
A Pilot Study on Behavioural and Physiological Indicators of Emotions in Donkeys

 , , ,
, , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Farm Selection
2.3. Experimental Design
2.3.1. Test Area and Manipulandum
2.3.2. Training and Testing
2.4. Behavioural Observations
2.5. Heart Rate and Heart Rate Variability Analysis
2.6. Statistical Analyses
3. Results
3.1. Behavioural Observations
3.2. Heart Rate and Heart Rate Variability
4. Discussion
4.1. Facial Expressions and Postures
4.2. HRV Analysis
4.3. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dawkins, M.S. Behavioural Deprivation: A Central Problem in Animal Welfare. Appl. Anim. Behav. Sci. 1988, 20, 209–225. [Google Scholar] [CrossRef]
- Fraser, D. Understanding Animal Welfare. Acta Vet. Scand. 2008, 50, S1. [Google Scholar] [CrossRef]
- Kremer, L.; Klein Holkenborg, S.E.J.; Reimert, I.; Bolhuis, J.E.; Webb, L.E. The Nuts and Bolts of Animal Emotion. Neurosci. Biobehav. Rev. 2020, 113, 273–286. [Google Scholar] [CrossRef] [PubMed]
- De Waal, F.B.M. What Is an Animal Emotion? Ann. N. Y. Acad. Sci. 2011, 1224, 191–206. [Google Scholar] [CrossRef]
- Neethirajan, S.; Reimert, I.; Kemp, B. Measuring Farm Animal Emotions—Sensor-Based Approaches. Sensors 2021, 21, 553. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.S.; Mendl, M.T. Animal Emotion: Descriptive and Prescriptive Definitions and Their Implications for a Comparative Perspective. Appl. Anim. Behav. Sci. 2018, 205, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of Positive Emotions in Animals to Improve Their Welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D. Animal Emotions, Behaviour and the Promotion of Positive Welfare States. N. Z. Vet. J. 2012, 60, 1–8. [Google Scholar] [CrossRef]
- Désiré, L.; Boissy, A.; Veissier, I. Emotions in Farm Animals: A New Approach to Animal Welfare in Applied Ethology. Behav. Processes 2002, 60, 165–180. [Google Scholar] [CrossRef]
- Mendl, M.; Burman, O.H.P.; Paul, E.S. An Integrative and Functional Framework for the Study of Animal Emotion and Mood. Proc. R. Soc. B Biol. Sci. 2010, 277, 2895–2904. [Google Scholar] [CrossRef]
- Panksepp, J. The Basic Emotional Circuits of Mammalian Brains: Do Animals Have Affective Lives? Neurosci. Biobehav. Rev. 2011, 35, 1791–1804. [Google Scholar] [CrossRef]
- Boissy, A.; Arnould, C.; Chaillou, E.; Désiré, L.; Duvaux-Ponter, C.; Greiveldinger, L.; Leterrier, C.; Richard, S.; Roussel, S.; Saint-Dizier, H.; et al. Emotions and Cognition: A New Approach to Animal Welfare. Anim. Welf. 2007, 16, 37–43. [Google Scholar] [CrossRef]
- De Vere, A.J.; Kuczaj, S.A. Where Are We in the Study of Animal Emotions? Wiley Interdiscip. Rev. Cogn. Sci. 2016, 7, 354–362. [Google Scholar] [CrossRef]
- Galardi, M.; Contalbrigo, L.; Toson, M.; Bortolotti, L.; Lorenzetto, M.; Riccioli, F.; Moruzzo, R. Donkey Assisted Interventions: A Pilot Survey on Service Providers in North-Eastern Italy. Explore 2022, 18, 10–16. [Google Scholar] [CrossRef] [PubMed]
- McLean, A.K.; Navas Gonzalez, F.J. Can Scientists Influence Donkey Welfare? Historical Perspective and a Contemporary View. J. Equine Vet. Sci. 2018, 65, 25–32. [Google Scholar] [CrossRef]
- Yeates, J.W.; Main, D.C.J. Assessment of Positive Welfare: A Review. Vet. J. 2008, 175, 293–300. [Google Scholar] [CrossRef]
- De Santis, M.; Seganfreddo, S.; Galardi, M.; Mutinelli, F.; Normando, S.; Contalbrigo, L. Donkey Behaviour and Cognition: A Literature Review. Appl. Anim. Behav. Sci. 2021, 244, 105485. [Google Scholar] [CrossRef]
- Tarazona, A.M.; Ceballos, M.C.; Broom, D.M. Human Relationships with Domestic and Other Animals: One Health, One Welfare, One Biology. Animals 2019, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Lansade, L.; Nowak, R.; Lainé, A.L.; Leterrier, C.; Bonneau, C.; Parias, C.; Bertin, A. Facial Expression and Oxytocin as Possible Markers of Positive Emotions in Horses. Sci. Rep. 2018, 8, 14680. [Google Scholar] [CrossRef]
- Descovich, K. Facial Expression: An under-Utilised Tool for the Assessment of Welfare in Mammals. ALTEX 2017, 34, 409–429. [Google Scholar] [CrossRef] [PubMed]
- Mogil, J.S.; Pang, D.S.J.; Silva Dutra, G.G.; Chambers, C.T. The Development and Use of Facial Grimace Scales for Pain Measurement in Animals. Neurosci. Biobehav. Rev. 2020, 116, 480–493. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Hernández, E.; Martínez-Burnes, J.; Whittaker, A.L. The Utility of Grimace Scales for Practical Pain Assessment in Laboratory Animals. Animals 2020, 10, 1838. [Google Scholar] [CrossRef]
- Häger, C.; Biernot, S.; Buettner, M.; Glage, S.; Keubler, L.M.; Held, N.; Bleich, E.M.; Otto, K.; Müller, C.W.; Decker, S.; et al. The Sheep Grimace Scale as an Indicator of Post-Operative Distress and Pain in Laboratory Sheep. PLoS ONE 2017, 12, e0175839. [Google Scholar] [CrossRef]
- Evangelista, M.C.; Watanabe, R.; Leung, V.S.Y.; Monteiro, B.P.; O’Toole, E.; Pang, D.S.J.; Steagall, P.V. Facial Expressions of Pain in Cats: The Development and Validation of a Feline Grimace Scale. Sci. Rep. 2019, 9, 19128. [Google Scholar] [CrossRef]
- Dalla Costa, E.; Minero, M.; Lebelt, D.; Stucke, D.; Canali, E.; Leach, M.C. Development of the Horse Grimace Scale (HGS) as a Pain Assessment Tool in Horses Undergoing Routine Castration. PLoS ONE 2014, 9, e92281. [Google Scholar] [CrossRef]
- Orth, E.K.; Navas González, F.J.; Iglesias Pastrana, C.; Berger, J.M.; Jeune, S.S.L.; Davis, E.W.; McLean, A.K. Development of a Donkey Grimace Scale to Recognize Pain in Donkeys (Equus Asinus) Post Castration. Animals 2020, 10, 1411. [Google Scholar] [CrossRef] [PubMed]
- Van Dierendonck, M.C.; Burden, F.A.; Rickards, K.; van Loon, J.P.A.M. Monitoring Acute Pain in Donkeys with the Equine Utrecht University Scale for Donkeys Composite Pain Assessment (EQUUS-DONKEY-COMPASS) and the Equine Utrecht University Scale for Donkey Facial Assessment of Pain (EQUUS-DONKEY-FAP). Animals 2020, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.G.C.; de Paula, V.V.; Mouta, A.N.; de Oliveira Lima, I.; de Macêdo, L.B.; Nunes, T.L.; Trindade, P.H.E.; Luna, S.P.L. Validation of the Donkey Pain Scale (DOPS) for Assessing Postoperative Pain in Donkeys. Front. Vet. Sci. 2021, 8, 671330. [Google Scholar] [CrossRef] [PubMed]
- Grint, N.J.; Beths, T.; Yvorchuk-St Jean, K.; Whay, H.R.; Murrell, J.C. Analysis of Behaviors Observed During Mechanical Nociceptive Threshold Testing in Donkeys and Horses. J. Equine Vet. Sci. 2017, 50, 102–109. [Google Scholar] [CrossRef]
- Regan, F.H.; Hockenhull, J.; Pritchard, J.C.; Waterman-Pearson, A.E.; Whay, H.R. Identifying behavioural differences in working donkeys in response to analgesic administration. Equine Vet. J. 2016, 48, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Von Borell, E.; Langbein, J.; Després, G.; Hansen, S.; Leterrier, C.; Marchant-Forde, J.; Marchant-Forde, R.; Minero, M.; Mohr, E.; Prunier, A.; et al. Heart Rate Variability as a Measure of Autonomic Regulation of Cardiac Activity for Assessing Stress and Welfare in Farm Animals—A Review. Physiol. Behav. 2007, 92, 293–316. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, F.; Ginsberg, J.P. An Overview of Heart Rate Variability Metrics and Norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef]
- De Santis, M.; Seganfreddo, S.; Greco, A.; Normando, S.; Benedetti, D.; Mutinelli, F.; Contalbrigo, L. Donkey Heart Rate and Heart Rate Variability: A Scoping Review. Animals 2023, 13, 408. [Google Scholar] [CrossRef]
- McLean, A.K.; Heleski, C.R.; Yokoyama, M.T.; Wang, W.; Doumbia, A.; Dembele, B. Improving Working Donkey (Equus Asinus) Welfare and Management in Mali, West Africa. J. Vet. Behav. Clin. Appl. Res. 2012, 7, 123–134. [Google Scholar] [CrossRef]
- Panzera, M.; Alberghina, D.; Statelli, A. Ethological and Physiological Parameters Assessment in Donkeys Used in Animal Assisted Interventions. Animals 2020, 10, 1867. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Kim, J.G.; Lee, H.; Kim, B.S. Behavioral and Cardiac Responses in Mature Horses Exposed to a Novel Object. J. Anim. Sci. Technol. 2021, 63, 651. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.; Randle, H.; Pearson, G.; Preshaw, L.; Waran, N. Assessing equine emotional state. Appl. Anim. Behav. Sci. 2018, 205, 183–193. [Google Scholar] [CrossRef]
- Guidi, A.; Lanata, A.; Baragli, P.; Valenza, G.; Scilingo, E. A Wearable System for the Evaluation of the Human-Horse Interaction: A Preliminary Study. Electronics 2016, 5, 63. [Google Scholar] [CrossRef]
- Stucke, D.; Große Ruse, M.; Lebelt, D. Measuring Heart Rate Variability in Horses to Investigate the Autonomic Nervous System Activity—Pros and Cons of Different Methods. Appl. Anim. Behav. Sci. 2015, 166, 1–10. [Google Scholar] [CrossRef]
- Seganfreddo, S.; Fornasiero, D.; De Santis, M.; Contalbrigo, L.; Mutinelli, F.; Normando, S. Investigation of Donkeys Learning Capabilities through an Operant Conditioning. Appl. Anim. Behav. Sci. 2022, 255, 105743. [Google Scholar] [CrossRef]
- Lambert, H.; Carder, G. Positive and negative emotions in dairy cows: Can ear postures be used as a measure? Behav. Process. 2019, 158, 172–180. [Google Scholar] [CrossRef]
- Lv, J.; Li, J.; Wang, C.; Zhao, P.; Bi, Y.; Zhang, X.; Yi, R.; Li, X.; Bao, J. Positive or negative emotion induced by feeding success or failure can affect behaviors, heart rate and immunity of suckling calves. Physiol Behav. 2018, 196, 185–189. [Google Scholar] [CrossRef]
- Reefmann, N.; Bütikofer Kaszàs, F.; Wechsler, B.; Gygax, L. Ear and Tail Postures as Indicators of Emotional Valence in Sheep. Appl. Anim. Behav. Sci. 2009, 118, 199–207. [Google Scholar] [CrossRef]
- Friard, O.; Gamba, M. BORIS: A Free, Versatile Open-source Event-logging Software for Video/Audio Coding and Live Observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Tarvainen, M.P.; Niskanen, J.P.; Lipponen, J.A.; Ranta-aho, P.O.; Karjalainen, P.A. Kubios HRV–Heart Rate Variability Analysis Software. Comput. Methods Programs Biomed. 2014, 113, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, M.; Hashimoto, S.; Ishii, K.; Yagi, Y.; Hada, T.; Hiraga, A.; Kai, M.; Kubo, K.; Oki, H.; Tsubone, H.; et al. Assessment of Autonomic Nervous Function by Power Spectral Analysis of Heart Rate Variability in the Horse. J. Auton. Nerv. Syst. 1996, 60, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Mott, R.; Dowell, F.; Evans, N. Use of the Polar V800 and Actiheart 5 Heart Rate Monitors for the Assessment of Heart Rate Variability (HRV) in Horses. Appl. Anim. Behav. Sci. 2021, 241, 105401. [Google Scholar] [CrossRef]
- Scopa, C.; Palagi, E.; Sighieri, C.; Baragli, P. Physiological Outcomes of Calming Behaviors Support the Resilience Hypothesis in Horses. Sci. Rep. 2018, 8, 17501. [Google Scholar] [CrossRef] [PubMed]
- Visser, E.K.; van Reenen, C.G.; van der Werf, J.T.N.; Schilder, M.B.H.; Knaap, J.H.; Barneveld, A.; Blokhuis, H.J. Heart Rate and Heart Rate Variability during a Novel Object Test and a Handling Test in Young Horses. Physiol. Behav. 2002, 76, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, S.Z.H.; Zhang, H.; Aziz, W.; Monfredi, O.; Abbas, S.A.; Shah, S.A.; Kazmi, S.S.H.; Butt, W.H. Inverse Correlation between Heart Rate Variability and Heart Rate Demonstrated by Linear and Nonlinear Analysis. PLoS ONE 2016, 11, e0157557. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal Component Analysis: A Review and Recent Developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. 2022. Available online: https://www.r-project.org/ (accessed on 21 February 2023).
- RStudio Team. RStudio: Integrated Development Environment for R. 2022. Available online: https://posit.co/ (accessed on 21 February 2023).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. vegan: Community Ecology Package. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 21 February 2023).
- Derrick, B. Partiallyoverlapping: Partially Overlapping Samples Tests. 2018. Available online: https://CRAN.R-project.org/package=Partiallyoverlapping (accessed on 21 February 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Vu, V.Q. ggbiplot: A ggplot2 Based Biplot. 2011. Available online: http://github.com/vqv/ggbiplot (accessed on 21 February 2023).
- Slowikowski, K. ggrepel: Automatically Position Non-Overlapping Text Labels with ’ggplot2’. 2021. Available online: https://CRAN.R-project.org/package=ggrepel (accessed on 21 February 2023).
- Fureix, C.; Jego, P.; Sankey, C.; Hausberger, M. How Horses (Equus Caballus) See the World: Humans as Significant “Objects”. Anim. Cogn. 2009, 12, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Sankey, C.; Richard-Yris, M.-A.; Henry, S.; Fureix, C.; Nassur, F.; Hausberger, M. Reinforcement as a Mediator of the Perception of Humans by Horses (Equus Caballus). Anim. Cogn. 2010, 13, 753–764. [Google Scholar] [CrossRef]
- Wong, P.T. Frustration, Exploration, and Learning. Can. Psychol. Rev. Psychol. Can. 1979, 20, 133–144. [Google Scholar] [CrossRef]
- Tamioso, P.R.; Maiolino Molento, C.F.; Boivin, X.; Chandèze, H.; Andanson, S.; Delval, É.; Hazard, D.; da Silva, G.P.; Taconeli, C.A.; Boissy, A. Inducing Positive Emotions: Behavioural and Cardiac Responses to Human and Brushing in Ewes Selected for High vs Low Social Reactivity. Appl. Anim. Behav. Sci. 2018, 208, 56–65. [Google Scholar] [CrossRef]
- Stomp, M.; Leroux, M.; Cellier, M.; Henry, S.; Lemasson, A.; Hausberger, M. An Unexpected Acoustic Indicator of Positive Emotions in Horses. PLoS ONE 2018, 13, e0197898. [Google Scholar] [CrossRef]
- Boissy, A.; Aubert, A.; Désiré, L.; Greiveldinger, L.; Delval, E.; Veissier, I. Cognitive Sciences to Relate Ear Postures to Emotions in Sheep. Anim. Welf. 2011, 20, 47–56. [Google Scholar] [CrossRef]
- Pannewitz, L.; Loftus, L. Frustration in Horses: Investigating Expert Opinion on Behavioural Indicators and Causes Using a Delphi Consultation. Appl. Anim. Behav. Sci. 2023, 258, 105818. [Google Scholar] [CrossRef]
- Lesimple, C. Indicators of Horse Welfare: State-of-the-Art. Animals 2020, 10, 294. [Google Scholar] [CrossRef]
- Rochais, C.; Henry, S.; Hausberger, M. “Hay-Bags” and “Slow Feeders”: Testing Their Impact on Horse Behaviour and Welfare. Appl. Anim. Behav. Sci. 2018, 198, 52–59. [Google Scholar] [CrossRef]
- Cooper, J.J.; Mason, G.J. The Identification of Abnormal Behaviour and Behavioural Problems in Stabled Horses and Their Relationship to Horse Welfare: A Comparative Review. Equine Vet. J. 2010, 30, 5–9. [Google Scholar] [CrossRef]
- McFarland, D.J. On the Causal and Functional Significance of Displacement Activities. Z. Tierpsychol. 2010, 23, 217–235. [Google Scholar] [CrossRef]
- Tinbergen, N. “Derived” Activities; Their Causation, Biological Significance, Origin, and Emancipation During Evolution. Q. Rev. Biol. 1952, 27, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Zeigler, H.P. Displacement Activity and Motivational Theory: A Case Study in the History of Ethology. Psychol. Bull. 1964, 61, 362–376. [Google Scholar] [CrossRef]
- Zucca, P.; Cerri, F.; Carluccio, A.; Baciadonna, L. Space Availability Influence Laterality in Donkeys (Equus Asinus). Behav. Processes 2011, 88, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Leliveld, L.M.C.; Langbein, J.; Puppe, B. The Emergence of Emotional Lateralization: Evidence in Non-Human Vertebrates and Implications for Farm Animals. Appl. Anim. Behav. Sci. 2013, 145, 1–14. [Google Scholar] [CrossRef]
- Delfour, F.; Vaicekauskaite, R.; García-Párraga, D.; Pilenga, C.; Serres, A.; Brasseur, I.; Pascaud, A.; Perlado-Campos, E.; Sánchez-Contreras, G.J.; Baumgartner, K.; et al. Behavioural Diversity Study in Bottlenose Dolphin (Tursiops Truncatus) Groups and Its Implications for Welfare Assessments. Animals 2021, 11, 1715. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.; Bryant, J.; Staley, M.; Whitham, J.C.; Miller, L.J. Behavioural Diversity as a Potential Welfare Indicator for Professionally Managed Chimpanzees (Pan Troglodytes): Exploring Variations in Calculating Diversity Using Species-Specific Behaviours. Anim. Welf. 2021, 30, 381–392. [Google Scholar] [CrossRef]
- Cronin, K.A.; Ross, S.R. Technical contribution: A cautionary note on the use of behavioural diversity (H-Index) in animal welfare science. Anim. Welf. 2019, 28, 157–164. [Google Scholar] [CrossRef]
- Baragli, P.; Vitale, V.; Sighieri, C.; Lanata, A.; Palagi, E.; Reddon, A.R. Consistency and Flexibility in Solving Spatial Tasks: Different Horses Show Different Cognitive Styles. Sci. Rep. 2017, 7, 16557. [Google Scholar] [CrossRef] [PubMed]
- Reefmann, N.; Wechsler, B.; Gygax, L. Behavioural and physiological assessment of positive and negative emotion in sheep. Anim. Behav. 2009, 78, 651–659. [Google Scholar] [CrossRef]
- Melo, H.M.; Hoeller, A.A.; Walz, R.; Takase, E. Resting Cardiac Vagal Tone Is Associated with Long-Term Frustration Level of Mental Workload: Ultra-Short Term Recording Reliability. Appl. Psychophysiol. Biofeedback 2020, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gygax, L.; Reefmann, N.; Wolf, M.; Langbein, J. Prefrontal Cortex Activity, Sympatho-Vagal Reaction and Behaviour Distinguish between Situations of Feed Reward and Frustration in Dwarf Goats. Behav. Brain Res. 2013, 239, 104–114. [Google Scholar] [CrossRef]
- Task Force of the European Society of Cardiology the North American Society of Pacing Electrophysiology. Heart Rate Variability: Standards of Measurement, Physiological Interpretation and Clinical Use. Circulation 1996, 93, 1043–1065. [CrossRef]
- Sheridan, D.C.; Domingo, K.N.; Dehart, R.; Baker, S.D. Heart Rate Variability Duration: Expanding the Ability of Wearable Technology to Improve Outpatient Monitoring? Front. Psychiatry 2021, 12, 943. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| Clinically healthy subjects | Individuals with physical or behavioural pathologies |
| Subject with typical behavioural development | Lactating individuals, females in the last third of pregnancy or in oestrous |
| Subjects used to contact with humans | Subjects not socialised with humans |
| Females or geldings | Stallions |
| Young and adult individuals (age > 12 months) | Subjects with a history of abuse |
| Day | Activity |
|---|---|
| 0 | Acclimatization |
| 1–7 | Training sessions |
| 8 | Positive test |
| 9 | Negative test |
| Behaviour | Variants | Acronym | Description | Event Type |
|---|---|---|---|---|
| Push | The complete pressure of the buzzer. This allows the animal to obtain the reinforcement, with the button starting to blink and making a sound indicating the proper pressure. | Point event | ||
| Attempt | The partial pressure of the button. The animal demonstrates intentionality in pressing the button, however resulting to be too light (with no trigger of the buzzer sound and light) or in the wrong place (e.g., on the coloured border surrounding the button). | Point event | ||
| Bite | The animal bites a part of the manipulandum. | Point event | ||
| Kick | The animal kicks against the manipulandum or towards other donkeys on the sides of the test area. | Point event | ||
| Walk | Walk | Movement from one point to another, with the execution of at least one complete step (movement of all 4 legs). | State event | |
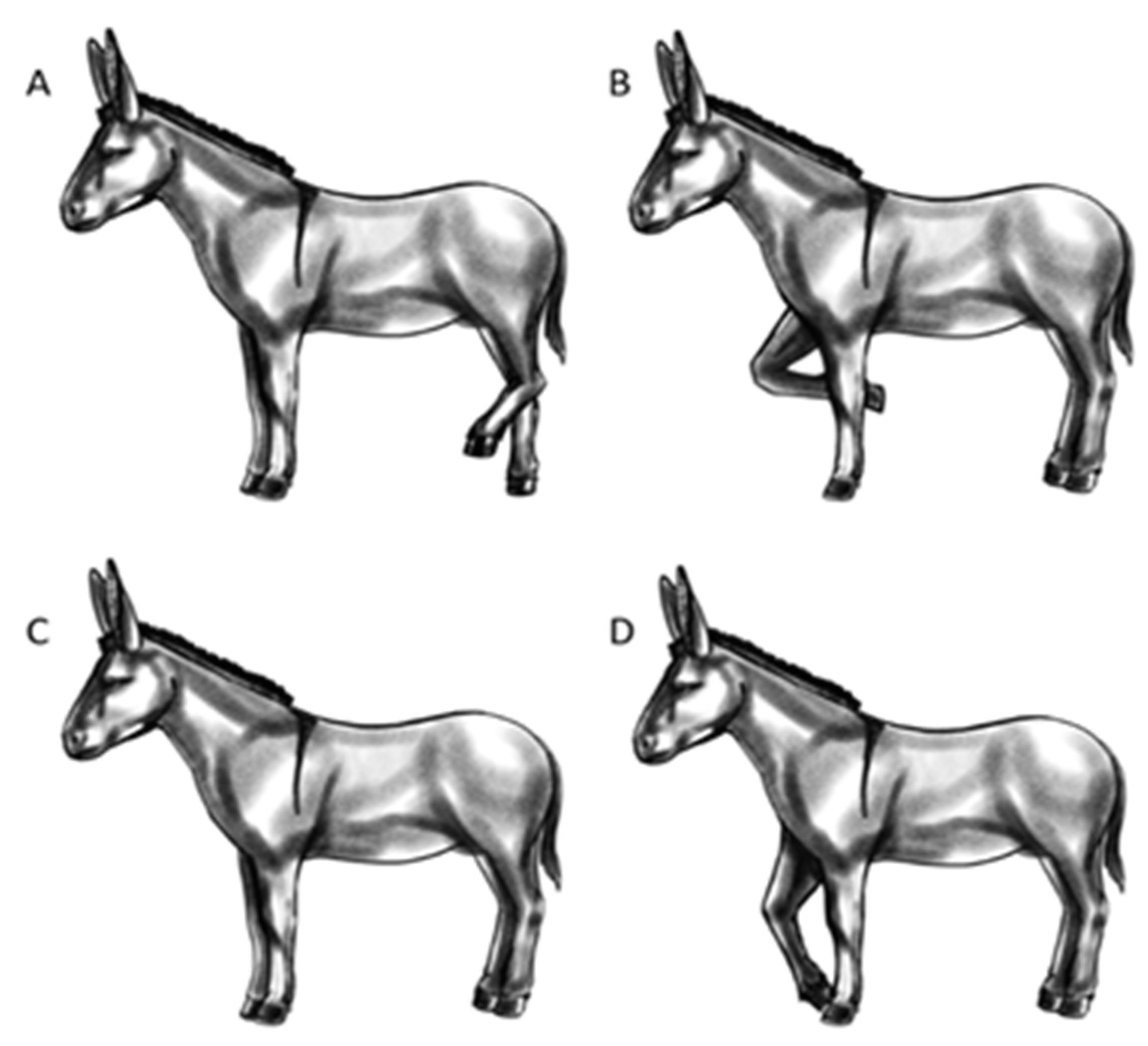
| Standing | 3-legs | 3-legs | The animal stands still (or performs less than 1 complete step). 4-legs: standing with weight-bearing on all 4 limbs with no preferred loading; 3-legs: the weight-bearing is on 3 limbs with a hind limb resting; pointing: the animal places a foot forwards outside of the main body frame (minimum 1 hoof length) with reduced weight-bearing; bent-knee: the animal has one of the front legs with the knee bent and that not bears weight. The standing position variants are graphically presented in Figure 3. | State event |
| 4-legs | 4-legs | |||
| bent knee | Bent knee | |||
| pointing | Pointing | |||
| Walk away | Walk away | The animal loses interest in the manipulandum and walks away from it. | State event | |
| Pawing | Pawing with the forelimbs/investigating with the paw. | Point event | ||
| Stomp | One foreleg is raised and lowered sharply and firmly against the ground. Stomping differs from pawing in that it is a vertical rather than horizontal movement of the leg. | Point event | ||
| Ground exploration | ground | Ground exp | The donkey sniffs the ground (when the donkey moves to sniff somewhere else, the walk is still counted). Ground: the donkey does not eat while sniffing on the ground; ground eating: the animal feeds off the ground while exploring. | State event |
| ground eating | Ground eat | |||
| Area exploration | The animal sniffs/explore other parts of the test area (i.e., fence, straps, iron grid). | State event | ||
| Manipulandum exploration | Manip exp | The animal sniffs/explore parts of the manipulandum. | State event | |
| Stones exploration | The animal sniffs/explore the stones (i.e., the negative reinforcement). | State event | ||
| Scanning environment | Scan env | The donkey looks towards its surroundings. Only the position/direction of the head with respect to the manipulandum is considered. | State event | |
| Scanning manipulandum | Scan Manip | The donkey looks towards the manipulandum (covered or uncovered). Only the position/direction of the head with respect to the manipulandum is considered. | State event | |
| Snort | Forceful exhalation through the nostrils and characterized by an audible flutter pulsation. | Point event | ||
| Vocalizations | Other type of sounds produced by the donkey and different from a snort. | Point event | ||
| Tray removal | The donkey removes/detaches the tray from the manipulandum. | Point event | ||
| Body position | frontal | Body F | The donkey body position with respect to the manipulandum. | State event |
| left | Body L | |||
| right | Body R | |||
| Head position | frontal | Head F | The donkey head position with respect to the manipulandum. | State event |
| left | Head L | |||
| right | Head R | |||
| Left ear position | back-up | L-ear BU | Position of the left ear as described in Figure 2. | State event |
| back-down | L-ear BD | |||
| side-up | L-ear SU | |||
| side-down | L-ear SD | |||
| front-up | L-ear FU | |||
| Right ear position | back-up | R-ear BU | Position of the right ear as described in Figure 2. | State event |
| back-down | R-ear BD | |||
| side-up | R-ear SU | |||
| side-down | R-ear SD | |||
| front-up | R-ear FU | |||
| Feed chewing | Feed chew | The donkey eats and chew feed. | State event | |
| Vacuum chewing | Vacuum chew | Chewing with nothing in the mouth. | State event | |
| Mouth movements | Mouth mov | Unusual movements of the mouth that are not related to feed chewing. | State event | |
| Yawn | The donkey yawns. | Point event | ||
| Flehmen | The upper lip curls back to expose gums with incisors meeting, head tips back and rapidly points muzzle upwards. | Point event |
| Index | Description | Numerical Range [Min; Max] |
|---|---|---|
| Behavioural frequencies | Number of behavioural variants manifested per minute for each type of behaviour. | [0; +∞] |
| Activity budgets | The percentage of time spent by donkeys in manifesting each behavioural variant. | [0; 100] |
| Richness | The count of the different behavioural types exhibited by the animals in the corresponding ethogram. | [0; +∞] |
| Shannon’s diversity index | Nonlinear index that considers both the behavioural richness and evenness. Higher index values reflect a larger numbers of behaviour types and more equal abundances. | [0; +∞] |
| Gini-Simpson index | A diversity index that measures the probability that two behaviours randomly selected from a sample will be different. | [0; 1] |
| (+) Test | (−) Test | |||||
|---|---|---|---|---|---|---|
| Pre a | Test a | Post b | Pre a | Test b | Post a | |
| RR (ms) | 1116.60 [998.0; 1187.80] | 1026.00 [944.40; 1119.10] | 1086.00 [896.00; 1182.00] | 1218.00 [1042.10; 1288.70] | 1122.80 [979.10; 1147.00] | 1079.00 [1007.20; 1150.70] |
| Std RR (ms) | 102.68 [88.72; 127.27] | 139.90 [133.74; 169.68] | 122.39 [84.77; 169.87] | 104.39 [90.62; 129.89] | 128.02 [109.98; 192.84] | 118.29 [51.72; 141.70] |
| HR (beats/min) | 53.74 [50.59; 60.25] | 58.49 [53.65; 63.69] | 55.26 [50.76; 67.20] | 49.26 [46.59; 57.58] | 53.44 [52.31; 61.68] | 55.62 [52.14; 59.57] |
| rMSSD (ms) | 94.00 [90.42; 112.88] | 127.77 [88.45; 150.37] | 120.38 [114.98; 155.50] | 99.90 [92.20; 133.70] | 125.43 [103.68; 172.36] | 62.85 [49.18; 86.67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seganfreddo, S.; Fornasiero, D.; De Santis, M.; Mutinelli, F.; Normando, S.; Contalbrigo, L. A Pilot Study on Behavioural and Physiological Indicators of Emotions in Donkeys. Animals 2023, 13, 1466. https://doi.org/10.3390/ani13091466
Seganfreddo S, Fornasiero D, De Santis M, Mutinelli F, Normando S, Contalbrigo L. A Pilot Study on Behavioural and Physiological Indicators of Emotions in Donkeys. Animals. 2023; 13(9):1466. https://doi.org/10.3390/ani13091466
Chicago/Turabian StyleSeganfreddo, Samanta, Diletta Fornasiero, Marta De Santis, Franco Mutinelli, Simona Normando, and Laura Contalbrigo. 2023. "A Pilot Study on Behavioural and Physiological Indicators of Emotions in Donkeys" Animals 13, no. 9: 1466. https://doi.org/10.3390/ani13091466
APA StyleSeganfreddo, S., Fornasiero, D., De Santis, M., Mutinelli, F., Normando, S., & Contalbrigo, L. (2023). A Pilot Study on Behavioural and Physiological Indicators of Emotions in Donkeys. Animals, 13(9), 1466. https://doi.org/10.3390/ani13091466