
Different Conditions during Confinement in Pasture-Based Systems and Feeding Systems Affect the Fatty Acid Profile in the Milk and Cheese of Holstein Dairy Cows
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Location, Animals, and Treatments
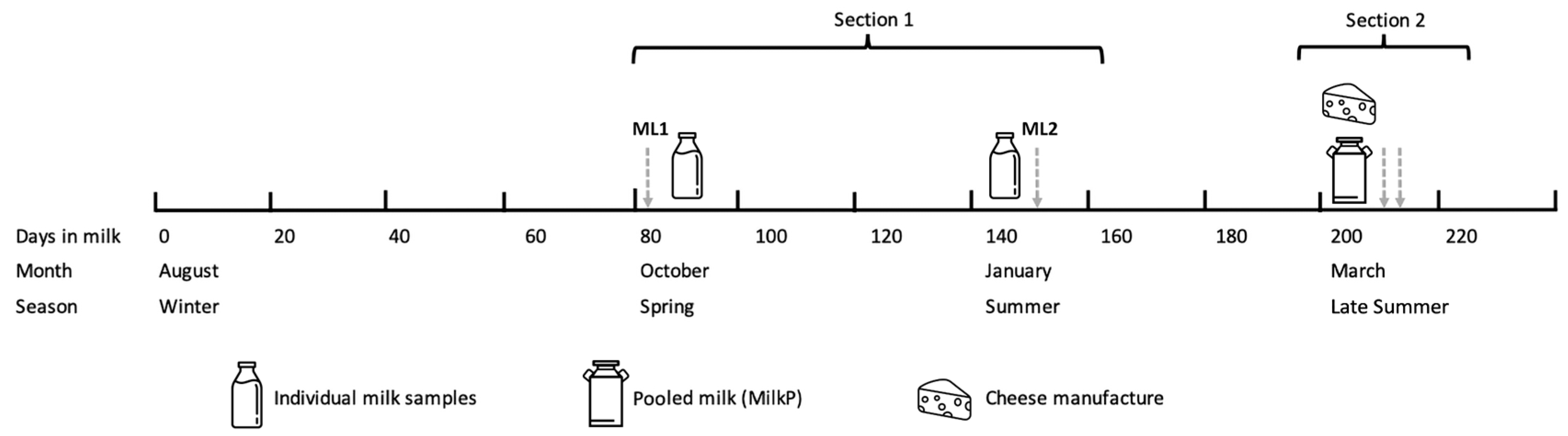
2.2. Experimental Design
2.2.1. Individual Cow Milk during Spring and Summer
2.2.2. Pooled Milk and Cheese Manufacture during Late Summer
2.3. Samples Analysis
2.3.1. Fat Yield
2.3.2. Fatty Acid Profile in Milk (Individual and Pooled) and Cheeses
2.3.3. Pasture and TMR Chemical Composition and Fatty Acid Profile
2.4. Statistical Analysis
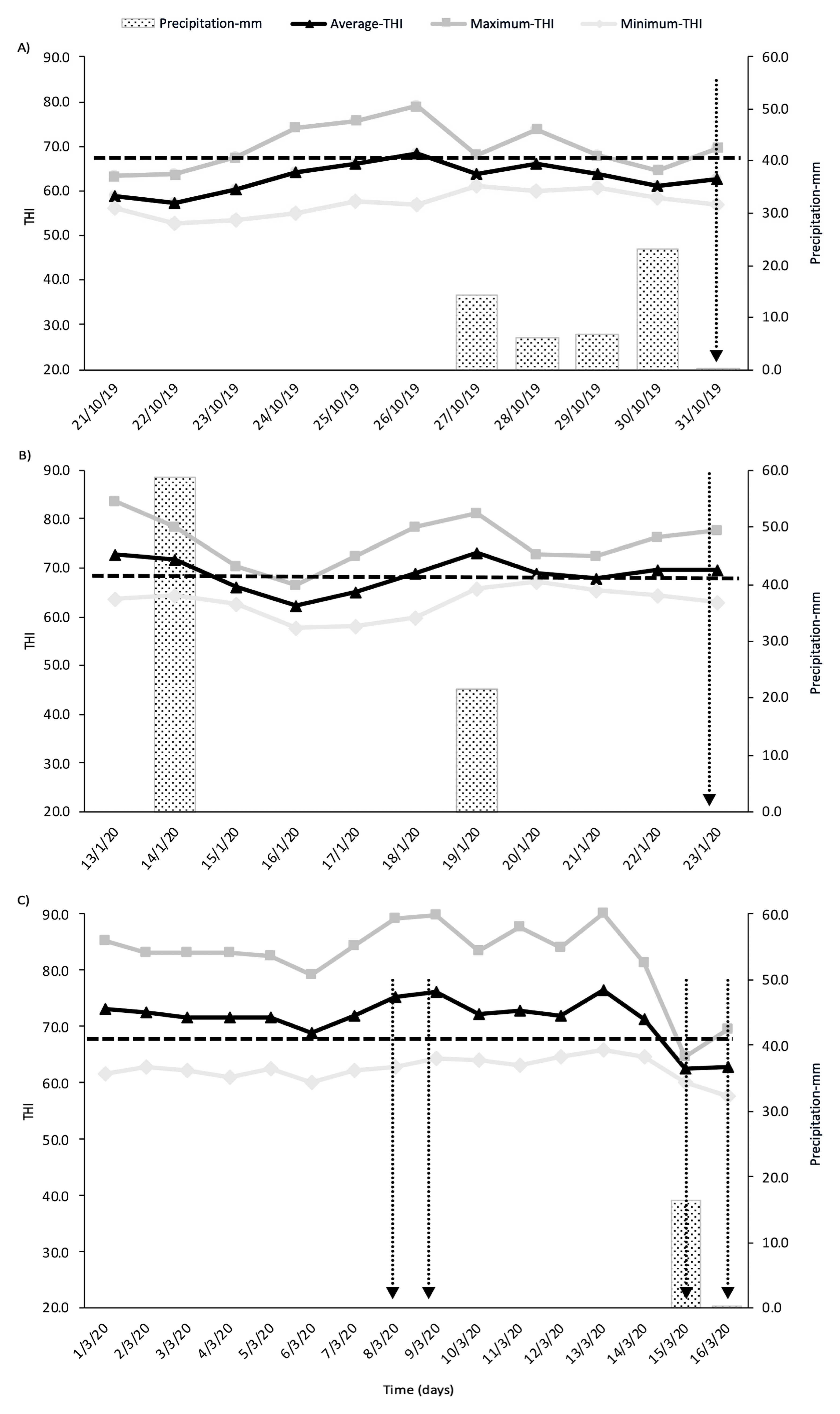
3. Results
3.1. Milk Yield and Fat (% and kg/d)
3.1.1. Individual Cow Milk during Spring and Summer
3.1.2. Fatty Acid Profile in Pooled Milk and Cheese
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- German, J.B.; Dillard, C.J. Composition, Structure and Absorption of Milk Lipids: A Source of Energy, Fat-Soluble Nutrients and Bioactive Molecules. Crit. Rev. Food Sci. Nutr. 2006, 46, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Moscovici Joubran, A.; Pierce, K.M.; Garvey, N.; Shalloo, L.; O’Callaghan, T.F. Invited Review: A 2020 Perspective on Pasture-Based Dairy Systems and Products. J. Dairy Sci. 2021, 104, 7364–7382. [Google Scholar] [CrossRef] [PubMed]
- Lock, A.L.; Bauman, D.E. Modifying Milk Fat Composition of Dairy Cows to Enhance Fatty Acids Beneficial to Human Health. Lipids 2004, 39, 1197–1206. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Mansbridge, R.M.; Doreau, M. Ruminant Milk Fat Plasticity: Nutritional Control of Saturated, Polyunsaturated, Trans and Conjugated Fatty Acids. Ann. Zootech. 2000, 49, 181–205. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.F.; Scollan, N.D. Increasing the Concentrations of Beneficial Polyunsaturated Fatty Acids in Milk Produced by Dairy Cows in High-Forage Systems. Anim. Feed. Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Barca, J.; Carriquiry, M.; Olazabal, L.; Fajardo, M.; Chilibroste, P.; Meikle, A. Milk Fatty Acid Profile from Cows Fed with Mixed Rations and Different Access Time to Pastureland during Early Lactation. J. Anim. Physiol. Anim. Nutr. 2017, 102, 620–629. [Google Scholar] [CrossRef]
- Grille, L.; Adrien, M.L.; Méndez, M.N.; Chilibroste, P.; Olazabal, L.; Damián, J.P. Milk Fatty Acid Profile of Holstein Cows When Changed from a Mixed System to a Confinement System or Mixed System with Overnight Grazing. Int. J. Food Sci. 2022, 2022, 5610079. [Google Scholar] [CrossRef]
- Elgersma, A. Grazing Increases the Unsaturated Fatty Acid Concentration of Milk from Grass-Fed Cows: A Review of the Contributing Factors, Challenges and Future Perspectives. Eur. J. Lipid Sci. Technol. 2015, 117, 1345–1369. [Google Scholar] [CrossRef]
- Alothman, M.; Hogan, S.A.; Hennessy, D.; Dillon, P.; Kilcawley, K.N.; O’donovan, M.; Tobin, J.; Fenelon, M.A.; O’callaghan, T.F. The “Grass-Fed” Milk Story: Understanding the Impact of Pasture Feeding on the Composition and Quality of Bovine Milk. Foods 2019, 8, 350. [Google Scholar] [CrossRef]
- Bauman, D.E.; Griinari, J.M. Regulation and Nutritional Manipulation of Milk Fat: Low-Fat Milk Syndrome. Livest. Prod. Sci. 2001, 70, 15–29. [Google Scholar] [CrossRef]
- Hu, F.B.; Manson, J.A.E.; Willett, W.C. Types of Dietary Fat and Risk of Coronary Heart Disease: A Critical Review. J. Am. Coll. Nutr. 2001, 20, 5–19. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, Rumen Biohydrogenation and Nutritional Quality of Cow and Goat Milk Fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Moate, P.J.; Chalupa, W.; Boston, R.C.; Lean, I.J. Milk Fatty Acids. I. Variation in the Concentration of Individual Fatty Acids in Bovine Milk. J. Dairy Sci. 2007, 90, 4730–4739. [Google Scholar] [CrossRef] [PubMed]
- Kucevic, D.; Trivunovic, S.; Bogdanovic, V.; Cobanovic, K.; Jankovic, D.; Stanojevic, D. Composition of Raw Milk from Conventional and Organic Dairy Farming. Biotehnol. Stocarstvu 2016, 32, 133–143. [Google Scholar] [CrossRef]
- Lucas, A.; Agabriel, C.; Martin, B.; Ferlay, A.; Verdier-Metz, I.; Coulon, J.-B.; Rock, E. Relationships between the Conditions of Cow’s Milk Production and the Contents of Components of Nutritional Interest in Raw Milk Farmhouse Cheese. Le Lait 2006, 86, 177–202. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Mannion, D.T.; Hennessy, D.; McAuliffe, S.; O’Sullivan, M.G.; Leeuwendaal, N.; Beresford, T.P.; Dillon, P.; Kilcawley, K.N.; Sheehan, J.J.; et al. Effect of Pasture versus Indoor Feeding Systems on Quality Characteristics, Nutritional Composition, and Sensory and Volatile Properties of Full-Fat Cheddar Cheese. J. Dairy Sci. 2017, 100, 6053–6073. [Google Scholar] [CrossRef] [PubMed]
- Frétin, M.; Martin, B.; Buchin, S.; Desserre, B.; Lavigne, R.; Tixier, E.; Cirié, C.; Bord, C.; Montel, M.-C.; Delbès, C.; et al. Milk Fat Composition Modifies the Texture and Appearance of Cantal-Type Cheeses but Not Their Flavor. J. Dairy Sci. 2019, 102, 1131–1143. [Google Scholar] [CrossRef]
- Lee, B.-H.; Corazzin, M.; Piasentier, E.; Huh, C.-S.; Sung, K.-I.; Park, S.-Y. Milk and Montasio-Type Cheese Fatty Acid Composition from Cows Grazing on Timothy and Reed Canarygrass Pasture or Fed Indoor. Grassl. Sci. 2019, 65, 226–232. [Google Scholar] [CrossRef]
- Charlton, G.L.; Rutter, S.M. The Behaviour of Housed Dairy Cattle with and without Pasture Access: A Review. Appl. Anim. Behav. Sci. 2017, 192, 2–9. [Google Scholar] [CrossRef]
- O’callaghan, T.F.; Hennessy, D.; McAuliffe, S.; Kilcawley, K.N.; O’donovan, M.; Dillon, P.; Ross, R.P.; Stanton, C. Effect of Pasture versus Indoor Feeding Systems on Raw Milk Composition and Quality over an Entire Lactation. J. Dairy Sci. 2016, 99, 9424–9440. [Google Scholar] [CrossRef]
- Hennessy, D.; Delaby, L.; van den Pol-van Dasselaar, A.; Shalloo, L. Increasing Grazing in Dairy Cow Milk Production Systems in Europe. Sustainability 2020, 12, 2443. [Google Scholar] [CrossRef]
- Fariña, S.R.; Chilibroste, P. Opportunities and Challenges for the Growth of Milk Production from Pasture: The Case of Farm Systems in Uruguay. Agric. Syst. 2019, 176, 102631. [Google Scholar] [CrossRef]
- Méndez, M.N.; Grille, L.; Mendina, G.R.; Robinson, P.H.; Adrien, M.d.L.; Meikle, A.; Chilibroste, P. Performance of Autumn and Spring Calving Holstein Dairy Cows with Different Levels of Environmental Exposure and Feeding Strategies. Animals 2023, 13, 1211. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.E.; Rogers, A.R.; Poulouin, Y.A.; Cox, N.R.; Tucker, C.B. The Amount of Shade Influences the Behavior and Physiology of Dairy Cattle. J. Dairy Sci. 2010, 93, 125–133. [Google Scholar] [CrossRef]
- Van Laer, E.; Tuyttens, F.A.M.; Ampe, B.; Sonck, B.; Moons, C.P.H.; Vandaele, L. Effect of Summer Conditions and Shade on the Production and Metabolism of Holstein Dairy Cows on Pasture in Temperate Climate. Animal 2015, 9, 1547–1558. [Google Scholar] [CrossRef]
- Kadzere, C.T.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat Stress in Lactating Dairy Cows: A Review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Moretti, R.; Biffani, S.; Chessa, S.; Bozzi, R. Heat Stress Effects on Holstein Dairy Cows’ Rumination. Animal 2017, 11, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, F.; Bragaglio, A.; Sabia, E.; Serrapica, F.; Braghieri, A.; De Rosa, G. The Human–Animal Relationship in Dairy Animals. J. Dairy Res. 2020, 87, 47–52. [Google Scholar] [CrossRef]
- Bernabucci, U.; Basiricò, L.; Morera, P.; Dipasquale, D.; Vitali, A.; Cappelli, F.; Calamari, L. Effect of Summer Season on Milk Protein Fractions in Holstein Cows. J. Dairy Sci. 2015, 98, 1815–1827. [Google Scholar] [CrossRef]
- Becker, C.A.; Collier, R.J.; Stone, A.E. Invited Review: Physiological and Behavioral Effects of Heat Stress in Dairy Cows. J. Dairy Sci. 2020, 103, 6751–6770. [Google Scholar] [CrossRef]
- Morales-Piñeyrúa, J.T.; Damián, J.P.; Banchero, G.; Sant Anna, A.C. The Effects of Heat Stress on Milk Production and the Grazing Behavior of Dairy Holstein Cows Milked by an Automatic Milking System. J. Anim. Sci. 2022, 100, skac225. [Google Scholar] [CrossRef]
- Maggiolino, A.; Dahl, G.E.; Bartolomeo, N.; Bernabucci, U.; Vitali, A.; Serio, G.; Cassandro, M.; Centoducati, G.; Santus, E.; De Palo, P. Estimation of Maximum Thermo-Hygrometric Index Thresholds Affecting Milk Production in Italian Brown Swiss Cattle. J. Dairy Sci. 2020, 103, 8541–8553. [Google Scholar] [CrossRef]
- Kumar, G.; Devi, P.; Sharma, N.; Yallappa, I.; Somagond, M.; Somagond, Y.M. Impact of Thermal Stress on Milk Production, Composition and Fatty Acid Profile in Dairy Cows: A Review. J. Entomol. Zool. Stud. 2020, 8, 1278–1283. [Google Scholar]
- Salamon, R.; Varga-Visi, É.; Sára, P.; Csapó-Kiss, Z.; Csapó, J. The Influence of the Season on the Fatty Acid Composition and Conjugated Linolic Acid Content of the Milk. Krmiva Časopis O Hranidbi Zivotinj. Proizv. I Tehnol. Krme 2006, 48, 193–200. [Google Scholar]
- Maniaci, G.; Di Grigoli, A.; Bonanno, A.; Giosuè, C.; Ilardi, V.; Alabiso, M. Fatty Acids as Biomarkers of the Production Season of Caciocavallo Palermitano Cheese. Animals 2021, 11, 2675. [Google Scholar] [CrossRef]
- Tzamaloukas, O.; Neofytou, M.C.; Simitzis, P.E.; Miltiadou, D. Effect of Farming System (Organic vs. Conventional) and Season on Composition and Fatty Acid Profile of Bovine, Caprine and Ovine Milk and Retail Halloumi Cheese Produced in Cyprus. Foods 2021, 10, 1016. [Google Scholar] [CrossRef]
- Paszczyk, B.; Polak-Śliwińska, M.; Zielak-Steciwko, A.E. Chemical Composition, Fatty Acid Profile, and Lipid Quality Indices in Commercial Ripening of Cow Cheeses from Different Seasons. Animals 2022, 12, 198. [Google Scholar] [CrossRef] [PubMed]
- Vibart, R.E.; Fellner, V.; Burns, J.C.; Huntington, G.B.; Green, J.T. Performance of Lactating Dairy Cows Fed Varying Levels of Total Mixed Ration and Pasture. J. Dairy Res. 2008, 75, 471–480. [Google Scholar] [CrossRef]
- Chen, J.M.; Stull, C.L.; Ledgerwood, D.N.; Tucker, C.B. Muddy Conditions Reduce Hygiene and Lying Time in Dairy Cattle and Increase Time Spent on Concrete. J. Dairy Sci. 2017, 100, 2090–2103. [Google Scholar] [CrossRef]
- Román, L.; Saravia, C.; Astigarraga, L.; Bentancur, O.; La Manna, A. Shade Access in Combination with Sprinkling and Ventilation Effects Performance of Holstein Cows in Early and Late Lactation. Anim. Prod. Sci. 2019, 59, 347. [Google Scholar] [CrossRef]
- INALE. Available online: https://www.inale.org/estadisticas/encuesta-lechera-2019-resultados-definitivos/ (accessed on 26 January 2023).
- Leso, L.; Barbari, M.; Lopes, M.A.; Damasceno, F.A.; Galama, P.; Taraba, J.L.; Kuipers, A. Invited Review: Compost-Bedded Pack Barns for Dairy Cows. J. Dairy Sci. 2020, 103, 1072–1099. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.M.; Robertson, L.M.; Eckelkamp, E.A. A 100-Year Review: Lactating Dairy Cattle Housing Management. J. Dairy Sci. 2017, 100, 10418–10431. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Masucci, F.; Napolitano, F.; Braghieri, A.; Romano, R.; Manzo, N.; Di Francia, A. Fatty Acid and Sensory Profiles of Caciocavallo Cheese as Affected by Management System. J. Dairy Sci. 2014, 97, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Bonanno, A.; Tornambè, G.; Bellina, V.; De Pasquale, C.; Mazza, F.; Maniaci, G.; Di Grigoli, A. Effect of Farming System and Cheesemaking Technology on the Physicochemical Characteristics, Fatty Acid Profile, and Sensory Properties of Caciocavallo Palermitano Cheese. J. Dairy Sci. 2013, 96, 710–724. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy of Sciences: Washington, DC, USA, 2001; Volume 519. [Google Scholar]
- Haydock, K.P.; Shaw, N.H. The Comparative Yield Method for Estimating Dry Matter Yield of Pasture. Aust. J. Exp. Agric. 1975, 15, 663–670. [Google Scholar] [CrossRef]
- Pastorini, M.; Pomiés, N.; Repetto, J.L.; Mendoza, A.; Cajarville, C. Productive Performance and Digestive Response of Dairy Cows Fed Different Diets Combining a Total Mixed Ration and Fresh Forage. J. Dairy Sci. 2019, 102, 4118–4130. [Google Scholar] [CrossRef]
- Mendoza, A.; Cajarville, C.; Repetto, J.L. Short Communication: Intake, Milk Production, and Milk Fatty Acid Profile of Dairy Cows Fed Diets Combining Fresh Forage with a Total Mixed Ration. J. Dairy Sci. 2016, 99, 1938–1944. [Google Scholar] [CrossRef]
- Escobar, D.; Pelaggio, R.; Grille, L.; Colzada, E.; Rampoldi, C.; Carro, S.; Delucci, I.; Viola, N.; Nolla, J.P.; Reinares, R.; et al. Effect of Caseins Profile, Somatic Cells Count and Milk Composition in Dambo Cheese Yield. Innotec 2014, 9, 31–42. [Google Scholar]
- Hara, A.; Radin, N.S. Lipid Extraction of Tissues. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipids from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Mossoba, M.M.; Yurawecz, M.P.; Roach, J.A.G.; McDonald, R.E.; Flickinger, B.D.; Perkins, E.G. Analysis of Cyclic Fatty Acid Monomer 2-Alkenyl-4,4-dimethyloxazoline Derivatives by Gas Chromatography−Matrix Isolation−Fourier Transform Infrared Spectroscopy. J. Agric. Food Chem. 1996, 44, 3193–3196. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary Heart Disease: Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- AOAC. Official Methods of Analysis, 14th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1984. [Google Scholar]
- Salado, E.E.; Bretschneider, G.; Cuatrin, A.; Descalzo, A.M.; Gagliostro, G.A. Productive Response of Dairy Cows Fed with Different Levels of Totally Mixed Ration and Pasture. Agric. Sci. 2018, 09, 824–851. [Google Scholar] [CrossRef]
- Bargo, F.; Muller, L.D.; Varga, G.A.; Delahoy, J.E.; Cassidy, T.W. Ruminal Digestion and Fermentation of High-Producing Dairy Cows with Three Different Feeding Systems Combining Pasture and Total Mixed Rations. J. Dairy Sci. 2002, 85, 2964–2973. [Google Scholar] [CrossRef] [PubMed]
- Mollica, M.P.; Trinchese, G.; Cimmino, F.; Penna, E.; Cavaliere, G.; Tudisco, R.; Musco, N.; Manca, C.; Catapano, A.; Monda, M.; et al. Milk Fatty Acid Profiles in Different Animal Species: Focus on the Potential Effect of Selected PUFAs on Metabolism and Brain Functions. Nutrients 2021, 13, 1111. [Google Scholar] [CrossRef]
- Ferlay, A.; Bernard, L.; Meynadier, A.; Malpuech-Brugère, C. Production of Trans and Conjugated Fatty Acids in Dairy Ruminants and Their Putative Effects on Human Health: A Review. Biochimie 2017, 141, 107–120. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Hirigoyen, D.; Santos, R.D.L.; Calvo, M.F.; González-Revello, A.; Constantin, M. Chemical Composition and Seasonal Changes in the Fatty Acid Profile of Uruguayan “Colonia” Cheeses. Grasas y Aceites 2018, 69, 254. [Google Scholar] [CrossRef]
- Kolver, E.S.; Muller, L.D. Performance and Nutrient Intake of High Producing Holstein Cows Consuming Pasture or a Total Mixed Ration. J. Dairy Sci. 1998, 81, 1403–1411. [Google Scholar] [CrossRef]
- Auldist, M.J.; Marett, L.C.; Greenwood, J.S.; Wright, M.M.; Hannah, M.; Jacobs, J.L.; Wales, W.J. Milk Production Responses to Different Strategies for Feeding Supplements to Grazing Dairy Cows. J. Dairy Sci. 2016, 99, 657–671. [Google Scholar] [CrossRef]
- Pons, V.; Damián, J.P.; Mattiauda, D.A.; Meikle, A.; Adrien, M.L.; Chilibroste, P. ¿Como Afectan Las Caracteristicas Del Encierro El Comportamiento de Vacas Lecheras En Sistemas Mixtos? In Proceedings of the IV Congreso de Colegio Latinoamericano de Bienestar Animal y Medicina del Comportamiento, Montevideo, Uruguay, 5–7 November 2021. [Google Scholar]
- Cook, N.B.; Bennett, T.B.; Nordlund, K.V. Monitoring Indices of Cow Comfort in Free-Stall-Housed Dairy Herds. J. Dairy Sci. 2005, 88, 3876–3885. [Google Scholar] [CrossRef] [PubMed]
- Krawczel, P.; Grant, R. Effects of Cow Comfort on Milk Quality, Productivity and Behavior. In Proceedings of the NMC Annual Meeting Proceedings, Charlotte, NC, USA, 25–28 January 2009; pp. 15–24. [Google Scholar]
- Grille, L.; Adrien, M.L.; Olmos, M.; Chilibroste, P.; Damián, J.P. Diet Change from a System Combining Total Mixed Ration and Pasture to Confinement System (Total Mixed Ration) on Milk Production and Composition, Blood Biochemistry and Behavior of Dairy Cows. Anim. Sci. J. 2019, 90, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.D.; Arney, D.R.; Phillips, C.J.C. Two- Or Four-Hour Lying Deprivation on the Behavior of Lactating Dairy Cows. J. Dairy Sci. 2007, 90, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.E.; Cave, V.M.; Cox, N.R.; Huddart, F.J.; Tucker, C.B. Effects of 3 Surface Types on Dairy Cattle Behavior, Preference, and Hygiene. J. Dairy Sci. 2019, 102, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Bernabucci, U.; Biffani, S.; Buggiotti, L.; Vitali, A.; Lacetera, N.; Nardone, A. The Effects of Heat Stress in Italian Holstein Dairy Cattle. J. Dairy Sci. 2014, 97, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Saravia, C.; Astigarraga, L.; Van Lier, E.; Bentancur, O. Impacto de Las Olas de Calor En Vacas Lecheras En Salto (Uruguay). Agrociencia Urug. 2011, 15, 93–102. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Individual Milk Samples | Cheese Manufacture | |||||
|---|---|---|---|---|---|---|
| Section 1 | Section 2 | |||||
| ML1 | ML2 | |||||
| TMR | Pasture | TMR | Pasture | TMR | Pasture | |
| CP (% DM) | 21.6 | 12.2 | 20.1 | 14.2 | 16.4 | 21.25 |
| NDF (% DM) | 34 | 59 | 33 | 26 | 26 | 44 |
| ADF (% DM) | 19 | 30 | 19 | 19 | 15 | 20 |
| FAs (g/100 g) | ||||||
| C8:0 | 0.07 | nd | 0.11 | nd | 0.5 | 0.7 |
| C10:0 | 0.04 | 1.56 | 0.02 | nd | 1.1 | 1.5 |
| C12:0 | 0.35 | 0.05 | 0.24 | 0.55 | 1.4 | 1.9 |
| C14:0 | 0.74 | 0.57 | 0.59 | 0.65 | 4.3 | 5.6 |
| C14:1 | 0.08 | 0.7 | 0.08 | 0.11 | 0.3 | 0.5 |
| C15:0 | 0.07 | 3.17 | 0.08 | 0.83 | 0.5 | 0.7 |
| C16:0 | 31.95 | 20.74 | 28.27 | 18.09 | 21.6 | 25.1 |
| C16:1 | 0.47 | 2.19 | 0.49 | 1.93 | 0.8 | 0.9 |
| C17:0 | 0.15 | 0.37 | 0.12 | 0.24 | 0.4 | 0.6 |
| C17:1 | 0.03 | 0.03 | 0.04 | nd | 0 | 0.1 |
| C18:0 | 3.51 | 2.04 | 2.99 | 5.23 | 4.8 | 6.9 |
| C18:1cis | 29.71 | 5.7 | 34 | 4.17 | 32 | 14.9 |
| C18:1trans | 0.19 | nd | 0.22 | nd | 0.9 | 1.7 |
| C18:2cis | 24.34 | 12.63 | 24.3 | 17.25 | 23.9 | 12.1 |
| C18:3 (n-3) | 3.86 | 32.51 | 3.97 | 39.1 | 4.3 | 18.5 |
| C18:3 (n-6) | 0.03 | 0.18 | 0.06 | 0.13 | . | . |
| C20:0 | 0.45 | 0.77 | 0.44 | 1.05 | 0.3 | 0.4 |
| C22:0 | 0.15 | 2.56 | 0.42 | 1.78 | 0.3 | 0.3 |
| C23:0 | nd | nd | nd | nd | 0.4 | 1.5 |
| C24:0 | 0.35 | 0.89 | 0.37 | 2 | 0.2 | 0.2 |
| Ether extract (g/100) | 3.5 | 1.6 | 4.1 | 1.5 | 3.5 | 2.4 |
| CB-TMR | CB-GRZ | OD-GRZ | p Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ML1 | ML2 | AT | ML1 | ML2 | AT | ML1 | ML2 | AT | AML1 | AML2 | T | ML | T * ML | |
| Milk yield (kg/d) | 41.3 ± 1.0 | 39.4 ± 1.0 | 40.7 ± 0.7 x | 29.9 ± 1.1 | 27.7 ± 1.1 | 28.3 ± 0.7 y | 28.7 ± 1.0 | 26.5 ± 1.0 | 27.5 ± 0.7 y | 32.9 ± 0.6 A | 31.1 ± 0.6 B | <0.001 | 0.03 | ns |
| 3.5% FCM (kg/d) | 39.5 ± 1.1 | 36.5 ± 1.1 | 38.5 ± 0.8 x | 26.8 ± 1.2 | 25.9 ± 1.2 | 26.4 ± 0.8 y | 27.4 ± 1.2 | 25.7 ± 1.2 | 26.6 ± 0.8 y | 31.4 ± 0.7 A | 29.5 ± 0.7 B | <0.001 | 0.04 | ns |
| Fat | ||||||||||||||
| (%) | 3.4 ± 0.1 | 3.2 ± 0.1 | 3.4 ± 0.1 | 3.1 ± 0.2 | 3.2 ± 0.2 | 3.1 ± 0.1 | 3.3 ± 0.2 | 3.3 ± 0.2 | 3.3 ± 0.1 | 3.3 ± 0.1 | 3.2 ± 0.1 | ns | ns | ns |
| (kg/d) | 1.4 ± 0.05 | 1.2 ± 0.05 | 1.3 ± 0.04 x | 0.9 ± 0.05 | 0.8 ± 0.05 | 0.9 ± 0.04 y | 0.9 ± 0.05 | 0.8 ± 0.05 | 0.9 ± 0.04 y | 1.05 ± 0.03 | 0.98 ± 0.03 | <0.001 | 0.07 | ns |
| CB-TMR | CB-GRZ | OD-GRZ | p Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ML1 | ML 2 | AT | ML 1 | ML 2 | AT | ML 1 | ML 2 | AT | AML 1 | A ML2 | T | ML | T * ML | |
| FA saturation (g/100 g of fat) | ||||||||||||||
| SFA | 65.7 ± 0.5 | 62.8 ± 0.5 | 64.3 ± 0.4 x | 62.4 ± 0.6 | 61.4 ± 0.6 | 61.9 ± 0.4 y | 63.4 ± 0.6 | 61.6 ± 0.6 | 62.3 ± 0.4 y | 63.7 ± 0.3 A | 61.9 ± 0.3 B | <0.001 | <0.001 | ns |
| UFA | 34.2 ± 0.5 | 37.1 ± 0.5 | 35.7 ± 0.4 y | 37.5 ± 0.6 | 38.4 ± 0.6 | 37.9 ± 0.4 x | 36.5 ± 0.6 | 38.2 ± 0.6 | 37.4 ± 0.4 x | 36.1 ± 0.3 B | 37.9 ± 0.3 A | <0.001 | <0.001 | ns |
| MUFA | 29.1 ± 0.5 | 32.3 ± 0.5 | 30.7 ± 0.3 y | 32.7 ± 0.5 | 33.7 ± 0.5 | 33.2 ± 0.4 x | 31.8 ± 0.5 | 33.6 ± 0.5 | 32.7 ± 0.4 x | 31.2 ± 0.3 B | 33.2 ± 0.3 A | <0.001 | <0.001 | ns |
| PUFA | 5.1 ± 0.1 | 4.8 ± 0.1 | 4.9 ± 0.1 | 4.8 ± 0.1 | 4.7 ± 0.1 | 4.7 ± 0.1 | 4.7 ± 0.1 | 4.6 ± 0.1 | 4.7 ± 0.1 | 4.9 ± 0.1 | 4.7 ± 0.1 | ns | 0.08 | ns |
| n-3 | 0.48 ± 0.04 bc | 0.45 ± 0.04 c | 0.46 ± 0.02 y | 0.67 ± 0.04 a | 0.61 ± 0.04 ab | 0.64 ± 0.03 x | 0.43 ± 0.04 c | 0.57 ± 0.04 abc | 0.51 ± 0.03 y | 0.53 ± 0.03 | 0.54 ± 0.03 | <0.001 | ns | 0.03 |
| n-6 | 3.4 ± 0.1 a | 2.9 ± 0.1 b | 3.1 ± 0.07 x | 2.6 ± 0.1 bc | 2.4 ± 0.1 c | 2.5 ± 0.07 y | 2.2 ± 0.1 | 2.3 ± 0.1 c | 2.2 ± 0.07 y | 2.7 ± 0.05 A | 2.5 ± 0.06 B | <0.001 | 0.03 | 0.02 |
| n6/n3 | 7.1 ± 0.3 | 6.4 ± 0.3 | 6.8 ± 0.1 x | 3.4 ± 0.3 | 4.7 ± 0.3 | 3.8 ± 0.2 y | 4.4 ± 0.3 | 3.9 ± 0.3 | 4.2 ± 0.2 y | 4.9 ± 0.2 | 4.8 ± 0.1 | <0.001 | ns | 0.08 |
| Trans | 4.1 ± 0.2 | 4.2 ± 0.2 | 4.2 ± 0.1 y | 5.1 ± 0.2 | 5.2 ± 0.2 | 5.2 ± 0.1 x | 5.5 ± 0.2 | 5.1 ± 0.2 | 5.3 ± 0.1 x | 4.9 ± 0.1 | 4.8 ± 0.1 | <0.001 | ns | ns |
| FA origin (g/100 g of fat) | ||||||||||||||
| De novo | ||||||||||||||
| (C4:0-C15:1) | 19.3 ± 0.5 | 17.2 ± 0.5 | 18.2 ± 0.3 | 19.4 ± 0.5 | 16.1 ± 0.5 | 17.7 ± 0.3 | 19.5 ± 0.5 | 15.9 ± 0.5 | 17.7 ± 0.3 | 19.4 ± 0.3 A | 16.4 ± 0.3 B | ns | <0.001 | ns |
| Mixed origin | ||||||||||||||
| (C16:0 + C16:1) | 37.6 ± 0.5 a | 37.4 ± 0.5 a | 37.5 ± 0.4 x | 33.6 ± 0.5 b | 36.6 ± 0.5 a | 35.1 ± 0.4 y | 33.2 ± 0.5 b | 36.5 ± 0.5 a | 34.8 ± 0.4 y | 34.8 ± 0.3 B | 36.8 ± 0.3 A | <0.001 | <0.001 | 0.002 |
| Preformed | ||||||||||||||
| (>C17:0) | 43.6 ± 0.7 | 45.2 ± 0.7 | 44.2 ± 0.5 x | 46.9 ± 0.7 | 47.1 ± 0.7 | 47.0 ± 0.5 y | 46.5 ± 0.7 | 47.4 ± 0.7 | 46.9 ± 0.5 y | 45.5 ± 0.7 | 46.5 ± 0.7 | <0.001 | 0.06 | ns |
| SFA/UFAratio | 1.9 ± 0.04 | 1.7 ± 0.04 | 1.8 ± 0.03 x | 1.6 ± 0.04 | 1.5 ± 0.04 | 1.6 ± 0.03 y | 1.7 ± 0.04 | 1.6 ± 0.04 | 1.6 ± 0.03 y | 1.7 ± 0.02 A | 1.6 ± 0.02 B | <0.001 | <0.001 | ns |
| Atherogenicity index (AI) | 2.2 ± 0.05 | 1.9 ± 0.05 | 2.1 ± 0.04 x | 1.9 ± 0.05 | 1.8 ± 0.05 | 1.9 ± 0.04 y | 2.1 ± 0.06 | 1.8 ± 0.06 | 1.9 ± 0.04 y | 2.1 ± 0.03 A | 1.9 ± 0.03 B | <0.001 | <0.001 | ns |
| Thrombogenicity index (TI) | 3.4 ± 0.05 | 3.0 ± 0.07 | 3.2 ± 0.05 x | 2.9 ± 0.08 | 2.8 ± 0.07 | 2.8 ± 0.05 y | 3.0 ± 0.07 | 2.9 ± 0.07 | 2.9 ± 0.05 y | 3.1 ± 0.04 A | 2.9 ± 0.04 B | <0.001 | 0.005 | ns |
| CB-TMR | CB-GRZ | OD-GRZ | p Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ML1 | ML2 | AT | ML1 | ML2 | AT | ML1 | ML2 | AT | AML1 | AML2 | T | ML | T * ML | |
| FA (g/100 g of fat) | ||||||||||||||
| C4:0 | 0.84 ± 0.03 | 0.6 ± 0.03 | 0.72 ± 0.02 x | 0.79 ± 0.03 | 0.52 ± 0.03 | 0.65 ± 0.02 xy | 0.7 ± 0.03 | 0.52 ± 0.03 | 0.61 ± 0.02 y | 0.77 ± 0.02 A | 0.55 ± 0.02 B | 0.003 | <0.001 | ns |
| C6:0 | 0.94 ± 0.03 | 0.73 ± 0.03 | 0.83 ± 0.02 x | 0.85 ± 0.03 | 0.67 ± 0.03 | 0.76 ± 0.02 y | 0.84 ± 0.03 | 0.61 ± 0.03 | 0.72 ± 0.02 y | 0.88 ± 0.01 A | 0.67 ± 0.01 B | 0.001 | <0.001 | ns |
| C8:0 | 0.77 ± 0.02 | 0.6 ± 0.02 | 0.68 ± 0.02 x | 0.70 ± 0.03 | 0.52 ± 0.03 | 0.68 ± 0.02 y | 0.66 ± 0.03 | 0.47 ± 0.03 | 0.57 ± 0.002 y | 0.71 ± 0.02 A | 0.53 ± 0.01 B | <0.001 | <0.001 | ns |
| C10:0 | 2.2 ± 0.07 | 1.7 ± 0.02 | 1.2 ± 0.05 x | 1.9 ± 0.08 | 1.4 ± 0.08 | 1.7 ± 0.06 y | 1.7 ± 0.08 | 1.3 ± 0.08 | 1.5 ± 0.06 y | 1.9 ± 0.04 A | 1.5 ± 0.04 B | <0.001 | <0.001 | ns |
| C12:0 | 2.5 ± 0.08 | 2.2 ± 0.08 | 2.4 ± 0.05 x | 2.3 ± 0.08 | 1.8 ± 0.08 | 2.1 ± 0.06 y | 2 ± 0.08 | 1.8 ± 0.08 | 1.9 ± 0.06 y | 2.3 ± 0.04 A | 1.9 ± 0.04 B | <0.001 | <0.001 | ns |
| C14:0 | 9.7 ± 0.3 b | 9.0 ± 0.2 bc | 9.3 ± 0.25 | 9.7 ± 0.3 b | 8.5 ± 0.2 c | 9.1 ± 0.26 | 10.8 ± 0.3 a | 8.4 ± 0.2 c | 9.6 ± 0.3 | 10.3 ± 0.2 A | 8.26 ± 0.2 B | ns | <0.001 | 0.005 |
| C14:1 cis | 0.63 ± 0.05 b | 0.81 ± 0.05 ab | 0.72 ± 0.03 | 0.88 ± 0.06 a | 0.76 ± 0.06 ab | 0.82 ± 0.04 | 0.64 ± 0.06 ab | 0.76 ± 0.06 ab | 0.72 ± 0.04 | 0.72 ± 0.03 | 0.78 ± 0.03 | 0.08 | ns | 0.03 |
| C15:0 | 1.2 ± 0.04 | 1.2 ± 0.04 | 1.2 ± 0.03 y | 1.9 ± 0.04 | 1.5 ± 0.04 | 1.7 ± 0.03 x | 1.7 ± 0.04 | 1.6 ± 0.04 | 1.7 ± 0.03 x | 1.6 ± 0.02 A | 1.46 ± 0.02 B | <0.001 | <0.001 | ns |
| C16:0 | 35.7 ± 0.5 a | 35.3 ± 0.5 a | 35.5 ± 0.35 x | 31.2 ± 0.5 b | 34.3 ± 0.5 a | 32.7 ± 0.38 y | 31.3 ± 0.5 b | 34.1 ± 0.5 a | 32.7 ± 0.29 y | 32.7 ± 0.3 B | 34.6 ± 0.3 A | <0.001 | <0.001 | 0.001 |
| C16:1 cis | 1.6 ± 0.08 ab | 1.8 ± 0.08 a | 1.7 ± 0.06 | 1.8 ± 0.09 a | 1.8 ± 0.09 a | 1.8 ± 0.06 | 1.3 ± 0.09 a | 1.9 ± 0.09 a | 1.6 ± 0.06 | 1.3 ± 0.05 B | 1.9 ± 0.05 A | ns | <0.001 | 0.001 |
| C16:1 trans | 0.33 ± 0.03 d | 0.35 ± 0.03 cd | 0.35 ± 0.02 z | 0.51 ± 0.03 b | 0.44 ± 0.03 bcd | 0.47 ± 0.02 y | 0.75 ± 0.03 a | 0.48 ± 0.03 bc | 0.62 ± 0.02 x | 0.53 ± 0.02 A | 0.43 ± 0.02 B | <0.001 | <0.001 | <0.001 |
| C18:0 | 10.6 ± 0.43 | 10.3 ± 0.43 | 10.5 ± 0.31 | 11.5 ± 0.45 | 10.6 ± 0.45 | 11 ± 0.32 | 11.5 ± 0.45 | 11.2 ± 0.45 | 11.4 ± 0.32 | 11.2 ± 0.2 | 10.7 ± 0.02 | ns | ns | ns |
| C18:1 cis | 22.8 ± 0.43 b | 25.6 ± 0.43 a | 24.2 ± 0.31 | 24.7 ± 0.45 a | 25.7 ± 0.45 a | 25.2 ± 0.32 | 24.6 ± 0.45 a | 25.7 ± 0.45 a | 25.2 ± 0.32 | 24.1 ± 0.3 B | 25.7 ± 0.3 A | 0.03 | <0.001 | 0.07 |
| C18:1 trans | 3.3 ± 0.15 | 3.3 ± 0.15 | 3.3 ± 0.11 y | 4.1 ± 0.16 | 4.4 ± 0.16 | 4.2 ± 0.11 x | 4 ± 0.16 | 4 ± 0.16 | 4 ± 0.11 x | 3.7 ± 0.09 | 3.9 ± 0.09 | <0.001 | ns | ns |
| C18:2 CLA | 0.73 ± 0.06 c | 0.82 ± 0.06 bc | 0.77 ± 0.04 y | 1.0 ± 0.06 ab | 1.15 ± 0.06 a | 1.1 ± 0.04 x | 1.2 ± 0.06 a | 1.1 ± 0.06 ab | 1.12 ± 0.04 x | 0.98 ± 0.03 | 1.0 ± 0.03 | <0.001 | ns | 0.04 |
| C18:3 (n-3) | 0.26 ± 0.03 | 0.24 ± 0.03 | 0.25 ± 0.02 z | 0.53 ± 0.03 | 0.46 ± 0.03 | 0.49 ± 0.02 x | 0.39 ± 0.03 | 0.36 ± 0.03 | 0.38 ± 0.02 y | 0.39 ± 0.02 | 0.35 ± 0.02 | <0.001 | ns | ns |
| MilkP | Cheese | |||||||
|---|---|---|---|---|---|---|---|---|
| CB-TMR | CB-GRZ | OD-GRZ | p Value | CB-TMR | CB-GRZ | OD-GRZ | p Value | |
| T | T | |||||||
| FA saturation (g/100 g of fat) | ||||||||
| SFA | 63.5 ± 0.9 | 64.5 ± 0.9 | 64.1 ± 0.9 | ns | 64.2 ± 0.5 | 64.4 ± 0.5 | 64.7 ± 0.5 | ns |
| MUFA | 32.3 ± 0.7 | 31.2 ± 0.7 | 31.6 ± 0.7 | ns | 31.8 ± 0.5 | 31.5 ± 0.5 | 31.3 ± 0.5 | ns |
| PUFA | 4.0 ± 0.2 | 3.0 ± 0.2 | 4.1 ± 0.2 | ns | 3.8 ± 0.1 | 3.8 ± 0.1 | 3.8 ± 0.1 | ns |
| n-3 | 0.26 ± 0.03 x | 0.51 ± 0.03 y | 0.50 ± 0.03 y | <0.001 | 0.43 ± 0.03 x | 0.55 ± 0.03 y | 0.46 ± 0.03 xy | <0.001 |
| n-6 | 2.5 ± 0.1 x | 1.9 ± 0.1 y | 1.85 ± 0.1 y | <0.001 | 2.9 ± 0.1 x | 2.6 ± 0.1 y | 2.3 ± 0.1 y | <0.001 |
| n6/n3 | 9.5 ± 0.1 x | 3.7 ± 0.1 y | 3.7 ± 0.1 y | <0.001 | 7.1 ± 0.5 x | 4.8 ± 0.5 y | 5.2 ± 0.5 y | <0.001 |
| Trans | 3.5 ± 0.1 | 4.2 ± 0.1 | 2.0 ± 0.1 | <0.001 | 3.6 ± 0.1 | 3.8 ± 0.1 | 3.9 ± 0.2 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grille, L.; Escobar, D.; Méndez, M.N.; Adrien, M.d.L.; Olazabal, L.; Rodríguez, V.; Pelaggio, R.; Chilibroste, P.; Meikle, A.; Damián, J.P. Different Conditions during Confinement in Pasture-Based Systems and Feeding Systems Affect the Fatty Acid Profile in the Milk and Cheese of Holstein Dairy Cows. Animals 2023, 13, 1426. https://doi.org/10.3390/ani13081426
Grille L, Escobar D, Méndez MN, Adrien MdL, Olazabal L, Rodríguez V, Pelaggio R, Chilibroste P, Meikle A, Damián JP. Different Conditions during Confinement in Pasture-Based Systems and Feeding Systems Affect the Fatty Acid Profile in the Milk and Cheese of Holstein Dairy Cows. Animals. 2023; 13(8):1426. https://doi.org/10.3390/ani13081426
Chicago/Turabian StyleGrille, Lucía, Daniela Escobar, Maria Noel Méndez, María de Lourdes Adrien, Laura Olazabal, Víctor Rodríguez, Ronny Pelaggio, Pablo Chilibroste, Ana Meikle, and Juan Pablo Damián. 2023. "Different Conditions during Confinement in Pasture-Based Systems and Feeding Systems Affect the Fatty Acid Profile in the Milk and Cheese of Holstein Dairy Cows" Animals 13, no. 8: 1426. https://doi.org/10.3390/ani13081426
APA StyleGrille, L., Escobar, D., Méndez, M. N., Adrien, M. d. L., Olazabal, L., Rodríguez, V., Pelaggio, R., Chilibroste, P., Meikle, A., & Damián, J. P. (2023). Different Conditions during Confinement in Pasture-Based Systems and Feeding Systems Affect the Fatty Acid Profile in the Milk and Cheese of Holstein Dairy Cows. Animals, 13(8), 1426. https://doi.org/10.3390/ani13081426