
Molecular Characterization and Functional Analysis of Hypoxia-Responsive Factor Prolyl Hydroxylase Domain 2 in Mandarin Fish (Siniperca chuatsi)
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Cells
2.2. Molecular Cloning of Mandarin Fish PHD2 (scPHD2) cDNAs
2.3. Sequence Analysis
2.4. Three-Dimensional Structure Prediction
2.5. Dual-Luciferase Reporter Gene Assays
2.6. Real-Time Quantitative PCR (RT-qPCR)
2.7. Indirect Immunofluorescence Assay (IFA)
2.8. Co-Immunoprecipitation (Co-IP) and Western Blot Analysis
2.9. Protein-Degradation Experiment
2.10. Statistical Analysis
3. Results
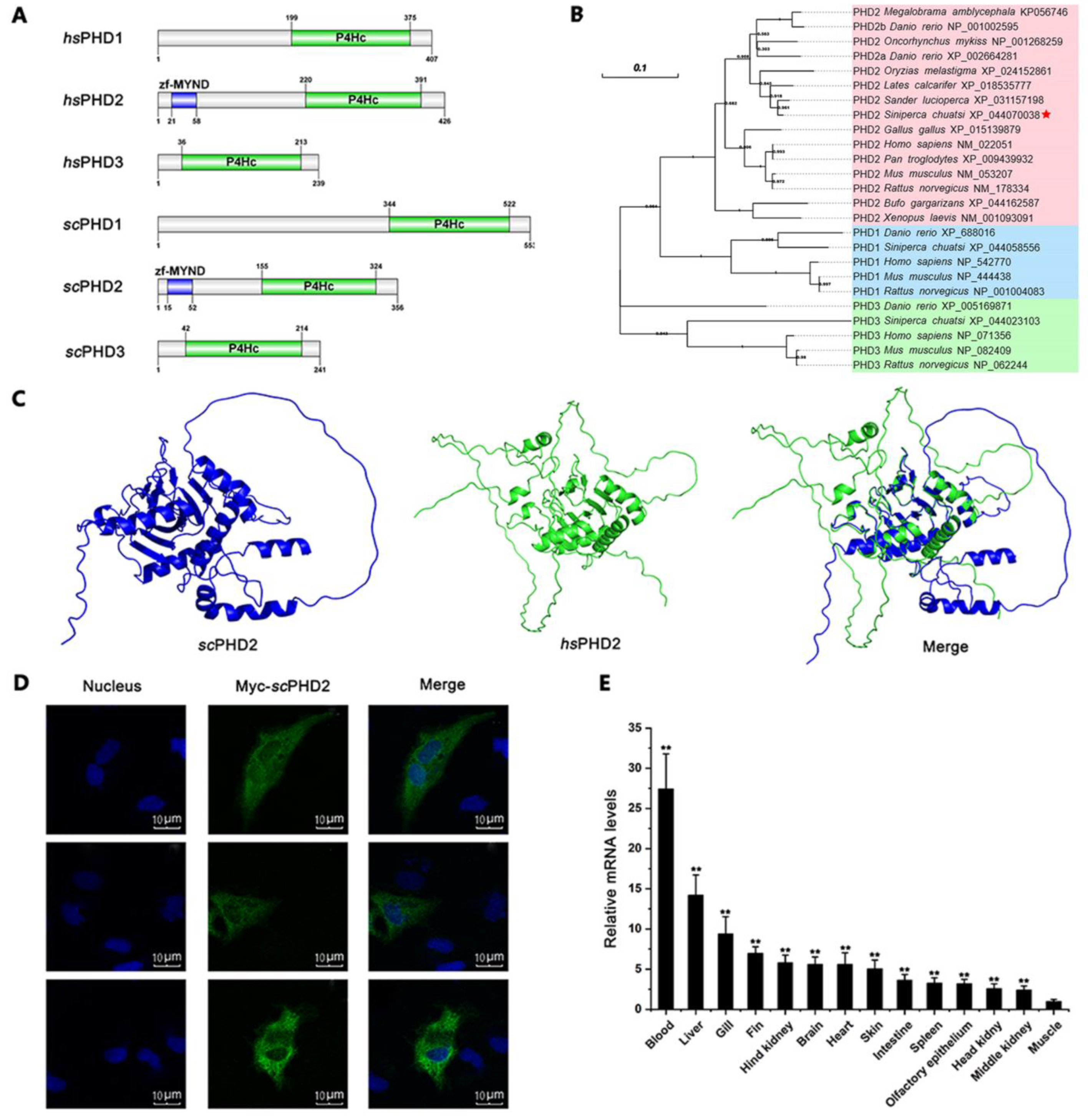
3.1. Molecular Characteristics of scPHD2
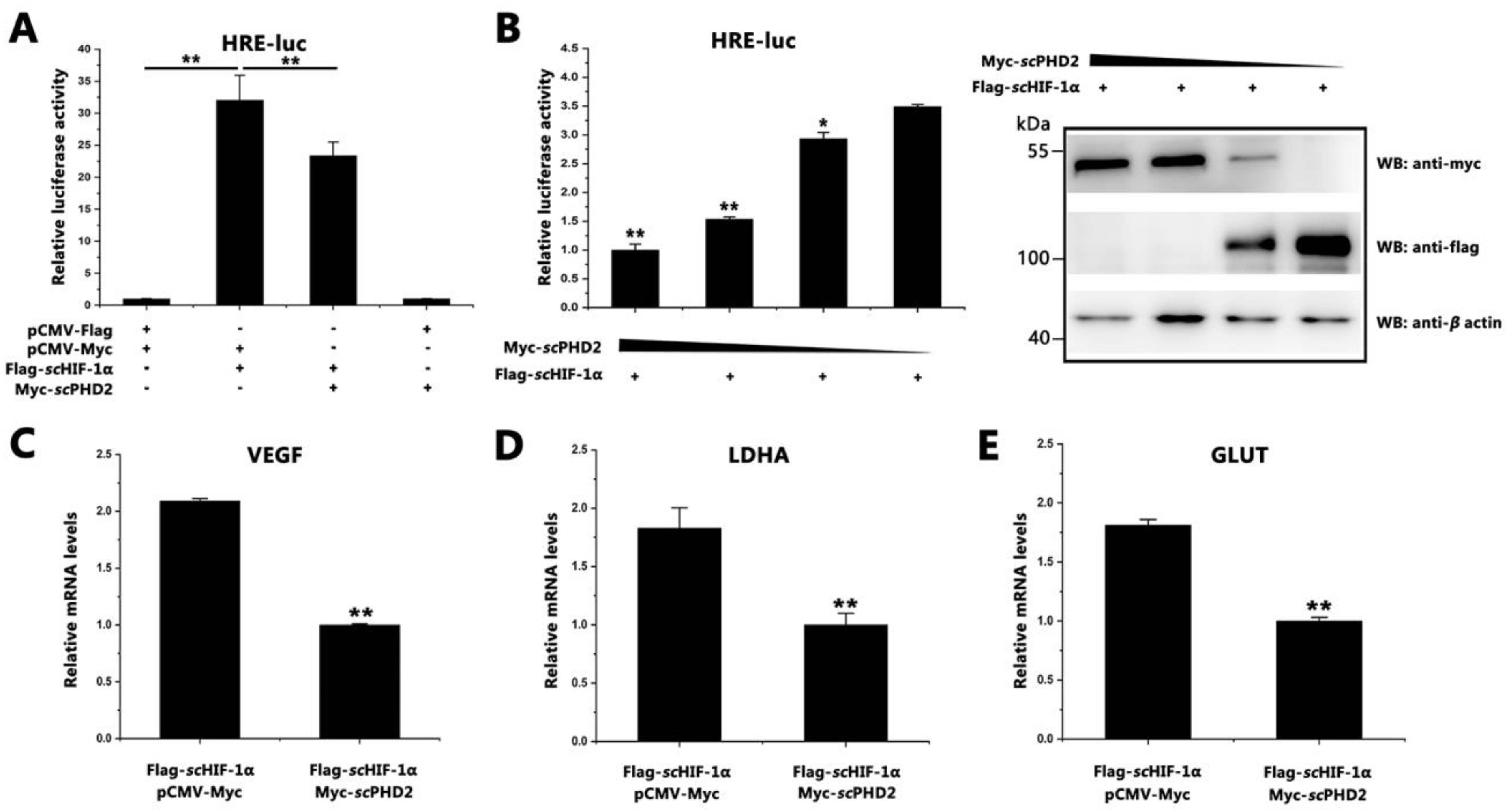
3.2. scPHD2 Inhibited the HIF-1 Signaling Pathway
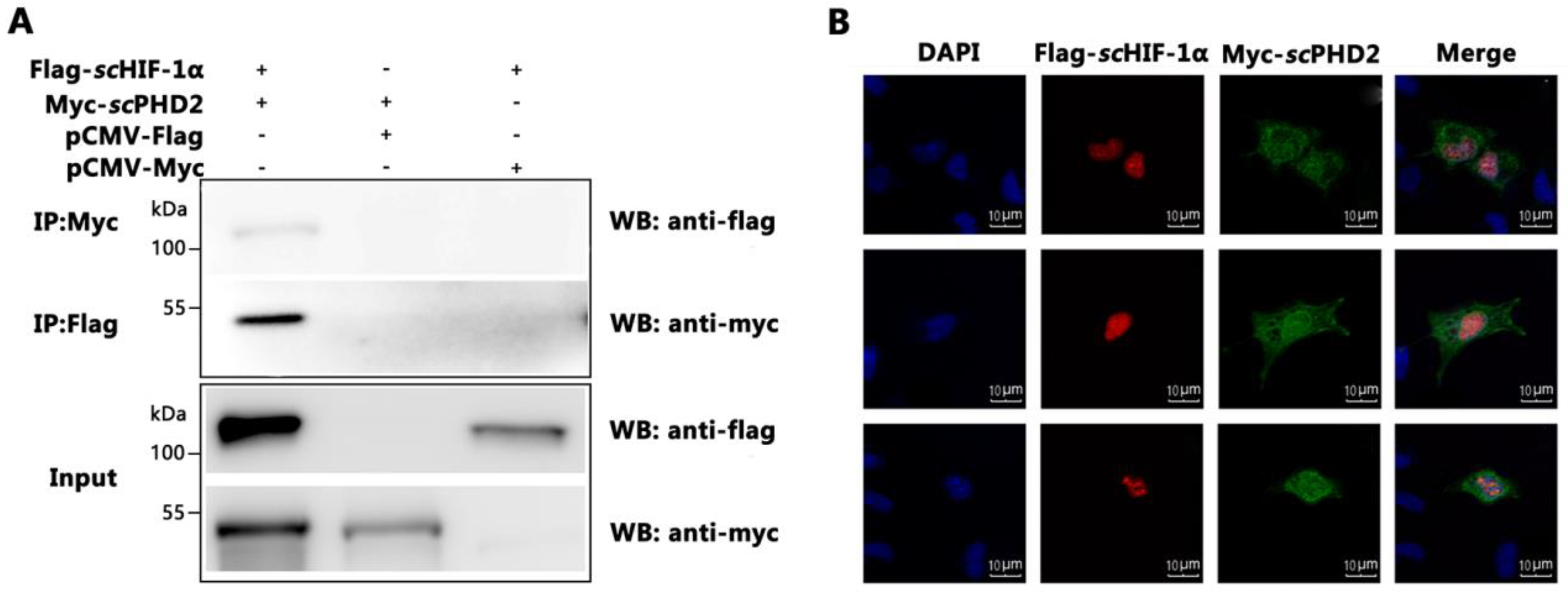
3.3. scPHD2 Interacted with scHIF-1α
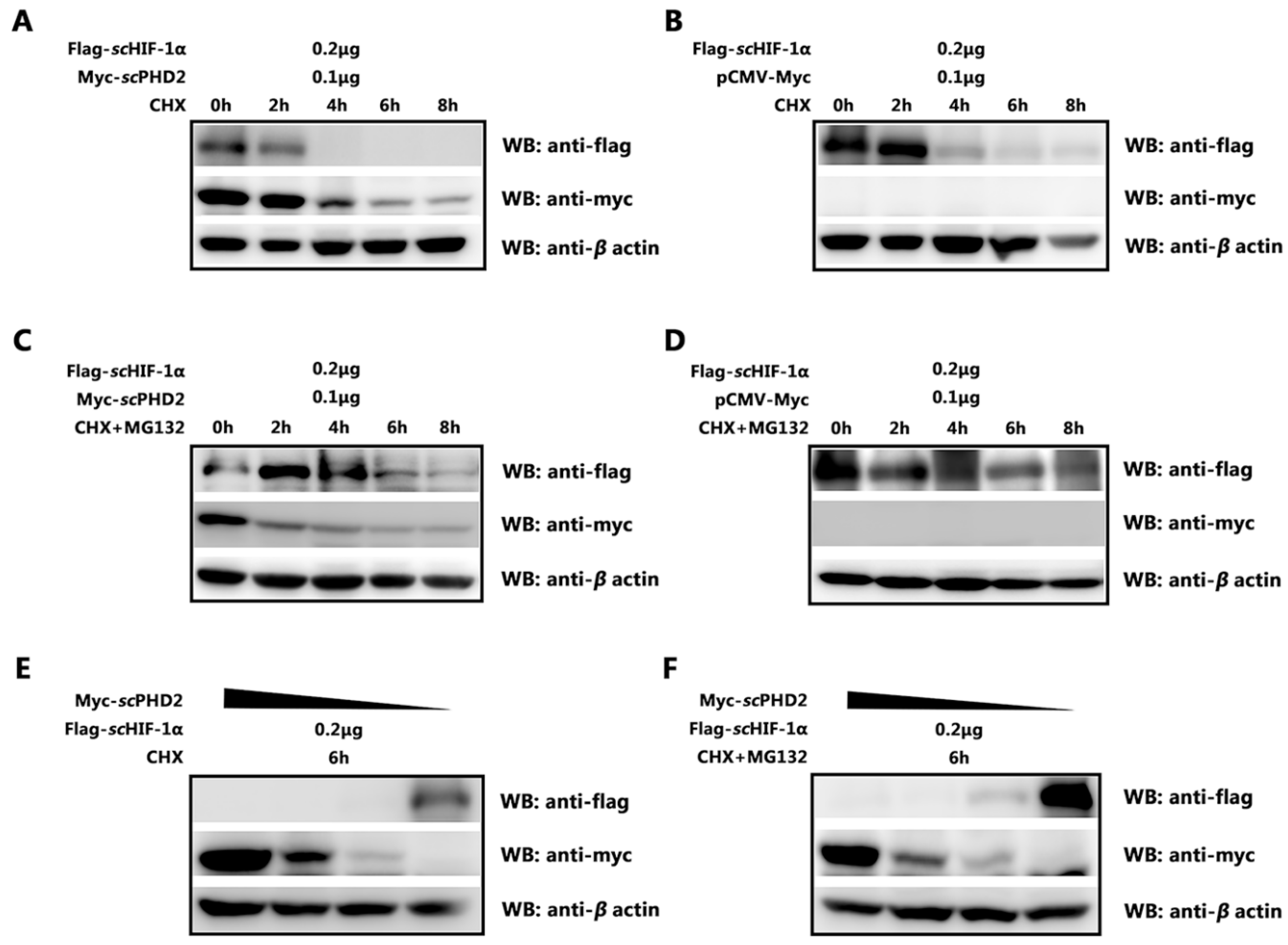
3.4. scPHD2 Promoted the Degradation of scHIF-1α and Its Degradation Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, C.T.; Pouyssegur, J. Oxygen, Hypoxia, and Stress. Ann. N. Y. Acad. Sci. 2007, 1113, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W.; Lutz, P.L. Mechanism, origin, and evolution of anoxia tolerance in animals☆. Comp. Biochem. Physiol. Part B 2001, 130, 435–459. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, C.A.; Hinshaw, J. Physiological and haematological responses in rainbow trout subjected to supplemental dissolved oxygen in fish culture. Aquaculture 1994, 126, 183–193. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Monier, M.N.; Hoseinifar, S.H.; Faggio, C. Fish response to hypoxia stress: Growth, physiological, and immunological biomarkers. Fish Physiol. Biochem. 2019, 45, 997–1013. [Google Scholar] [CrossRef]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen Sensing by Metazoans: The Central Role of the HIF Hydroxylase Pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef]
- Pouysségur, J.; Dayan, F.; Mazure, N.M. Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature 2006, 441, 437–443. [Google Scholar] [CrossRef]
- Piontkivska, H.; Chung, J.S.; Ivanina, A.; Sokolov, E.; Techa, S.; Sokolova, I. Molecular characterization and mRNA expression of two key enzymes of hypoxia-sensing pathways in eastern oysters Crassostrea virginica (Gmelin): Hypoxia-inducible factor α (HIF-α) and HIF-prolyl hydroxylase (PHD). Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 103–114. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1, O2, and the 3 PHDs: How animal cells signal hypoxia to the nucleus. Cell 2001, 107, 1–20. [Google Scholar] [CrossRef]
- Bruick, R.K.; McKnight, S.L. A Conserved Family of Prolyl-4-Hydroxylases That Modify HIF. Science 2001, 294, 1337–1340. [Google Scholar] [CrossRef]
- McNeill, L.A.; Hewitson, K.S.; Gleadle, J.M.; Horsfall, L.E.; Oldham, N.J.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J.; Schofield, C.J. The use of dioxygen by HIF prolyl hydroxylase (PHD1). Bioorganic Med. Chem. Lett. 2002, 12, 1547–1550. [Google Scholar] [CrossRef]
- Giaccia, A.J.; Simon, M.C.; Johnson, R. The biology of hypoxia: The role of oxygen sensing in development, normal function, and disease. Genes Dev. 2004, 18, 2183–2194. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, Q.; Mooney, S.M.; Lee, F.S. Sequence determinants in hypoxia-inducible factor-1α for hydroxylation by the prolyl hydroxylases PHD1, PHD2, and PHD3. J. Biol. Chem. 2002, 277, 39792–39800. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.S. Characterization and comparative analysis of the EGLN gene family. Gene 2001, 275, 125–132. [Google Scholar] [CrossRef]
- Rytkönen, K.T.; Williams, T.A.; Renshaw, G.M.; Primmer, C.R.; Nikinmaa, M. Molecular Evolution of the Metazoan PHD–HIF Oxygen-Sensing System. Mol. Biol. Evol. 2011, 28, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, P.H.; Wiesener, M.S.; Chang, G.-W.; Clifford, S.C.; Vaux, E.C.; Cockman, M.E.; Wykoff, C.C.; Pugh, C.W.; Maher, E.R.; Ratcliffe, P.J. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 1999, 399, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Ivan, M.; Kondo, K.; Yang, H.; Kim, W.; Valiando, J.; Ohh, M.; Salic, A.; Asara, J.M.; Lane, W.S.; Kaelin, W.G., Jr. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Science 2001, 292, 464–468. [Google Scholar] [CrossRef]
- Jaakkola, P.; Mole, D.R.; Tian, Y.M.; Wilson, M.I.; Gielbert, J.; Gaskell, S.J.; von Kriegsheim, A.; Hebestreit, H.F.; Mukherji, M.; Schofield, C.J.; et al. Targeting of HIF-alpha to the von Hippel–Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001, 292, 468–472. [Google Scholar] [CrossRef]
- Yu, F.; White, S.B.; Zhao, Q.; Lee, F.S. HIF-1alpha binding to VHL is regulated by stimulus-sensitive proline hydroxylation. Proc. Natl. Acad. Sci. USA 2001, 98, 9630–9635. [Google Scholar] [CrossRef]
- Freeman, R.S.; Hasbani, D.M.; Lipscomb, E.A.; Straub, J.A.; Xie, L. SM-20, EGL-9, and the EGLN family of hypoxia-inducible factor prolyl hydroxylases. Mol. Cells 2003, 16, 1–12. [Google Scholar]
- Han, W.Q.; Zhu, Q.; Hu, J.; Li, P.L.; Zhang, F.; Li, N. Hypoxia-inducible factor prolyl-hydroxylase-2 mediates transforming growth factor beta 1-induced epithelial–mesenchymal transition in renal tubular cells. Biochim. Biophys. Acta 2013, 1833, 1454–1462. [Google Scholar] [CrossRef]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-Inducible Factors and the Response to Hypoxic Stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Yu, K.; Yang, M.; Almahi, W.A.; Nie, L.; Chen, G.; Han, W. Micheliolide Enhances Radiosensitivities of p53-Deficient Non-Small-Cell Lung Cancer via Promoting HIF-1α Degradation. Int. J. Mol. Sci. 2020, 21, 3392. [Google Scholar] [CrossRef] [PubMed]
- Freedman, S.J.; Sun, Z.-Y.J.; Poy, F.; Kung, A.L.; Livingston, D.M.; Wagner, G.; Eck, M.J. Structural basis for recruitment of CBP/p300 by hypoxia-inducible factor-1α. Proc. Natl. Acad. Sci. USA 2002, 99, 5367–5372. [Google Scholar] [CrossRef] [PubMed]
- Wenger, R.; Stiehl, D.; Camenisch, G. Integration of Oxygen Signaling at the Consensus HRE. Sci. STKE 2005, 2005, re12. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Munday, J.S.; Perrott, M.; Wang, G.; Liu, X. Association of age with the expression of hypoxia-inducible factors HIF-1α, HIF-2α, HIF-3α and VEGF in lung and heart of tibetan sheep. Animals 2019, 9, 673. [Google Scholar] [CrossRef]
- Jones, N.M.; Bergeron, M. Hypoxic preconditioning induces changes in HIF-1 target genes in neonatal rat brain. J. Cereb. Blood Flow Metab. 2001, 21, 1105–1114. [Google Scholar] [CrossRef]
- Bernaudin, M.; Tang, Y.; Reilly, M.; Petit, E.; Sharp, F.R. Brain genomic responses following hypoxia and re-oxygenation in neonatal rat. J. Biol. Chem. 2002, 277, 39728–39738. [Google Scholar] [CrossRef]
- Aragonés, J.; Fraisl, P.; Baes, M.; Carmeliet, P. Oxygen Sensors at the Crossroad of Metabolism. Cell Metab. 2009, 9, 11–22. [Google Scholar] [CrossRef]
- McMahon, S.; Charbonneau, M.; Grandmont, S.; Richard, D.; Dubois, C. Transforming growth factor beta1 induces hypoxia-inducible factor-1 stabilization through selective inhibition of PHD2 expression. J. Biol. Chem. 2006, 281, 24171–24181. [Google Scholar] [CrossRef]
- Chen, D.; Guo, X.; Nie, P. Phylogenetic studies of sinipercid fish (Perciformes: Sinipercidae) based on multiple genes, with first application of an immune-related gene, the virus-induced protein (viperin) gene. Mol. Phylogenetics Evol. 2010, 55, 1167–1176. [Google Scholar] [CrossRef]
- Liu, J.; Cui, Y.; Liu, J. Food consumption and growth of two piscivorous fishes, the mandarin fish and the Chinese snakehead. J. Fish Biol. 1998, 53, 1071–1083. [Google Scholar] [CrossRef]
- Duan, Y.; Dong, X.; Zhang, X.; Miao, Z. Effects of dissolved oxygen concentration and stocking density on the growth, energy budget and body composition of juvenile Japanese flounder, Paralichthys olivaceus (Temminck et Schlegel). Aquac. Res. 2010, 42, 407–416. [Google Scholar] [CrossRef]
- Richards, J.G.; Farrell, A.P.; Brauner, C.J. Fish Physiology: Hypoxia; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Geng, X.; Feng, J.; Liu, S.; Wang, Y.; Arias, C.; Liu, Z. Transcriptional regulation of hypoxia inducible factors alpha (HIF-α) and their inhibiting factor (FIH-1) of channel catfish (Ictalurus punctatus) under hypoxia. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 169, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Weng, S.; Shi, X.; Xu, X.; Shi, N.; He, J. Development of a mandarin fish Siniperca chuatsi fry cell line suitable for the study of infectious spleen and kidney necrosis virus (ISKNV). Virus Res. 2008, 135, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Evans, R.; O’Neill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Žídek, A.; Bates, P.; Blackwell, P.; Yim, P.; et al. Protein complex prediction with AlphaFold-Multimer. BioRxiv 2022. BioRxiv 2021–10. [Google Scholar]
- He, J.; Xie, T.-L.; Li, X.; Yu, Y.; Zhan, Z.-P.; Weng, S.-P.; Guo, C.-J. Molecular cloning of Y-Box binding protein-1 from mandarin fish and its roles in stress-response and antiviral immunity. Fish Shellfish. Immunol. 2019, 93, 406–415. [Google Scholar] [CrossRef]
- Xiao, W. The hypoxia signaling pathway and hypoxic adaptation in fishes. Sci. China Life Sci. 2015, 58, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Powell-Coffman, J.A. Genetic analysis of hypoxia signaling and response in C. Elegans Ann. N. Y. Acad. Sci. 2003, 995, 191–199. [Google Scholar] [CrossRef]
- Webster, K.A. Evolution of the coordinate regulation of glycolytic enzyme genes by hypoxia. J. Exp. Biol. 2003, 206, 2911–2922. [Google Scholar] [CrossRef]
- Semenza, G.L. Hydroxylation of HIF-1: Oxygen Sensing at the Molecular Level. Physiology 2004, 19, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Nettleton, D.; Jiang, M.; Kim, S.K.; Powell-Coffman, J.A. Roles of the HIF-1 Hypoxia-inducible Factor during Hypoxia Response in Caenorhabditis elegans. J. Biol. Chem. 2005, 280, 20580–20588. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Roy, A.; Baby, S.M.; Hoshi, T.; Semenza, G.L.; Prabhakar, N.R. Oxygen sensing in the body. Prog. Biophys. Mol. Biol. 2006, 91, 249–286. [Google Scholar] [CrossRef]
- Heise, K.; Puntarulo, S.; Nikinmaa, M.; Lucassen, M.; Pörtner, H.-O. Abele. Oxidative stress and HIF-1 DNA binding during stressful cold exposure and recovery in the North Sea eelpout (Zoarces viviparus). Comp. Biochem. Physiol. A 2006, 143, 494–503. [Google Scholar] [CrossRef]
- Rissanen, E.; Tranberg, H.K.; Nikinmaa, M. Oxygen availability regulates metabolism and gene expression in trout hepatocyte cultures. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1507–R1515. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, E.; Tranberg, H.K.; Sollid, J.; Nilsson, G.E.; Nikinmaa, M. Temperature regulates hypoxia-inducible factor-1 (HIF-1) in a poikilothermic vertebrate, crucian carp (Carassius carassius). J. Exp. Biol. 2006, 209, 994–1003. [Google Scholar] [CrossRef]
- Wang, H.; Huang, C.; Chen, N.; Zhu, K.; Chen, B.; Wang, W.; Wang, H. Molecular characterization and mRNA expression of HIF-prolyl hydroxylase-2 (phd2) in hypoxia-sensing pathways from Megalobrama amblycephala. Comp. Biochem. Physiol. Part B 2015, 186, 28–35. [Google Scholar] [CrossRef]
- Lin, X.H.; Ye, M.H.; Seatan, W.; Pan, Y.Y.; Chen, F.Y.; Huang, Y.; Zhu, C.H.; Li, G.L.; Tian, C.X. Sequence Characteristics and mRNA Expression Analysis of the phds Gene Family in Silver Sillago (Sillago sihama) under Hypoxia Stress. J. Guangdong Ocean. Univ. 2020, 40, 1–8. [Google Scholar]
- Li, X.; Zhang, M.; Ling, C.; Sha, H.; Zou, G.; Liang, H. Molecular Characterization and Response of Prolyl Hydroxylase Domain (PHD) Genes to Hypoxia Stress in Hypophthalmichthys molitrix. Animals 2022, 12, 131. [Google Scholar] [CrossRef]
- Pektas, S.; Knapp, M.J. Substrate preference of the HIF-prolyl hydroxylase-2 (PHD2) and substrate-induced conformational change. J. Inorg. Biochem. 2013, 126, 55–60. [Google Scholar] [CrossRef]
- Myllyharju, J. Prolyl 4-hydroxylases, key enzymes in the synthesis of collagens and regulation of the response to hypoxia, and their roles as treatment targets. Ann. Med. 2008, 40, 402–417. [Google Scholar] [CrossRef]
- Myllyharju, J. Prolyl 4-hydroxylases, master regulators of the hypoxia response. Acta Physiol. 2013, 208, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L.; Bels, D.D.; Corazza, F.; Kappler, M.; Eckert, A.W. Oxygen sensing, homeostasis, and disease. N. Engl. J. Med. 2011, 365, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Minamishima, Y.A.; Moslehi, J.; Padera, R.F.; Bronson, R.T.; Liao, R.; Kaelin, W.G. A Feedback Loop Involving the Phd3 Prolyl Hydroxylase Tunes the Mammalian Hypoxic Response In Vivo. Mol. Cell. Biol. 2009, 29, 5729–5741. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Wen, L.; Chen, Y. Molecular Cloning of phd1 and Comparative Analysis of phd1, 2, and 3 Expression in Xenopus laevis. Sci. World J. 2012, 2012, 689287. [Google Scholar] [CrossRef]
- Moslehi, J.; Minamishima, Y.A.; Shi, J.; Neuberg, D.M.; Charytan, D.; Padera, R.F.; Signoretti, S.; Liao, R.; Kaelin, W.G. Loss of Hypoxia-Inducible Factor Prolyl Hydroxylase Activity in Cardiomyocytes Phenocopies Ischemic Cardiomyopathy. Circulation 2010, 122, 1004–1016. [Google Scholar] [CrossRef]
- Loenarz, C.; Coleman, M.L.; Boleininger, A.; Schierwater, B.; Holland, P.W.H.; Ratcliffe, P.J.; Schofield, C.J. The hypoxia-inducible transcription factor pathway regulates oxygen sensing in the simplest animal, Trichoplax adhaerens. EMBO Rep. 2011, 12, 63–70. [Google Scholar] [CrossRef]
- Matthews, J.M.; Bhati, M.; Lehtomaki, E.; Mansfield, R.E.; Cubeddu, L.; Mackay, J.P. It Takes Two to Tango: The Structure and Function of LIM, RING, PHD and MYND Domains. Curr. Pharm. Des. 2009, 15, 3681–3696. [Google Scholar] [CrossRef]
- Edurne, B.; Emmanuel, B.; Amandine, G.; Véronique, V.; Danièle, R.; Jacques, P. HIF prolyl-hydroxylase 2 is the key oxygen sensor setting low steady-state levels of HIF-1alpha in normoxia. EMBO J. 2003, 22, 4082–4090. [Google Scholar]
- Shen, R.J.; Jiang, X.Y.; Pu, J.W.; Zou, S.M. HIF-1α and -2α genes in a hypoxia-sensitive teleost species Megalobrama amblycephala: cDNA cloning, expression and different responses to hypoxia. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 157, 273–280. [Google Scholar] [CrossRef]
- Lieb, M.E.; Menzies, K.; Moschella, M.C.; Ni, R.; Taubman, M.B. Mammalian EGLN genes have distinct patterns of mRNA expression and regulation. Biochem. Cell Biol. 2002, 80, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Appelhoff, R.J.; Tian, Y.-M.; Raval, R.R.; Turley, H.; Harris, A.L.; Pugh, C.W.; Ratcliffe, P.J.; Gleadle, J.M. Differential Function of the Prolyl Hydroxylases PHD1, PHD2, and PHD3 in the Regulation of Hypoxia-inducible Factor. J. Biol. Chem. 2004, 279, 38458–38465. [Google Scholar] [CrossRef] [PubMed]
- Groulx, I.; Lee, S. Oxygen-dependent ubiquitination and degradation of hypoxia-inducible factor requires nuclear-cytoplasmic trafficking of the von Hippel-Lindau tumor suppressor protein. Mol. Cell. Biol. 2002, 22, 5319–5336. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (For Initial PCR) | Sequences |
|---|---|
| 5′ RACE for scPHD2-F | 5′–CTAATAGCACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT–3′ |
| 5′ RACE for scPHD2-R | 5′–ACACTGTGATGGCATACCTGGTGGC–3′ |
| 3′ RACE for scPHD2-F | 5′–GGACTACGAGGCACCGGAGATAA–3′ |
| 3′ RACE for scPHD2-R | 5′–ACTCTGCGTTGATACCACTGCTTGCCCTATAGTGAGTGCTATTAG–3′ |
| scPHD2-F1 | 5′–ATGGAGAAGCAGCAGAGCGATTTGGAC–3′ |
| scPHD2-R1 | 5′–CTAGCTGGGATCTGATGGTTTGCCGA–3′ |
| Genes | Primers | Sequences | Primer Efficiency |
|---|---|---|---|
| scvegf | Forward Reverse | 5′–ACCGAAGGAAACAGAAAGAGG–3′ 5′–CAGGACGGGATGAAGATGTG–3′ | 0.98 |
| scldha | Forward Reverse | 5′–GGTCTTCCTGAGCATCCCTT–3′ 5′–TTCTCCTCTTCGGGCTTCA–3′ | 0.98 |
| scglut 1 | Forward Reverse | 5′–GGTTTATTGTGGCAGAGTTGTT–3′ 5′–CCCACTATGAAGTTGGCAGTC–3′ | 0.97 |
| β-actin | Forward Reverse | 5′–CCCTCTGAACCCCAAAGCCA–3′ 5′–CAGCCTGGATGGCAACGTACA–3′ | 0.96 |
| scphd2 | Forward Reverse | 5′–ACACCGCCACATCTAACG–3′ 5′–GTGCAGGGATTTGACATTCT–3′ | 0.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; He, J.; Liu, W.; Li, Z.; Weng, S.; He, J.; Guo, C. Molecular Characterization and Functional Analysis of Hypoxia-Responsive Factor Prolyl Hydroxylase Domain 2 in Mandarin Fish (Siniperca chuatsi). Animals 2023, 13, 1556. https://doi.org/10.3390/ani13091556
Yu Y, He J, Liu W, Li Z, Weng S, He J, Guo C. Molecular Characterization and Functional Analysis of Hypoxia-Responsive Factor Prolyl Hydroxylase Domain 2 in Mandarin Fish (Siniperca chuatsi). Animals. 2023; 13(9):1556. https://doi.org/10.3390/ani13091556
Chicago/Turabian StyleYu, Yang, Jian He, Wenhui Liu, Zhimin Li, Shaoping Weng, Jianguo He, and Changjun Guo. 2023. "Molecular Characterization and Functional Analysis of Hypoxia-Responsive Factor Prolyl Hydroxylase Domain 2 in Mandarin Fish (Siniperca chuatsi)" Animals 13, no. 9: 1556. https://doi.org/10.3390/ani13091556
APA StyleYu, Y., He, J., Liu, W., Li, Z., Weng, S., He, J., & Guo, C. (2023). Molecular Characterization and Functional Analysis of Hypoxia-Responsive Factor Prolyl Hydroxylase Domain 2 in Mandarin Fish (Siniperca chuatsi). Animals, 13(9), 1556. https://doi.org/10.3390/ani13091556