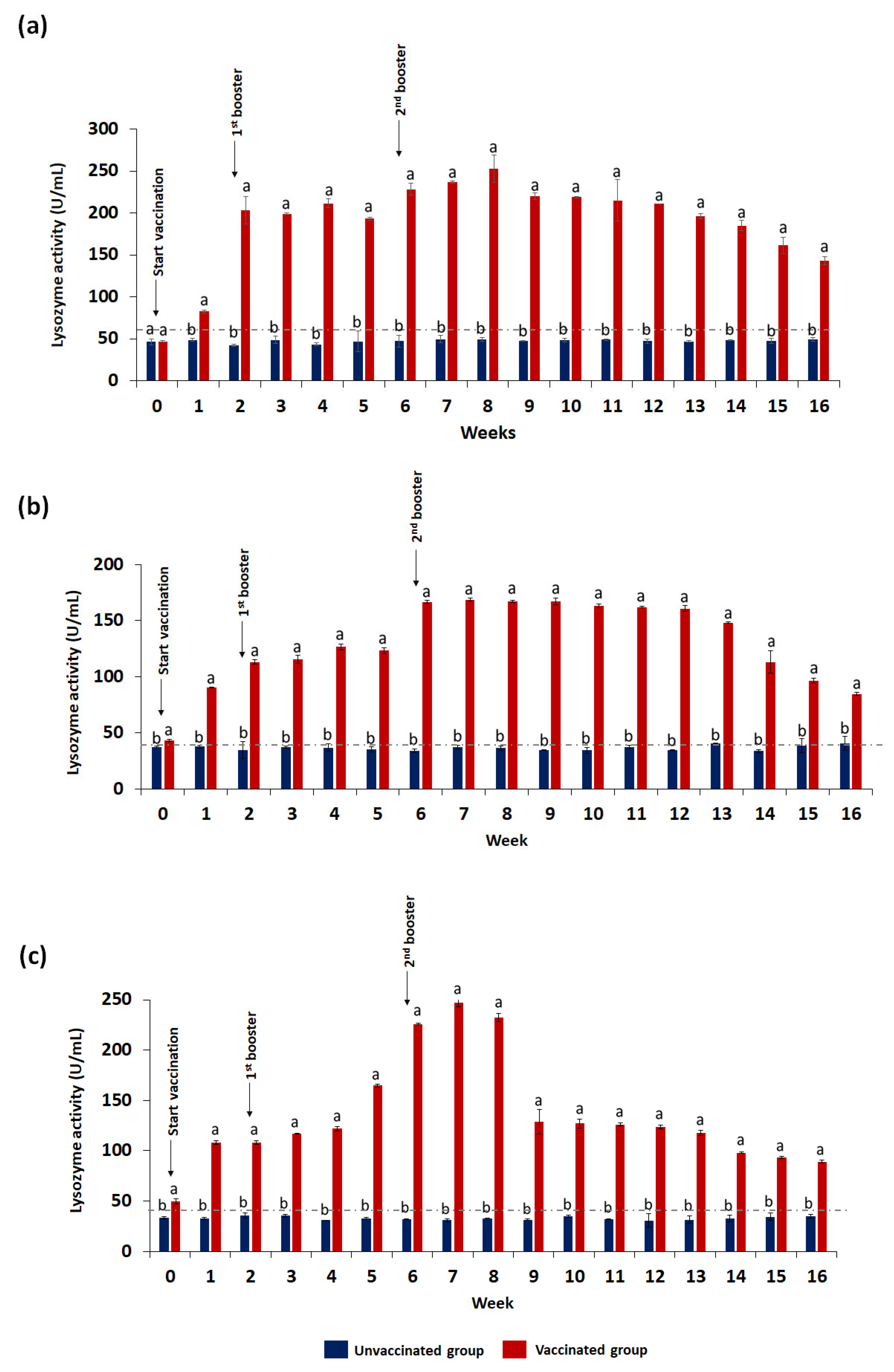
3.3.1. Lysozyme Activity
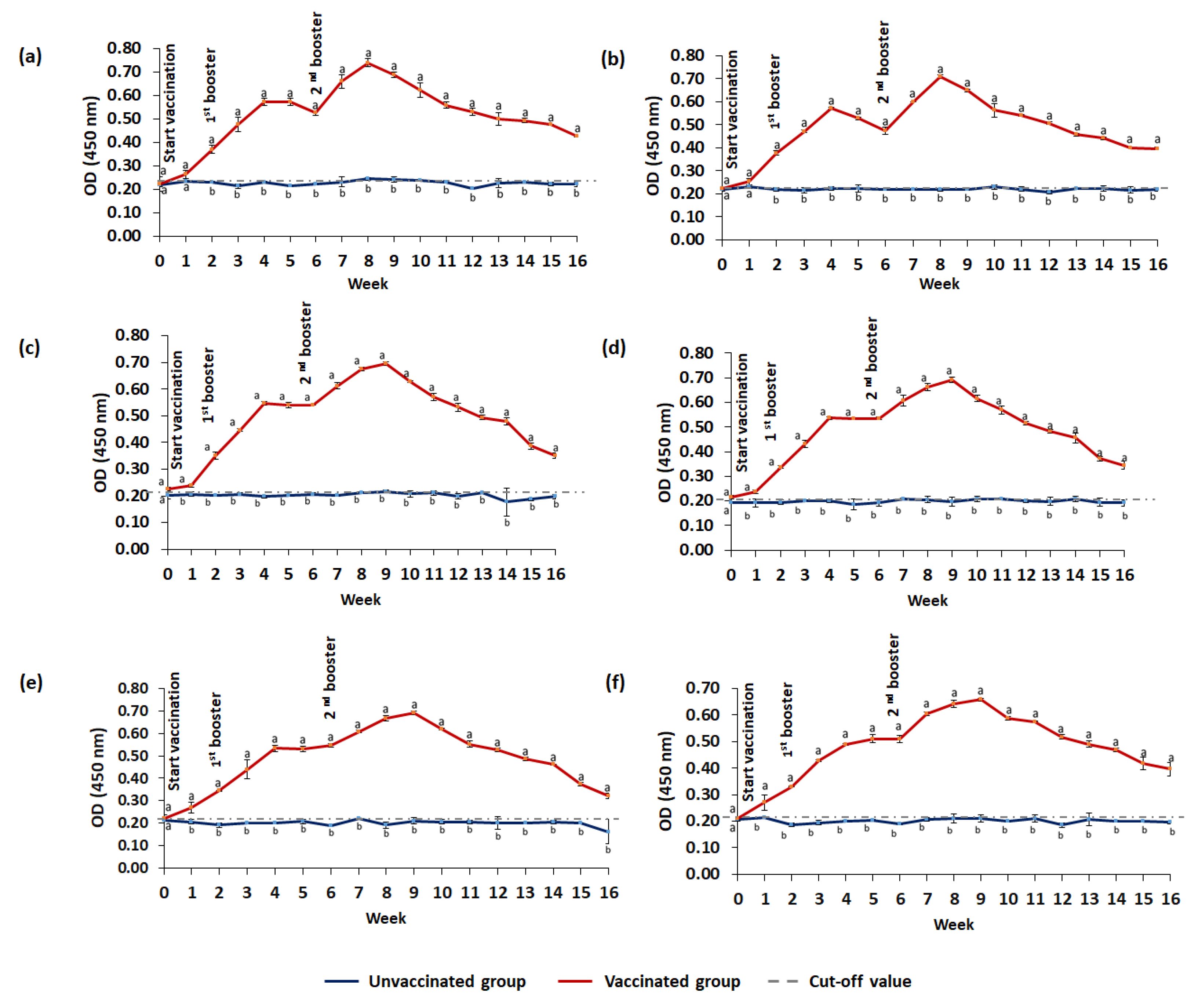
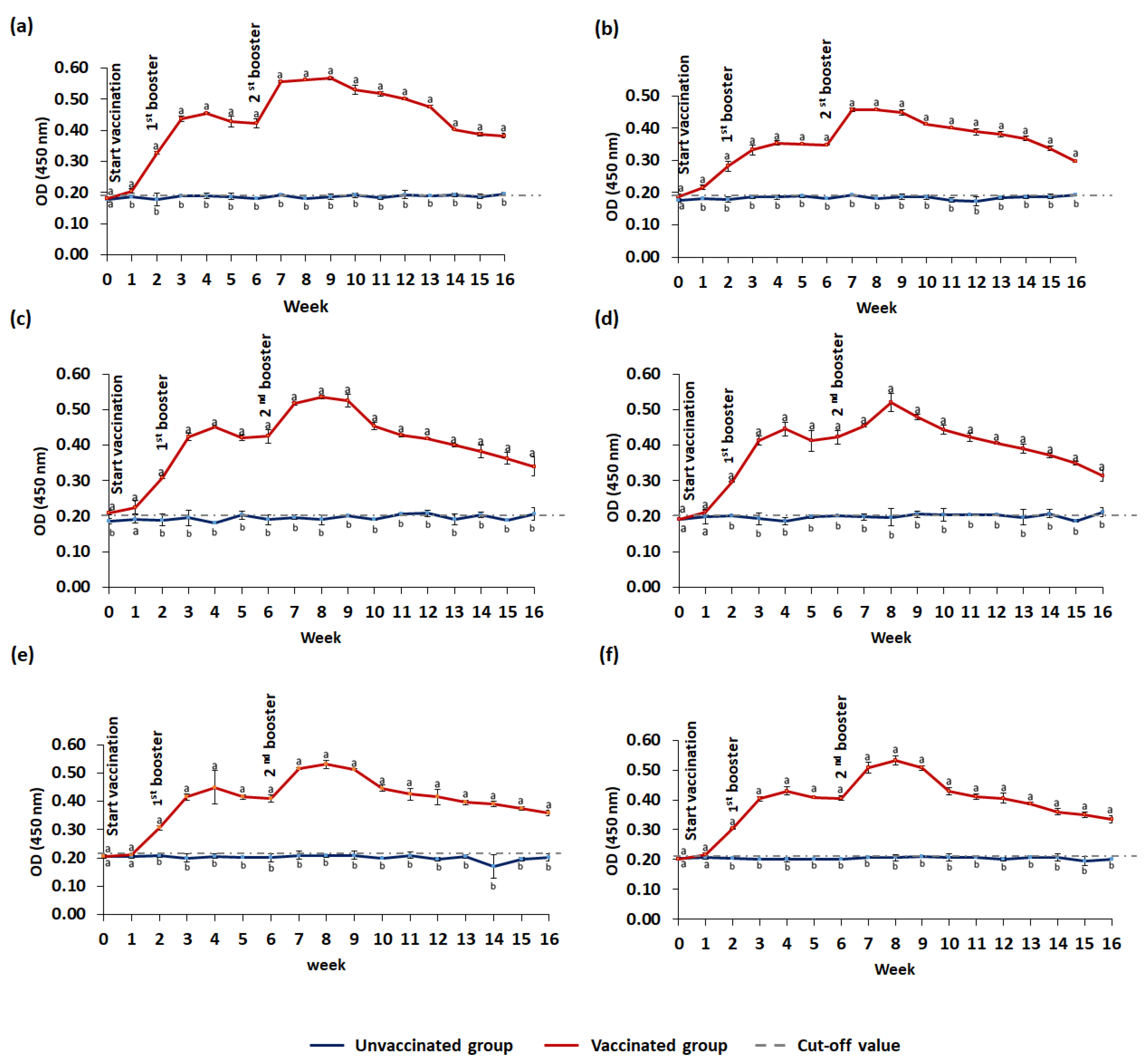
3.3.2. IgM Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2022. Repurposing Food and Agricultural Policies to Make Healthy Diets More Affordable; FAO: Rome, Italy, 2022. [Google Scholar]
- Zeng, W.; Wang, Y.; Chen, X.; Wang, Q.; Bergmann, S.M.; Yang, Y.; Wang, Y.; Li, B.O.; Lv, Y.; Li, H.; et al. Potency and efficacy of VP20-based vaccine against tilapia lake virus using different prime-boost vaccination regimens in tilapia. Aquaculture 2021, 539, 736654. [Google Scholar] [CrossRef]
- Mohamad, S.N.; Noordin, W.N.M.; Ismail, N.F.; Hamzah, A. Red hybrid tilapia (Oreochromis spp.) broodstock development programme in Malaysia: Status, challenges and prospects for future development. Asian Fish. Sci. 2021, 34, 73–81. [Google Scholar] [CrossRef]
- Lim, S.Y.; Ooi, A.L.; Wong, W.L. Gill monogeneans of Nile tilapia (Oreochromis niloticus) and red hybrid tilapia (Oreochromis spp.) from the wild and fish farms in Perak, Malaysia: Infection dynamics and spatial distribution. SpringerPlus 2016, 5, 1609. [Google Scholar] [CrossRef] [Green Version]
- FAO. Statistics and Information Branch, Fisheries and Aquaculture Department/FAO. Fishery and Aquaculture Statistics. Global Production by Production Source 1950–2016 (FishstatJ). FAO: Rome, Italy, 2018. [Google Scholar]
- Abdel-Latif, H.M.; Dawood, M.A.; Menanteau-Ledouble, S.; El-Matbouli, M. The nature and consequences of co-infections in tilapia: A review. J. Fish Dis. 2020, 43, 651–664. [Google Scholar] [CrossRef]
- Cui, M.; Zhang, H.; Li, J.; Liu, R.; Wu, M.; Xu, D.; Zhang, Q. Differential PCR detection of Streptococcus agalactiae and Streptococcus iniae in tilapia (Oreochromis niloticus) by a single primer pair. J. Fish Dis. 2019, 42, 1211–1216. [Google Scholar] [CrossRef]
- Wang, J.; Du, J.J.; Jiang, B.; He, R.Z.; Li, A.X. Effects of short-term fasting on the resistance of Nile tilapia (Oreochromis niloticus) to Streptococcus agalactiae infection. Fish Shellfish Immunol. 2019, 94, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.T.; Techatanakitarnan, C.; Jindakittikul, P.; Thaiprayoon, A.; Taengphu, S.; Charoensapsri, W.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2017, 40, 1395–1403. [Google Scholar] [CrossRef]
- AlYahya, S.A.; Ameen, F.; Al-Niaeem, K.S.; Al-Sa’adi, B.A.; Hadi, S.; Mostafa, A.A. Histopathological studies of experimental Aeromonas hydrophila infection in blue tilapia, Oreochromis aureus. Saudi J. Biol. Sci. 2018, 25, 182–185. [Google Scholar] [CrossRef]
- Basri, L.; Nor, R.M.; Salleh, A.; Md Yasin, I.S.; Saad, M.Z.; Abd. Rahaman, N.Y.; Barkham, T.; Amal, M.N.A. Co-infections of tilapia lake virus, Aeromonas hydrophila and Streptococcus agalactiae in farmed red hybrid tilapia. Animals 2020, 10, 2141. [Google Scholar] [CrossRef]
- Assis, G.B.N.; Tavares, G.C.; Pereira, F.L.; Figueiredo, H.C.P.; Leal, C.A.G. Natural coinfection by Streptococcus agalactiae and Francisella noatunensis subsp. orientalis in farmed Nile tilapia (Oreochromis niloticus L.). J. Fish Dis. 2017, 40, 51–63. [Google Scholar] [CrossRef]
- Dong, H.T.; Nguyen, V.V.; Le, H.D.; Sangsuriya, P.; Jitrakorn, S.; Saksmerprome, V.; Senapin, S.; Rodkhum, C. Naturally concurrent infections of bacterial and viral pathogens in disease outbreaks in cultured Nile tilapia (Oreochromis niloticus) farms. Aquaculture 2015, 448, 427–435. [Google Scholar] [CrossRef]
- Assane, I.M.; Gozi, K.S.; Valladão, G.M.R.; Pilarski, F. Combination of antimicrobials as an approach to reduce their application in aquaculture: Emphasis on the use of thiamphenicol/florfenicol against Aeromonas hydrophila. Aquaculture 2019, 507, 238–245. [Google Scholar] [CrossRef]
- Amal, M.N.A.; Zamri-Saad, M. Streptococcosis in Tilapia (Oreochromis niloticus): A Review. Pertanika J. Trop. Agric. Sci. 2011, 34, 195–206. [Google Scholar]
- Shirajum Monir, M.; Yusoff, S.M.; Mohamad, A.; Ina-Salwany, M.Y. Vaccination of Tilapia against Motile Aeromonas Septicemia: A Review. J. Aquat. Anim. Health 2020, 32, 65–76. [Google Scholar] [CrossRef]
- Watts, J.E.; Schreier, H.J.; Lanska, L.; Hale, M.S. The rising tide of antimicrobial resistance in aquaculture: Sources, sinks and solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignesh, R.; Karthikeyan, B.S.; Periyasamy, N.; Devanathan, K. Antibiotics in Aquaculture: An Overview. South Asian J. Exp. Biol. 2011, 1, 114–120. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H. Major bacterial diseases in aquaculture and their vaccine development. CAB Reviews 2012, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Assefa, A.; Abunna, F. Maintenance of fish health in aquaculture: Review of epidemiological approaches for prevention and control of infectious disease of fish. Vet. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef] [PubMed]
- Plant, K.P.; LaPatra, S.E. Advances in fish vaccine delivery. Dev. Comp. Immunol. 2011, 35, 1256–1262. [Google Scholar] [CrossRef]
- Ram, M.K.; Kumar, B.N.; Poojary, S.R.; Abhiman, P.B.; Patil, P.; Ramesh, K.S.; Shankar, K.M. Evaluation of biofilm of Vibrio anguillarum for oral vaccination of Asian seabass, Lates calcarifer (BLOCH, 1790). Fish Shellfish Immunol. 2019, 94, 746–751. [Google Scholar] [CrossRef]
- Monir, M.S.; Yusoff, S.B.M.; Mohamad, A.; Ngoo, M.S.B.M.H.; Ina-Salwany, M.Y. Haemato-immunological responses and effectiveness of feed-based bivalent vaccine against Streptococcus iniae and Aeromonas hydrophila infections in hybrid red tilapia (Oreochromis mossambicus × O. niloticus). Biotechnol. Adv. 2020, 16, 226. [Google Scholar] [CrossRef]
- Monir, M.S.; Yusoff, M.S.M.; Zulperi, Z.M.; Hassim, H.A.; Zamri-Saad, M.; Amal, M.N.A.; Salleh, A.; Mohamad, A.; Yie, L.J.; Ina-Salwany, M.Y. Immuno-protective efficiency of feed-based whole-cell inactivated bivalent vaccine against Streptococcus and Aeromonas infections in red hybrid tilapia (Oreochromis niloticus × Oreochromis mossambicus). Fish Shellfish Immunol. 2021, 113, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Amal, M.N.A.; Zamri-Saad, M.; Siti-Zahrah, A.; Zulkafli, A.R.; Nur-Nazifah, M. Molecular characterization of Streptococcus agalactiae strains isolated from fishes in Malaysia. J. Appl. Microbiol. 2013, 115, 20–29. [Google Scholar] [CrossRef]
- Matusin, S. Molecular Characterization of Aeromonas hydrophila and Development of Recombinant Cells Vaccine Expressing Outer Membrane Proteins against Its in African Catfish (Clarias gariepinus Burchell). Ph.D. Thesis, Dissertation, Universiti Putra Malaysia, Selangor, Malaysia, 2015. [Google Scholar]
- Rahmatullah, M.; Ariff, M.; Kahieshesfandiari, M.; Daud, H.M.; Zamri-Saad, M.; Sabri, M.Y.; Amal, M.N.A.; Ina-Salwany, M.Y. Isolation and pathogenicity of Streptococcus iniae in cultured red hybrid tilapia in Malaysia. J. Aquat. Anim. Health 2017, 29, 208–213. [Google Scholar] [CrossRef]
- Amal, M.N.A.; Koh, C.B.; Nurliyana, M.; Suhaiba, M.; Nor-Amalina, Z.; Santha, S.; Diyana-Nadhirah, K.P.; Yusof, M.T.; Ina-Salwany, M.Y.; Zamri-Saad, M. A case of natural co-infection of Tilapia Lake Virus and Aeromonas veronii in a Malaysian red hybrid tilapia (Oreochromis niloticus × O. mossambicus) farm experiencing high mortality. Aquaculture 2018, 485, 12–16. [Google Scholar] [CrossRef]
- Wohlsen, T.; Bates, J.; Vesey, G.; Robinson, W.A.; Katouli, M. Evaluation of the methods for enumerating coliform bacteria from water samples using precise reference standards. Lett. Appl. Microbiol. 2006, 42, 350–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.S.; Syafiq, M.R.; Siti-Zahrah, A.; Fahmi, S.; Shahidan, H.; Hanan, Y.; Amal, M.N.A.; Saad, M.Z. The effect of feed-based vaccination on tilapia farm endemic for streptococcosis. Fish Shellfish Immunol. 2017, 60, 21–24. [Google Scholar] [CrossRef]
- Dong, C.; He, G.; Mai, K.; Zhou, H.; Xu, W. Palatability of water-soluble extracts of protein sources and replacement of fishmeal by a selected mixture of protein sources for juvenile turbot (Scophthalmus maximus). J. Ocean Univ. China 2016, 15, 561–567. [Google Scholar] [CrossRef]
- Obaldo, L.G.; Divakaran, S.; Tacon, A.G. Method for determining the physical stability of shrimp feeds in water. Aquac. Res. 2002, 33, 369–377. [Google Scholar] [CrossRef]
- Abu Nor, N.; Zamri-Saad, M.; Md Yasin, I.S.; Salleh, A.; Mustaffa-Kamal, F.; Matori, M.F.; Azmai, M.N.A. Efficacy of whole cell inactivated Vibrio harveyi vaccine against vibriosis in a marine red hybrid tilapia (Oreochromis niloticus × O. mossambicus) model. Vaccines 2020, 8, 734. [Google Scholar] [CrossRef]
- Nur-Nazifah, M.; Sabri, M.Y.; Siti-Zahrah, A. Development and efficacy of feed-based recombinant vaccine encoding the cell wall surface anchor family protein of Streptococcus agalactiae against streptococcosis in Oreochromis sp. Fish Shellfish Immunol. 2014, 37, 193–200. [Google Scholar] [CrossRef]
- Byadgi, O.; Uyen, N.H.N.; Chou, R.L.; Guo, J.J.; Lee, Y.H.; Lee, J.W.; Cheng, T.C. Immunogenicity of inactivated formalin-killed Photobacterium damselae subsp. piscicida combined with Toll-like receptor 9 agonist in Cobia Rachycentron canadum. Aquaculture 2018, 492, 369–378. [Google Scholar] [CrossRef]
- Firdaus-Nawi, M.; Yusoff, S.M.; Yusof, H.; Abdullah, S.Z.; Zamri-Saad, M. Efficacy of feed-based adjuvant vaccine against Streptococcus agalactiae in Oreochromis spp. in Malaysia. Aquac. Res. 2013, 45, 87–96. [Google Scholar] [CrossRef]
- Syuhada, R.; Zamri-Saad, M.; Ina-Salwany, M.Y.; Mustafa, M.; Nasruddin, N.N.; Desa, M.N.M.; Nordin, S.A.; Barkham, T.; Amal, M.N.A. Molecular characterization and pathogenicity of Streptococcus agalactiae serotypes Ia ST7 and III ST283 isolated from cultured red hybrid tilapia in Malaysia. Aquaculture 2020, 515, 734543. [Google Scholar] [CrossRef]
- Amal, M.N.A.; Roseli, M.; Aidil, M.F.; Sutra, J.; Roslan, S.; Mazlan, S.N.K.; Basri, L.; Yie, L.J.; Mohamad, N.; Ismail, A.; et al. Javanese Medaka (Oryzias javanicus Bleeker, 1854) as potential model organism for aeromoniasis and vibriosis study in fish. Pertanika J. Trop. Agric. Sci. 2019, 42, 1049–1065. [Google Scholar]
- Pauzi, N.A.; Mohamad, N.; Azzam-Sayuti, M.; Yasin, I.S.M.; Saad, M.Z.; Nasruddin, N.S.; Azmai, M.N.A. Antibiotic susceptibility and pathogenicity of Aeromonas hydrophila isolated from red hybrid tilapia (Oreochromis niloticus × Oreochromis mossambicus) in Malaysia. Vet. World 2020, 13, 2166–2171. [Google Scholar] [CrossRef]
- Bidin, N.R.; Amal, M.N.A.; Mohamad, N.; Salleh, A.; Yasin, I.S.M.; Nasruddin, N.S.; Saad, M.Z. Antibiotic sensitivity and pathogenicity of Aeromonas veronii isolated from diseased red hybrid tilapia in Malaysia. Pertanika J. Trop. Agric. Sci. 2019, 42, 1263–1271. [Google Scholar]
- Laith, A.A.; Ambak, M.A.; Hassan, M.; Sheriff, S.M.; Nadirah, M.; Draman, A.S.; Wahab, W.; Ibrahim, W.N.W.; Aznan, A.S.; Jabar, A.; et al. Molecular identification and histopathological study of natural Streptococcus agalactiae infection in hybrid tilapia (Oreochromis niloticus). Vet. World 2017, 10, 101–111. [Google Scholar]
- Zhou, M.; Yu, S.; Hong, B.; Li, J.; Han, H.; Qie, G. Antibiotics control in aquaculture requires more than antibiotic-free feeds: A tilapia farming case. Environ. Pollut. 2021, 268, 115854. [Google Scholar] [CrossRef]
- Chen, J.; Huang, L.; Wang, Q.; Zeng, H.; Xu, J.; Chen, Z. Antibiotics in aquaculture ponds from Guilin, South of China: Occurrence, distribution, and health risk assessment. Environ. Res. 2022, 204, 112084. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Chen, D.D.; Cui, Z.W.; Zhang, X.Y.; Zhou, Y.Y.; Guo, X.; Li, A.H.; Zhang, Y.A. Oral vaccination of tilapia against Streptococcus agalactiae using Bacillus subtilis spores expressing Sip. Fish Shellfish Immunol. 2019, 86, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yang, Q.; Huang, L.; Wang, K.; Wang, X.; Chen, D.; Geng, Y.; Huang, X.; Ouyang, P.; Lai, W. Effectivity of oral recombinant DNA vaccine against Streptococcus agalactiae in Nile tilapia. Dev. Comp. Immunol. 2017, 77, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, A.; Zamri-Saad, M.; Amal, M.N.A.; Al-Saari, N.; Monir, M.S.; Chin, Y.K.; Md Yasin, I.S. Vaccine efficacy of a newly developed feed-based whole-cell polyvalent vaccine against vibriosis, streptococcosis and motile aeromonad septicemia in asian seabass, Lates calcarifer. Vaccines 2021, 9, 368. [Google Scholar] [CrossRef]
- Ridzuan, M.S.M.; Abdullah, A.; Ramly, R.; Mansor, N.N.; Ramli, N.; Firdaus-Nawi, M. Current status and advances of fish vaccines in Malaysia. Vet. World 2022, 15, 465–482. [Google Scholar] [CrossRef]
- Aich, N.; Paul, A.; Choudhury, T.G.; Saha, H. Tilapia Lake Virus (TiLV) disease: Current status of understanding. Aquac. Fish 2022, 7, 7–17. [Google Scholar] [CrossRef]
- Kai-yu, W.; De-fang, C.; Ling-yuan, H.; Hai, L.; Jun, W.; Dan, X.; Yi, G.; Ze-xiao, Y.; Wei-ming, L. Isolation and characterization of Streptococcus agalactiae from Nile Tilapia Oreochromis niloticus in China. Afr. J. Microbiol. Res. 2013, 7, 317–323. [Google Scholar] [CrossRef]
- Asencios, Y.O.; Sánchez, F.B.; Mendizábal, H.B.; Pusari, K.H.; Alfonso, H.O.; Sayán, A.M.; Figueiredo, M.A.P.; Manrique, W.G.; de Andrade Belo, M.A.; Chaupe, N.S. First report of Streptococcus agalactiae isolated from Oreochromis niloticus in Piura, Peru: Molecular identification and histopathological lesions. Aquac. Rep. 2016, 4, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Zamri-Saad, M.; Amal, M.N.A.; Siti-Zahrah, A.; Zulkafli, A.R. Control and prevention of streptococcosis in cultured tilapia in Malaysia: A Review. Pertanika J. Trop. Agric. Sci. 2014, 37, 389–410. [Google Scholar]
- Hamid, N.H.; Hassan, M.D.; Md Sabri, M.Y.; Hasliza, A.H.; Hamdan, R.H.; Afifah, M.N.; Raina, M.S.; Nadia, A.; Fuad, M.M. Studies on Pathogenicity Effect of Aeromonas hydrophila Infection in Juvenile Red Hybrid Tilapia Oreochromis sp. In Proceedings of the International Seminar on Livestock Production and Veterinary Technology, Denpasar, Indonesia, 10–12 August 2016; pp. 532–539. [Google Scholar]
- Craig, S.R.; Helfrich, L.A.; Kuhn, D.; Schwarz, M.H. Understanding Fish Nutrition, Feeds, and Feeding; Virginia Polytechnic Institute and State University: Blacksburg, VA, USA, 2009. [Google Scholar]
- FAO. Nile tilapia—Nutritional Requirements. Food and Agriculture Organization of the United Nations. 2017. Available online: http://www.fao.org/fishery/affris/species-profiles/nile-tilapia/nutritionalrequirements/en/ (accessed on 12 November 2022).
- Bahurmiz, O.M.; Ng, W.K. Effects of dietary palm oil source on growth, tissue fatty acid composition and nutrient digestibility of red hybrid tilapia, Oreochromis sp., raised from stocking to marketable size. Aquaculture 2007, 262, 382–392. [Google Scholar] [CrossRef]
- Ayisi, C.L.; Zhao, J.; Rupia, E.J. Growth performance, feed utilization, body and fatty acid composition of Nile tilapia (Oreochromis niloticus) fed diets containing elevated levels of palm oil. Aquac. Fish. 2017, 2, 67–77. [Google Scholar] [CrossRef]
- Suwannasang, A.; Suanyuk, N.; Issaro, A.; Phromkunthong, W.; Tantikitti, C.; Itami, T.; Yoshida, T. Growth, immune responses and protection of Nile tilapia Oreochromis niloticus immunized with formalin-killed Streptococcus agalactiae serotype Ia and III vaccines. Songklanakarin J. Sci. Technol. 2017, 39, 429–437. [Google Scholar]
- Kahiesh Esfandiari, M.; Ina-Salwany, M.Y.; Aslah, M.; Soltani, M.; Sabili, A.; Karimi, M.; Muthukrishnan, S. Growth performance, palatability and water stability of oral feed-based vaccines against Streptococcus agalactiae in red tilapia (Oreochromis sp.). J. Biochem. Tech. 2019, 10, 106–115. [Google Scholar]
- Giri, S.S.; Sen, S.S.; Jun, J.W.; Sukumaran, V.; Park, S.C. Role of Bacillus subtilis VSG4-derived biosurfactant in mediating immune responses in Labeo rohita. Fish Shellfish Immunol. 2016, 54, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Feng, F.; Wang, Y.; Guo, L.; Chen, L.; Chen, K. Advances in research into oral vaccines for fish. Int. J. Fish. Aquac. 2018, 8, 19–40. [Google Scholar]
- Jiao, X.D.; Cheng, S.; Hu, Y.H.; Sun, L. Comparative study of the effects of aluminum adjuvants and Freund’s incomplete adjuvant on the immune response to an Edwardsiella tarda major antigen. Vaccine 2010, 28, 1832–1837. [Google Scholar] [CrossRef]
- Cai, X.; Wang, B.; Peng, Y.; Li, Y.; Lu, Y.; Huang, Y.; Jian, J.; Wu, Z. Construction of a Streptococcus agalactiae phoB mutant and evaluation of its potential as an attenuated modified live vaccine in golden pompano, Trachinotus ovatus. Fish Shellfish Immunol. 2017, 63, 405–416. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Description | Standard Dietary | Commercial Tilapia Pellet | Formulated Pellet | |
|---|---|---|---|---|
| Control | Bivalent Vaccine | |||
| Carbohydrate (%) | >25 | 44.26 ± 0.15 a | 40.90 ± 0.47 b | 39.57 ± 0.13 c |
| Energy (%) | na | 354.00 ± 0.00 b | 397.50 ± 2.12 a | 395.00 ± 0.00 a |
| Moisture (%) | <10 | 7.87 ± 0.02 a | 6.08 ± 0.01 c | 7.73 ± 0.02 b |
| Protein (%) | 28–35 | 34.36 ± 0.02 a | 31.87 ± 0.12 b | 31.18 ± 0.04 c |
| Total ash (%) | <16 | 9.17 ± 0.02 b | 9.31 ± 0.01 a | 9.10 ± 0.01 c |
| Total fat (%) | 6–13 | 4.36 ± 0.02 c | 11.85 ± 0.37 b | 12.43 ± 0.05 a |
| Parameters | Treatment Group | |
|---|---|---|
| Group 1 Unvaccinated (Control) | Group 2 Vaccinated (Bivalent Vaccine) | |
| Initial body weight (g) | 12.94 ± 0.46 | 13.10 ± 0.72 |
| Final body weight (g) | 33.67 ± 0.98 | 35.83 ± 1.20 |
| Weight gain (g) | 20.73 ± 0.97 | 22.73 ± 1.83 |
| SGR (%/day) | 1.59 ± 0.49 | 1.68 ± 0.35 |
| FCR (g/g) | 1.97 ± 0.07 | 1.82 ± 0.10 |
| Challenge Bacteria | Treatment Group | Challenge Dose (CFU/mL) | Dead/ Total | Mortality (%) | RPS (%) |
|---|---|---|---|---|---|
| S. agalactiae | Vaccinated | 1.8 × 103 | 6/30 | 20.00 | 80.00 ± 10.00 b |
| Unvaccinated | 30/30 | 100.00 | - | ||
| A. hydrophila | Vaccinated | 2.0 × 104 | 3/30 | 10.00 | 90.00 ± 10.00 a |
| Unvaccinated | 30/30 | 100.00 | - | ||
| S. iniae | Vaccinated | 1.5 × 103 | 11/30 | 36.66 | 63.33 ± 5.77 c |
| Unvaccinated | 30/30 | 100.00 | - | ||
| A. veronii | Vaccinated | 1.8 × 104 | 12/30 | 40.00 | 60.00 ± 10.00 c |
| Unvaccinated | 30/30 | 100.00 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Ali, N.S.; Saad, M.Z.; Azmai, M.N.A.; Salleh, A.; Zulperi, Z.M.; Manchanayake, T.; Zahaludin, M.A.D.; Basri, L.; Mohamad, A.; Md Yasin, I.S. Immunogenicity and Efficacy of a Feed-Based Bivalent Vaccine against Streptococcosis and Motile Aeromonad Septicemia in Red Hybrid Tilapia (Oreochromis sp.). Animals 2023, 13, 1346. https://doi.org/10.3390/ani13081346
Mohd Ali NS, Saad MZ, Azmai MNA, Salleh A, Zulperi ZM, Manchanayake T, Zahaludin MAD, Basri L, Mohamad A, Md Yasin IS. Immunogenicity and Efficacy of a Feed-Based Bivalent Vaccine against Streptococcosis and Motile Aeromonad Septicemia in Red Hybrid Tilapia (Oreochromis sp.). Animals. 2023; 13(8):1346. https://doi.org/10.3390/ani13081346
Chicago/Turabian StyleMohd Ali, Nur Shidaa, Mohd Zamri Saad, Mohammad Noor Amal Azmai, Annas Salleh, Zarirah Mohamed Zulperi, Tilusha Manchanayake, Muhammad Amir Danial Zahaludin, Lukman Basri, Aslah Mohamad, and Ina Salwany Md Yasin. 2023. "Immunogenicity and Efficacy of a Feed-Based Bivalent Vaccine against Streptococcosis and Motile Aeromonad Septicemia in Red Hybrid Tilapia (Oreochromis sp.)" Animals 13, no. 8: 1346. https://doi.org/10.3390/ani13081346
APA StyleMohd Ali, N. S., Saad, M. Z., Azmai, M. N. A., Salleh, A., Zulperi, Z. M., Manchanayake, T., Zahaludin, M. A. D., Basri, L., Mohamad, A., & Md Yasin, I. S. (2023). Immunogenicity and Efficacy of a Feed-Based Bivalent Vaccine against Streptococcosis and Motile Aeromonad Septicemia in Red Hybrid Tilapia (Oreochromis sp.). Animals, 13(8), 1346. https://doi.org/10.3390/ani13081346