
A Coupled Human and Natural Systems Framework to Characterize Emerging Infectious Diseases—The Case of Fibropapillomatosis in Marine Turtles

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Emerging Wildlife Diseases in Coupled Human and Natural Systems
1.1. Increased Movement: Travel and Products Trade
1.2. Anthropogenic Presence: Urbanization and Human Encroachment
1.3. Animal Use: Wildlife Trade and Markets, Unsustainable Farming, Ecotourism, Bushmeat
1.4. Environmental Alterations: Climate Change and Pollution
2. The Case of Sea Turtle Fibropapillomatosis
2.1. Coastal Coupled Human and Natural Systems
2.2. Human Encroachment and Urbanization: Anthropogenic Presence and Sea Turtle Health
2.3. Pollution: Role of Water Quality in Sea Turtle Disease
2.4. Climate Change: Effect of Fluctuating Temperatures on Coastal Ecosystems
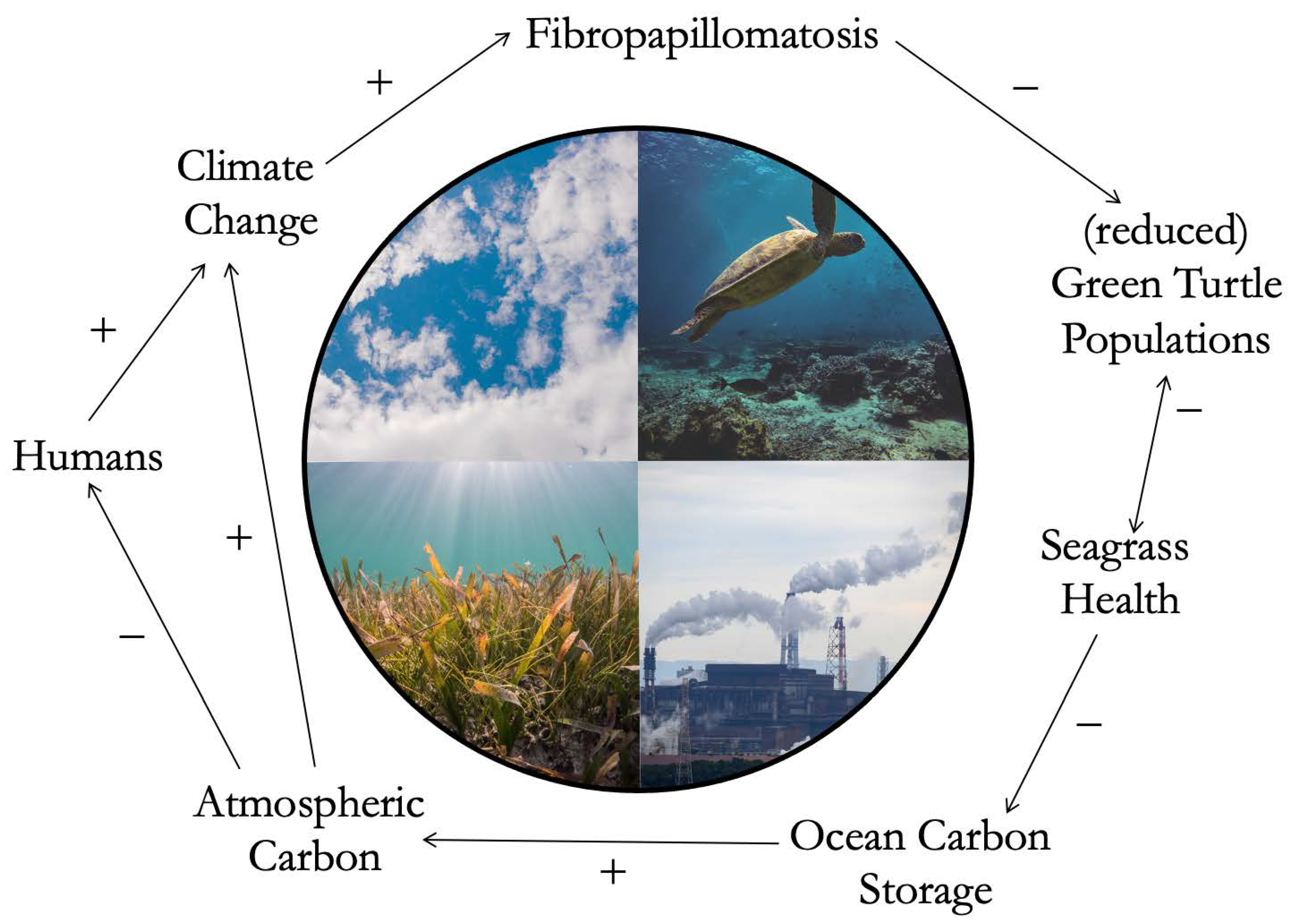
3. The CHANS Novelty of Recognizing Feedback Loops
4. Importance of One Health and Multifocal Interventions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunningham, A.A.; Daszak, P.; Wood, J.L.N. One Health, Emerging Infectious Diseases and Wildlife: Two Decades of Progress? Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160167. [Google Scholar] [CrossRef]
- Tompkins, D.M.; Carver, S.; Jones, M.E.; Krkošek, M.; Skerratt, L.F. Emerging Infectious Diseases of Wildlife: A Critical Perspective. Trends Parasitol. 2015, 31, 149–159. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic Environmental Change and the Emergence of Infectious Diseases in Wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Kramer, D.B.; Hartter, J.; Boag, A.E.; Jain, M.; Stevens, K.; Nicholas, K.A.; McConnell, W.J.; Liu, J. Top 40 Questions in Coupled Human and Natural Systems (CHANS) Research. Ecol. Soc. 2017, 22, 44. [Google Scholar] [CrossRef]
- Liu, J.; Dietz, T.; Carpenter, S.R.; Alberti, M.; Folke, C.; Moran, E.; Pell, A.N.; Deadman, P.; Kratz, T.; Lubchenco, J.; et al. Complexity of Coupled Human and Natural Systems. Science 2007, 317, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Thirgood, S. New Perspectives on Managing Wildlife Diseases. J. Appl. Ecol. 2008, 45, 1321–1329. [Google Scholar] [CrossRef]
- Wasik, B.R.; De Wit, E.; Munster, V.; Lloyd-Smith, J.O.; Martinez-Sobrido, L.; Parrish, C.R. Onward Transmission of Viruses: How Do Viruses Emerge to Cause Epidemics after Spillover? Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190017. [Google Scholar] [CrossRef]
- Wilson, M.E. Travel and the Emergence of Infectious Diseases. J. Agromed. 2005, 9, 159–177. [Google Scholar] [CrossRef]
- Findlater, A.; Bogoch, I.I. Human Mobility and the Global Spread of Infectious Diseases: A Focus on Air Travel. Trends Parasitol. 2018, 34, 772–783. [Google Scholar] [CrossRef]
- Tuite, A.R.; Bhatia, D.; Moineddin, R.; Bogoch, I.I.; Watts, A.G.; Khan, K. Global Trends in Air Travel: Implications for Connectivity and Resilience to Infectious Disease Threats. J. Travel Med. 2020, 27, taaa070. [Google Scholar] [CrossRef]
- Chase-Dunn, C.; Kawano, Y.; Brewer, B.D. Trade Globalization since 1795: Waves of Integration in the World-System. Am. Sociol. Rev. 2016, 65, 77–95. [Google Scholar] [CrossRef]
- Nentwig, W. Introduction, Establishment Rate, Pathways and Impact of Spiders Alien to Europe. Biol. Invasions 2015, 17, 2757–2778. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, Transport and Trouble: Managing Invasive Species Pathways in an Era of Globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Frick, W.F.; Pollock, J.F.; Hicks, A.C.; Langwig, K.E.; Reynolds, D.S.; Turner, G.G.; Butchkoski, C.M.; Kunz, T.H. An Emerging Disease Causes Regional Population Collapse of a Common North American Bat Species. Science 2010, 329, 679–682. [Google Scholar] [CrossRef]
- Perrings, C.; Levin, S.; Daszak, P. The Economics of Infectious Disease, Trade and Pandemic Risk. Ecohealth 2018, 15, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Davis, K. The Urbanization of the Human Population. Sci. Am. 1965, 5, 248–253. [Google Scholar] [CrossRef]
- Zúñiga-Sarango, W.; Gaona, F.P.; Reyes-Castillo, V.; Iñiguez-Armijos, C. Disrupting the Biodiversity–Ecosystem Function Relationship: Response of Shredders and Leaf Breakdown to Urbanization in Andean Streams. Front. Ecol. Evol. 2020, 8, 592404. [Google Scholar] [CrossRef]
- Cohen, B. Urbanization in Developing Countries: Current Trends, Future Projections, and Key Challenges for Sustainability. Technol. Soc. 2006, 28, 63–80. [Google Scholar] [CrossRef]
- Ren, Q.; He, C.; Huang, Q.; Zhang, D.; Shi, P.; Lu, W. Impacts of Global Urban Expansion on Natural Habitats Undermine the 2050 Vision for Biodiversity. Resour. Conserv. Recycl. 2023, 190, 106834. [Google Scholar] [CrossRef]
- Neiderud, C.J. How Urbanization Affects the Epidemiology of Emerging Infectious Diseases. Afr. J. Disabil. 2015, 5, 27060. [Google Scholar] [CrossRef]
- Tong, M.X.; Hansen, A.; Hanson-Easey, S.; Cameron, S.; Xiang, J.; Liu, Q.; Sun, Y.; Weinstein, P.; Han, G.S.; Williams, C.; et al. Infectious Diseases, Urbanization and Climate Change: Challenges in Future China. Int. J. Environ. Res. Public Health 2015, 12, 11025–11036. [Google Scholar] [CrossRef]
- Combs, M.A.; Kache, P.A.; VanAcker, M.C.; Gregory, N.; Plimpton, L.D.; Tufts, D.M.; Fernandez, M.P.; Diuk-Wasser, M.A. Socio-Ecological Drivers of Multiple Zoonotic Hazards in Highly Urbanized Cities. Glob. Change Biol. 2022, 28, 1705–1724. [Google Scholar] [CrossRef]
- Wilkinson, D.A.; Marshall, J.C.; French, N.P.; Hayman, D.T.S. Habitat Fragmentation, Biodiversity Loss and the Risk of Novel Infectious Disease Emergence. J. R. Soc. Interface 2018, 15, 20180403. [Google Scholar] [CrossRef] [PubMed]
- Pongsiri, M.J.; Roman, J.; Ezenwa, V.O.; Goldberg, T.L.; Koren, H.S.; Newbold, S.C.; Ostfeld, R.S.; Pattanayak, S.K.; Salkeld, D.J. Biodiversity Loss Affects Global Disease Ecology. Bioscience 2009, 59, 945–954. [Google Scholar] [CrossRef]
- McKee, C.D.; Islam, A.; Luby, S.P.; Salje, H.; Hudson, P.J.; Plowright, R.K.; Gurley, E.S. The Ecology of Nipah Virus in Bangladesh: A Nexus of Land-Use Change and Opportunistic Feeding Behavior in Bats. Viruses 2021, 13, 169. [Google Scholar] [CrossRef]
- Peterson, P.K.; Chao, C.C.; Molitor, T.; Murtaugh, M.; Strgar, F.; Sharp, B.M. Stress and Pathogenesis of Infectious Disease. Rev. Infect. Dis. 1991, 13, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.H.; Sánchez, C.A.; Becker, D.J.; Byers, K.A.; Worsley-Tonks, K.E.L.; Craft, M.E. City Sicker? A Meta-Analysis of Wildlife Health and Urbanization. Front. Ecol. Environ. 2019, 17, 575–583. [Google Scholar] [CrossRef]
- Kaisin, O.; Fuzessy, L.; Poncin, P.; Brotcorne, F.; Culot, L. A Meta-Analysis of Anthropogenic Impacts on Physiological Stress in Wild Primates. Conserv. Biol. 2021, 35, 101–114. [Google Scholar] [CrossRef]
- Ditchkoff, S.S.; Saalfeld, S.T.; Gibson, C.J. Animal Behavior in Urban Ecosystems: Modifications Due to Human-Induced Stress. Urban Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Marano, N.; Arguin, P.M.; Pappaioanou, M. Impact of Globalization and Animal Trade on Infectious Disease Ecology. Emerg. Infect. Dis. 2007, 13, 1807–1809. [Google Scholar] [CrossRef]
- Fèvre, E.M.; Bronsvoort, B.M.D.C.; Hamilton, K.A.; Cleaveland, S. Animal Movements and the Spread of Infectious Diseases. Trends Microbiol. 2006, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- McMurray, C.A. Wildlife Trafficking: U.S. Efforts to Tackle a Global Crisis. Nat. Resour. Environ. 2014, 23, 16–18. [Google Scholar]
- Schloegel, L.M.; Daszak, P.; Cunningham, A.A.; Speare, R.; Hill, B. Two Amphibian Diseases, Chytridiomycosis and Ranaviral Disease, Are Now Globally Notifiable to the World Organization for Animal Health (OIE): An Assessment. Dis. Aquat. Organ. 2010, 92, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.Y.; Lau, S.K.P.; Yuen, K.Y. Infectious Diseases Emerging from Chinese Wet-Markets: Zoonotic Origins of Severe Respiratory Viral Infections. Curr. Opin. Infect. Dis. 2006, 19, 401–407. [Google Scholar] [CrossRef]
- Webster, R.G. Wet Markets—A Continuing Source of Severe Acute Respiratory Syndrome and Influenza? Lancet 2004, 363, 234–236. [Google Scholar] [CrossRef]
- Espinosa, R.; Tago, D.; Treich, N. Infectious Diseases and Meat Production. Environ. Resour. Econ. 2020, 76, 1019–1044. [Google Scholar] [CrossRef]
- Whitfort, A. COVID-19 and Wildlife Farming in China: Legislating to Protect Wild Animal Health and Welfare in the Wake of a Global Pandemic. J. Environ. Law 2021, 33, 57–84. [Google Scholar] [CrossRef]
- Ostrowski, S.; Madad, R.A.; Hafizullah, N. Controlling the Risk of Spill over of Peste Des Petits Ruminants Virus from Livestock to Threatened Wild Ungulates in Wakhan National Park, Afghanistan. In Proceedings of the Conservation Asia 2018, Bishkek, Kyrgyzstan, 6–10 August 2018; pp. 1–18. [Google Scholar]
- Muehlenbein, M.P.; Wallis, J. Considering Risks of Pathogen Transmission Associated with Primate-Based Tourism. Primate Tour. Tool Conserv., 1st Edition; Cambridge University Press: Cambridge, UK, 2014; Chapter 16; pp. 278–291. ISBN 9781107018129. [Google Scholar]
- Wolfe, N.D.; Daszak, P.; Kilpatrick, A.M.; Burke, D.S. Bushmeat Hunting, Deforestation, and Prediction of Zoonotic Disease Emergence. Emerg. Infect. Dis. 2005, 11, 1822–1827. [Google Scholar] [CrossRef]
- Karesh, W.B.; Noble, E. The Bushmeat Trade: Increased Opportunities for Transmission of Zoonotic Disease. Medicine 2007, 76, 15–21. [Google Scholar] [CrossRef]
- van Velden, J.; Wilson, K.; Biggs, D. The Evidence for the Bushmeat Crisis in African Savannas: A Systematic Quantitative Literature Review. Biol. Conserv. 2018, 221, 345–356. [Google Scholar] [CrossRef]
- Pourrut, X.; Kumulungui, B.; Wittmann, T.; Moussavou, G.; Délicat, A.; Yaba, P.; Nkoghe, D.; Gonzalez, J.P.; Leroy, E.M. The Natural History of Ebola Virus in Africa. Microbes Infect. 2005, 7, 1005–1014. [Google Scholar] [CrossRef]
- Leroy, E.M.; Epelboin, A.; Mondonge, V.; Pourrut, X.; Gonzalez, J.P.; Muyembe-Tamfum, J.J.; Formenty, P. Human Ebola Outbreak Resulting from Direct Exposure to Fruit Bats in Luebo, Democratic Republic of Congo, 2007. Vector-Borne Zoonotic Dis. 2009, 9, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.; Cui, X.; Gillson, L.; Nelson, D.; Taylor, M.P.; Vanacker, V.; Wang, E. Anthropocene in an Age of Pandemics. Anthropocene 2020, 30, 100247. [Google Scholar] [CrossRef]
- Lafferty, K.D. The Ecology of Climate Change and Infectious Diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef]
- Kock, R.A.; Orynbayev, M.; Robinson, S.; Zuther, S.; Singh, N.J.; Beauvais, W.; Morgan, E.R.; Kerimbayev, A.; Khomenko, S.; Martineau, H.M.; et al. Saigas on the Brink: Multidisciplinary Analysis of the Factors Influencing Mass Mortality Events. Sci. Adv. 2018, 4, eaao2314. [Google Scholar] [CrossRef] [PubMed]
- Epstein, P.R. Climate Change and Infectious Disease: Stormy Weather Ahead? Epidemiology 2002, 13, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, E.B.; Peel, A.J.; Reid, S.A.; Jansen, C.C.; McCallum, H. The Non-Human Reservoirs of Ross River Virus: A Systematic Review of the Evidence. Parasites Vectors 2018, 11, 188. [Google Scholar] [CrossRef]
- Martin, L.B.; Hopkins, W.A.; Mydlarz, L.D.; Rohr, J.R. The Effects of Anthropogenic Global Changes on Immune Functions and Disease Resistance. Ann. N. Y. Acad. Sci. 2010, 1195, 129–148. [Google Scholar] [CrossRef] [PubMed]
- Bosi, S.; Desmarchelier, D. Pollution and Infectious Diseases. Int. J. Econ. Theory 2018, 14, 351–372. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging Infectious Diseases of Plants: Pathogen Pollution, Climate Change and Agrotechnology Drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Morley, N.J. Interactive Effects of Infectious Diseases and Pollution in Aquatic Molluscs. Aquat. Toxicol. 2010, 96, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hidaka, T.; Kumagai, Y.; Yamamoto, M. Environmental Pollutants and the Immune Response. Nat. Immunol. 2020, 21, 1486–1495. [Google Scholar] [CrossRef]
- Wilson, S.C.; Eybatov, T.M.; Amano, M.; Jepson, P.D.; Goodman, S.J. The Role of Canine Distemper Virus and Persistent Organic Pollutants in Mortality Patterns of Caspian SEALS (Pusa caspica). PLoS ONE 2014, 9, e99265. [Google Scholar] [CrossRef]
- Browning, H.M.; Gulland, F.M.D.; Hammond, J.A.; Colegrove, K.M.; Hall, A.J. Common Cancer in a Wild Animal: The California Sea Lion (Zalophus californianus) as an Emerging Model for Carcinogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140228. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.M.; Coates, C.W. Fibro-Epithelial Growths of the Skin in Large Marine Turtles, Chelonia mydas (Linnaeus). Zoologica 1938, 23, 93–98. [Google Scholar] [CrossRef]
- Perrault, J.R.; Levin, M.; Mott, C.R.; Bovery, C.M.; Bresette, M.J.; Chabot, R.M.; Gregory, C.R.; Guertin, J.R.; Hirsch, S.E.; Ritchie, B.W.; et al. Insights on Immune Function in Free-Ranging Green Sea Turtles (Chelonia mydas) with and without Fibropapillomatosis. Animals 2021, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- Herbst, L.H.; Jacobson, E.R.; Moretti, R.; Brown, T.; Sundberg, J.P.; Klein, P.A. Experimental Transmission of Green Turtle Fibropapillomatosis Using Cell-Free Tumor Extracts. Dis. Aquat. Organ. 1995, 22, 1–12. [Google Scholar] [CrossRef]
- Alfaro-Núñez, A.; Bertelsen, M.F.; Bojesen, A.M.; Rasmussen, I.; Zepeda-Mendoza, L.; Olsen, M.T.; Gilbert, M.T.P. Global Distribution of Chelonid Fibropapilloma-Associated Herpesvirus among Clinically Healthy Sea Turtles. BMC Evol. Biol. 2014, 14, 206. [Google Scholar] [CrossRef]
- Greenblatt, R.J.; Work, T.M.; Dutton, P.; Sutton, C.A.; Spraker, T.R.; Casey, R.N.; Diez, C.E.; Parker, D.; St. Leger, J.; Balazs, G.H.; et al. Geographic Variation in Marine Turtle Fibropapillomatosis. J. Zoo Wildl. Med. 2005, 36, 527–530. [Google Scholar] [CrossRef]
- Morrison, C.L.; Iwanowicz, L.; Work, T.M.; Fahsbender, E.; Breitbart, M.; Adams, C.; Iwanowicz, D.; Sanders, L.; Ackermann, M.; Cornman, R.S. Genomic Evolution, Recombination, and Inter-Strain Diversity of Chelonid Alphaherpesvirus 5 from Florida and Hawaii Green Sea Turtles with Fibropapillomatosis. PeerJ 2018, 6, e4386. [Google Scholar] [CrossRef]
- Page-Karjian, A.; Norton, T.M.; Ritchie, B.; Brown, C.; Mancia, C.; Jackwood, M.; Gottdenker, N.L. Quantifying Chelonid Herpesvirus 5 in Symptomatic and Asymptomatic Rehabilitating Green Sea Turtles. Endanger. Species Res. 2015, 28, 135–146. [Google Scholar] [CrossRef]
- Jones, K.; Ariel, E.; Burgess, G.; Read, M. A Review of Fibropapillomatosis in Green Turtles (Chelonia mydas). Vet. J. 2016, 212, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Herbst, L.; Ene, A.; Su, M.; Desalle, R.; Lenz, J. Tumor Outbreaks in Marine Turtles Are Not Due to Recent Herpesvirus Mutations. Curr. Biol. 2004, 14, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Manes, C.; Pinton, D.; Canestrelli, A. Occurrence of Fibropapillomatosis in Green Turtles (Chelonia mydas) in Relation to Environmental Changes in Coastal Ecosystems in Texas and Florida: A Retrospective Study. Animals 2022, 12, 1236. [Google Scholar] [CrossRef]
- Hazel, J.; Lawler, I.R.; Hamann, M. Diving at the Shallow End: Green Turtle Behaviour in near-Shore Foraging Habitat. J. Exp. Mar. Bio. Ecol. 2009, 371, 84–92. [Google Scholar] [CrossRef]
- Martin, K.R.; Mansfield, K.L.; Savage, A.E. Adaptive Evolution of Major Histocompatibility Complex Class i Immune Genes and Disease Associations in Coastal Juvenile Sea Turtles. R. Soc. Open Sci. 2022, 9, 211190. [Google Scholar] [CrossRef]
- Williams, B.; Watson, J.; Angeles, L.; Runting, R.K.; Roberson, L.A. The Global Rarity of Intact Coastal Regions. bioRxiv 2021, 36, e13874. [Google Scholar] [CrossRef]
- Luijendijk, A.; Hagenaars, G.; Ranasinghe, R.; Baart, F.; Donchyts, G.; Aarninkhof, S. The State of the World’s Beaches. Sci. Rep. 2018, 8, 6641. [Google Scholar] [CrossRef]
- Malvarez, G.; Ferreira, O.; Navas, F.; Cooper, J.A.G.; Gracia-Prieto, F.J.; Talavera, L. Storm Impacts on a Coupled Human-Natural Coastal System: Resilience of Developed Coasts. Sci. Total Environ. 2021, 768, 144987. [Google Scholar] [CrossRef]
- Lester, L.A.; Avery, H.W.; Harrison, A.S.; Standora, E.A. Recreational Boats and Turtles: Behavioral Mismatches Result in High Rates of Injury. PLoS ONE 2013, 8, e82370. [Google Scholar] [CrossRef]
- Hing, S.; Narayan, E.J.; Thompson, R.C.A.; Godfrey, S.S. The Relationship between Physiological Stress and Wildlife Disease: Consequences for Health and Conservation. Wildl. Res. 2016, 43, 51–60. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Martins, A.S.; Torezani, E.; Baptistotte, C.; Da Nóbrega Farias, J.; Horta, P.A.; Work, T.M.; Balazs, G.H. Relationship between Fibropapillomatosis and Environmental Quality: A Case Study with Chelonia mydas off Brazil. Dis. Aquat. Organ. 2010, 89, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Hirama, S.; Ehrhart, L.M.; Rea, L.D.; Kiltie, R.A. Relating Fibropapilloma Tumor Severity to Blood Parameters in Green Turtles Chelonia mydas. Dis. Aquat. Organ. 2014, 111, 61–68. [Google Scholar] [CrossRef]
- Keller, J.M.; Balazs, G.H.; Nilsen, F.; Rice, M.; Work, T.M.; Jensen, B.A. Investigating the Potential Role of Persistent Organic Pollutants in Hawaiian Green Sea Turtle Fibropapillomatosis. Environ. Sci. Technol. 2014, 48, 7807–7816. [Google Scholar] [CrossRef] [PubMed]
- Patrício, A.R.; Diez, C.E.; Van Dam, R.P.; Godley, B.J. Novel Insights into the Dynamics of Green Turtle Fibropapillomatosis. Mar. Ecol. Prog. Ser. 2016, 547, 247–255. [Google Scholar] [CrossRef]
- Villa, C.A.; Flint, M.; Bell, I.; Hof, C.; Limpus, C.J.; Gaus, C. Trace Element Reference Intervals in the Blood of Healthy Green Sea Turtles to Evaluate Exposure of Coastal Populations. Environ. Pollut. 2017, 220, 1465–1476. [Google Scholar] [CrossRef]
- Farrell, J.; Thomas, R.; Martindale, M.Q.; David, J. Characterisation of Fibropapillomatosis Tumour Growth Profiles in Green Sea Turtles (Chelonia mydas). Testudo 2018, 8, 12–29. [Google Scholar]
- Luck, G.W. A Review of the Relationships between Human Population Density and Biodiversity. Biol. Rev. 2007, 82, 607–645. [Google Scholar] [CrossRef]
- McKee, J.K.; Sciulli, P.W.; David Fooce, C.; Waite, T.A. Forecasting Global Biodiversity Threats Associated with Human Population Growth. Biol. Conserv. 2004, 115, 161–164. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Bastos, K.V.; Machado, L.P.; Joyeux, J.C.; Ferreira, J.S.; Militão, F.P.; de Oliveira Fernandes, V.; Santos, R.G. Coastal Degradation Impacts on Green Turtle’s (Chelonia mydas) Diet in Southeastern Brazil: Nutritional Richness and Health. Sci. Total Environ. 2022, 823, 153593. [Google Scholar] [CrossRef] [PubMed]
- Hirama, S.; Ehrhart, L.M. Description, Prevalence and Severity of Green Turtle Fibropapillomatosis in Three Developmental Habitats on the East Coast of Florida. Fla. Sci. 2007, 70, 435–448. [Google Scholar]
- Aguirre, A.A. Green Turtle Fibropapilloma: An Epidemiologic Perspective. NOAA Tech. Memo. NMFS 1991, 156, 107–113. [Google Scholar]
- Wetz, J.J.; Lipp, E.K.; Griffin, D.W.; Lukasik, J.; Wait, D.; Sobsey, M.D.; Scott, T.M.; Rose, J.B. Presence, Infectivity, and Stability of Enteric Viruses in Seawater: Relationship to Marine Water Quality in the Florida Keys. Mar. Pollut. Bull. 2004, 48, 698–704. [Google Scholar] [CrossRef]
- Van Houtan, K.S.; Smith, C.M.; Dailer, M.L.; Kawachi, M. Eutrophication and the Dietary Promotion of Sea Turtle Tumors. PeerJ 2014, 2, e602. [Google Scholar] [CrossRef]
- da Silva, C.C.; Klein, R.D.; Barcarolli, I.F.; Bianchini, A. Metal Contamination as a Possible Etiology of Fibropapillomatosis in Juvenile Female Green Sea Turtles Chelonia mydas from the Southern Atlantic Ocean. Aquat. Toxicol. 2016, 170, 42–51. [Google Scholar] [CrossRef]
- Baines, C.; Lerebours, A.; Thomas, F.; Fort, J.; Kreitsberg, R.; Gentes, S.; Meitern, R.; Saks, L.; Ujvari, B.; Giraudeau, M.; et al. Linking Pollution and Cancer in Aquatic Environments: A Review. Environ. Int. 2021, 149, 106391. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sarmiento, A.M.; Rossi, S.; Vilca, F.Z.; Thijl Vanstreels, R.E.; Monteiro, S.H.; Vale, L.A.S.; Dos Santos, R.G.; Marigo, J.; Bertozzi, C.P.; Grisi Filho, J.H.H.; et al. Organochlorine Pesticides in Green Sea Turtles (Chelonia mydas) with and without Fibropapillomatosis Caught at Three Feeding Areas off Brazil. J. Mar. Biol. Assoc. UK 2017, 97, 215–223. [Google Scholar] [CrossRef]
- Yan, M.; Nie, H.; Wang, W.; Huang, Y.; Li, Q.X.; Wang, J. The Risk of Polychlorinated Biphenyls Facilitating Tumors in Hawaiian Green Sea Turtles (Chelonia mydas). Int. J. Environ. Res. Public Health 2018, 15, 1243. [Google Scholar] [CrossRef]
- Broderick, A.C.; Coyne, M.S.; Fuller, W.J.; Glen, F.; Godley, B.J. Fidelity and Over-Wintering of Sea Turtles. Proc. R. Soc. B Biol. Sci. 2007, 274, 1533–1538. [Google Scholar] [CrossRef]
- Tucker, A.D.; MacDonald, B.D.; Seminoff, J.A. Foraging Site Fidelity and Stable Isotope Values of Loggerhead Turtles Tracked in the Gulf of Mexico and Northwest Caribbean. Mar. Ecol. Prog. Ser. 2014, 502, 267–279. [Google Scholar] [CrossRef]
- Palaniappan, P.; Hamid, H.H.A. Spatial Site Fidelity of Sea Turtles at a Foraging Ground in Mabul Island, Sabah. Int. J. Fish. Aquat. Stud. 2017, 5, 140–144. [Google Scholar]
- Siegwalt, F.; Benhamou, S.; Girondot, M.; Jeantet, L.; Martin, J.; Bonola, M.; Lelong, P.; Grand, C.; Chambault, P.; Benhalilou, A.; et al. High Fidelity of Sea Turtles to Their Foraging Grounds Revealed by Satellite Tracking and Capture-Mark-Recapture: New Insights for the Establishment of Key Marine Conservation Areas. Biol. Conserv. 2020, 250, 108742. [Google Scholar] [CrossRef]
- Avens, L.; Goshe, L.R.; Harms, C.A.; Anderson, E.T.; Hall, A.G.; Cluse, W.M.; Godfrey, M.H.; McNeill, J.B.; Stacy, B.; Bailey, R.; et al. Population Characteristics, Age Structure, and Growth Dynamics of Neritic Juvenile Green Turtles in the Northeastern Gulf of Mexico. Mar. Ecol. Prog. Ser. 2012, 458, 213–229. [Google Scholar] [CrossRef]
- Gilad, O.; Yun, S.; Adkison, M.A.; Way, K.; Willits, N.H.; Bercovier, H.; Hedrick, R.P. Molecular Comparison of Isolates of an Emerging Fish Pathogen, Koi Herpesvirus, and the Effect of Water Temperature on Mortality of Experimentally Infected Koi. J. Gen. Virol. 2003, 84, 2661–2668. [Google Scholar] [CrossRef]
- Foley, A.M.; Schroeder, B.A.; Redlow, A.E.; Fick-Child, K.J.; Teas, W.G. Fibropapillomatosis in Stranded Green Turtles (Chelonia mydas) from the Eastern United States (1980–98): Trends and Associations with Environmental Factors. J. Wildl. Dis. 2005, 41, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Haines, H.; Kleese, W. Effect of Water Temperature on Performance. Infect. Immun. 2013, 15, 2012–2013. [Google Scholar]
- McGarrity, M.E. Stress Protein Expression and Green Turtle Fibropapillomatosis. Master’s Thesis, Florida Atlantic University, Boca Raton, FL, USA, 2005; pp. 1–73. [Google Scholar]
- Herbst, L.H.; Klein, P.A. Green Turtle Fibropapillomatosis: Challenges to Assessing the Role of Environmental Cofactors. Environ. Health Perspect. 1995, 103, 27–30. [Google Scholar] [CrossRef]
- Page-Karjian, A.; Norton, T.M.; Krimer, P.; Groner, M.; Nelson, S.E.; Gottdenker, N.L. Factors Influencing Survivorship of Rehabilitating Green Sea Turtles (Chelonia mydas) with Fibropapillomatosis. J. Zoo Wildl. Med. 2014, 45, 507–519. [Google Scholar] [CrossRef]
- Abraham, J.P.; Baringer, M.; Bindoff, N.L.; Boyer, T.; Cheng, L.J.; Church, J.A.; Conroy, J.L.; Domingues, C.M.; Fasullo, J.T.; Gilson, J.; et al. A Review of Global Ocean Temperature Observations: Implications for Ocean Heat Content Estimates and Climate Change. Rev. Geophys. 2013, 51, 450–483. [Google Scholar] [CrossRef]
- Jorgenson, A.K.; Fiske, S.; Hubacek, K.; Li, J.; McGovern, T.; Rick, T.; Schor, J.B.; Solecki, W.; York, R.; Zycherman, A. Social Science Perspectives on Drivers of and Responses to Global Climate Change. Wiley Interdiscip. Rev. Clim. Chang. 2019, 10, e554. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.J.; Martindale, M.Q. Perspectives on the Expansion of Human Precision Oncology and Genomic Approaches to Sea Turtle Fibropapillomatosis. Commun. Biol. 2019, 2, 54. [Google Scholar] [CrossRef]
- Herren, R.M.; Bagley, D.A.; Bresette, M.J.; Holloway-Adkins, K.G.; Clark, D.; Witherington, B.E. Sea Turtle Abundance and Demographic Measurements in a Marine Protected Areas in the Florida Keys (USA). Herpetol. Conserv. Biol. 2018, 26, 224–239. [Google Scholar]
- Heithaus, M.R.; Alcoverro, T.; Arthur, R.; Burkholder, D.A.; Coates, K.A.; Christianen, M.J.A.; Kelkar, N.; Manuel, S.A.; Wirsing, A.J.; Kenworthy, W.J.; et al. Seagrasses in the Age of Sea Turtle Conservation and Shark Overfishing. Front. Mar. Sci. 2014, 1, 28. [Google Scholar] [CrossRef]
- McLeod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A Blueprint for Blue Carbon: Toward an Improved Understanding of the Role of Vegetated Coastal Habitats in Sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Hofmann, D.J.; Butler, J.H.; Tans, P.P. A New Look at Atmospheric Carbon Dioxide. Atmos. Environ. 2009, 43, 2084–2086. [Google Scholar] [CrossRef]
- McMichael, A.J.; Woodruff, R.E.; Hales, S. Climate Change and Human Health: Present and Future Risks. Lancet 2006, 367, 859–869. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Lutz, P. Marine Turtles as Sentinels of Ecosystem Health: Is Fibropapillomatosis an Indicator? Ecohealth 2004, 1, 275–283. [Google Scholar] [CrossRef]
- Capua, I.; Cattoli, G. One Health (r)Evolution: Learning from the Past to Build a New Future. Viruses 2018, 10, 725. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manes, C.; Carthy, R.R.; Hull, V. A Coupled Human and Natural Systems Framework to Characterize Emerging Infectious Diseases—The Case of Fibropapillomatosis in Marine Turtles. Animals 2023, 13, 1441. https://doi.org/10.3390/ani13091441
Manes C, Carthy RR, Hull V. A Coupled Human and Natural Systems Framework to Characterize Emerging Infectious Diseases—The Case of Fibropapillomatosis in Marine Turtles. Animals. 2023; 13(9):1441. https://doi.org/10.3390/ani13091441
Chicago/Turabian StyleManes, Costanza, Raymond R. Carthy, and Vanessa Hull. 2023. "A Coupled Human and Natural Systems Framework to Characterize Emerging Infectious Diseases—The Case of Fibropapillomatosis in Marine Turtles" Animals 13, no. 9: 1441. https://doi.org/10.3390/ani13091441
APA StyleManes, C., Carthy, R. R., & Hull, V. (2023). A Coupled Human and Natural Systems Framework to Characterize Emerging Infectious Diseases—The Case of Fibropapillomatosis in Marine Turtles. Animals, 13(9), 1441. https://doi.org/10.3390/ani13091441