
Dynamic Distribution of Skin Microorganisms in Donkeys at Different Ages and Various Sites of the Body


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
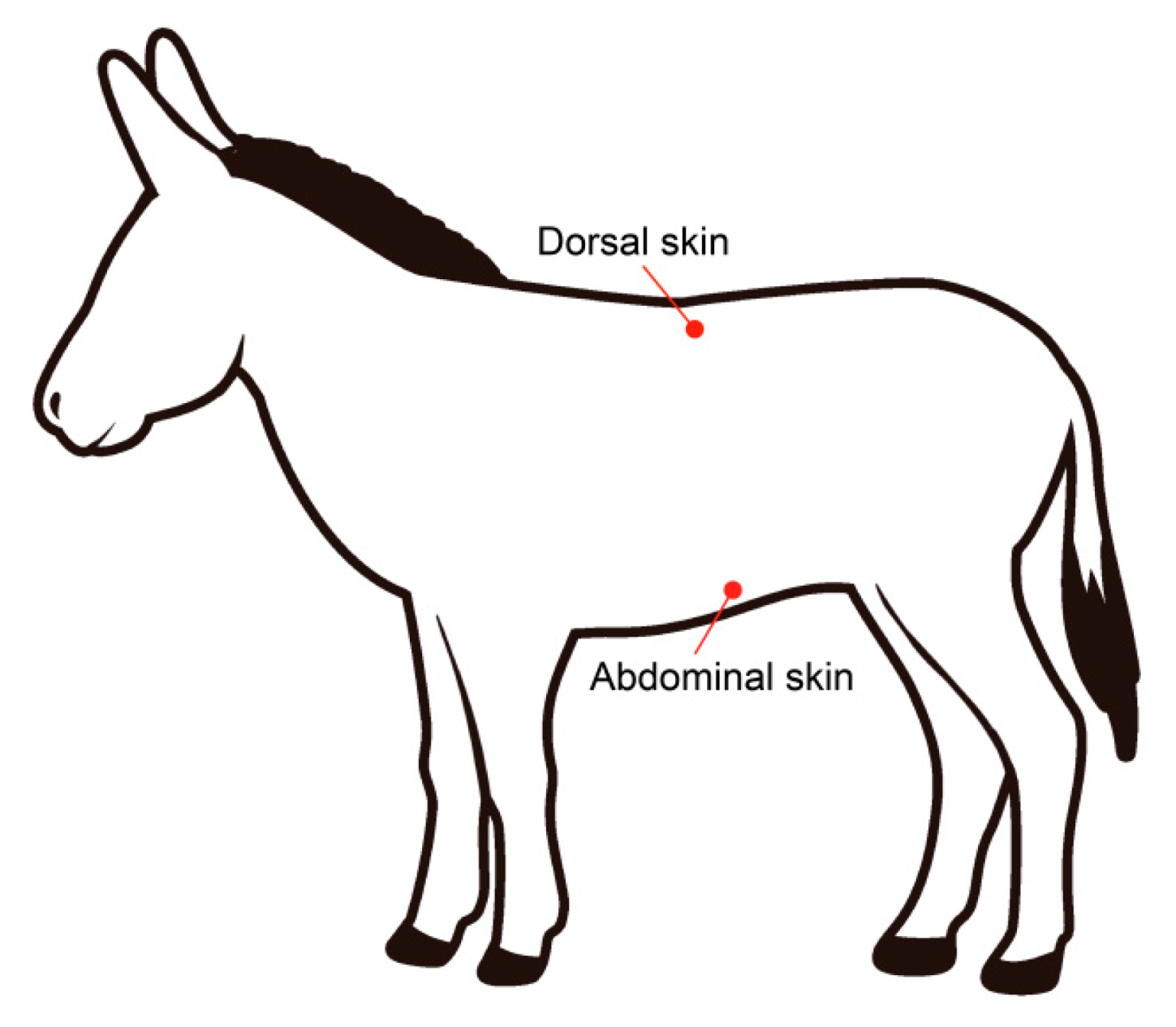
2.2. Sample Collection
2.3. DNA Extraction and PCR Amplification
2.4. Analysis of 16S and ITS Amplicon Sequencing Data
2.5. Statistical Analysis
3. Results
3.1. Skin Microbiome Structure of Donkeys Aged 1 Month to 4 Years
3.2. Age-Related Differences in Alpha Diversity
3.3. Age-Related Differences in Beta Diversity
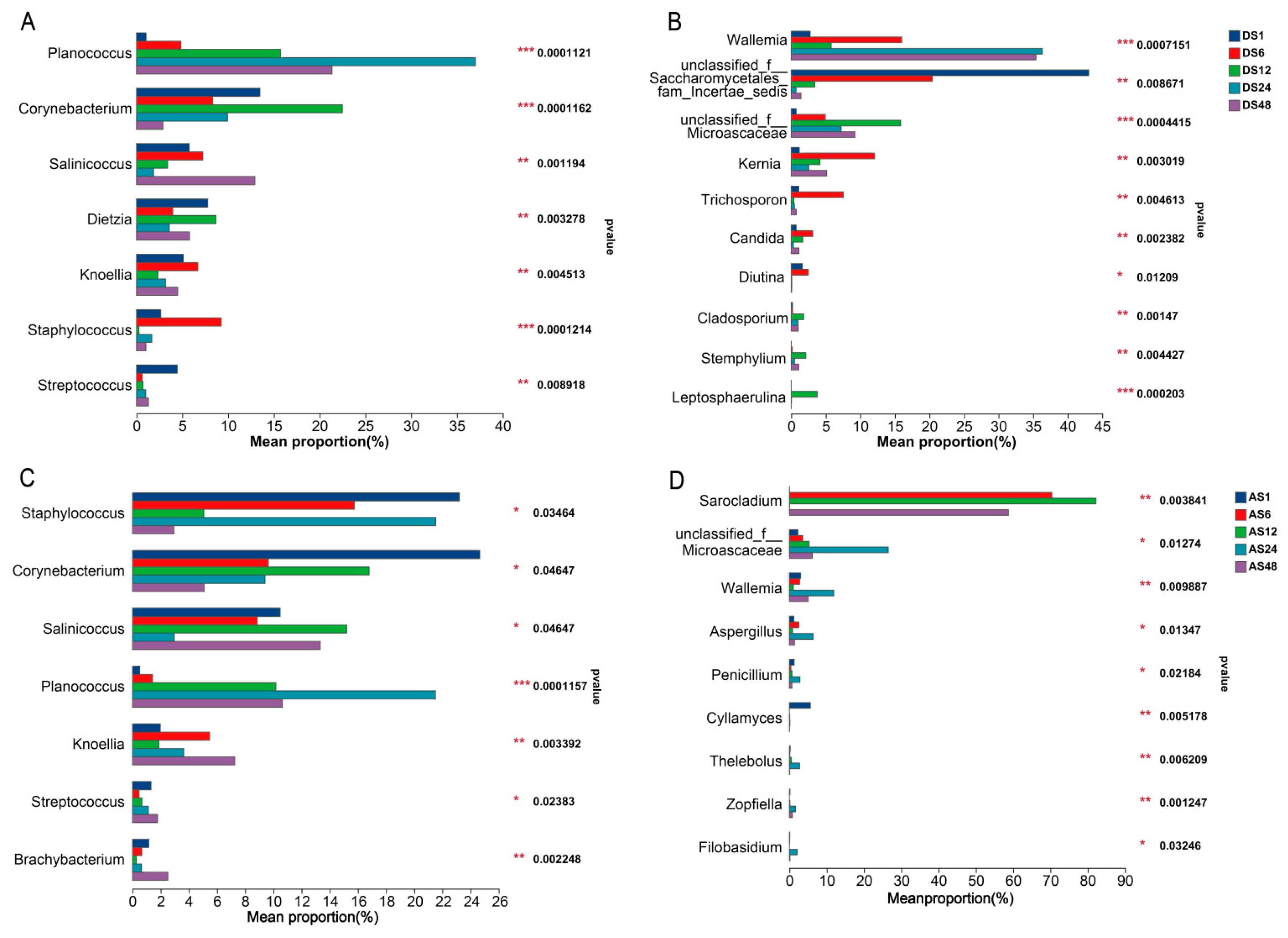
3.4. Microbial Community Composition Differences in Donkey Skin with Age
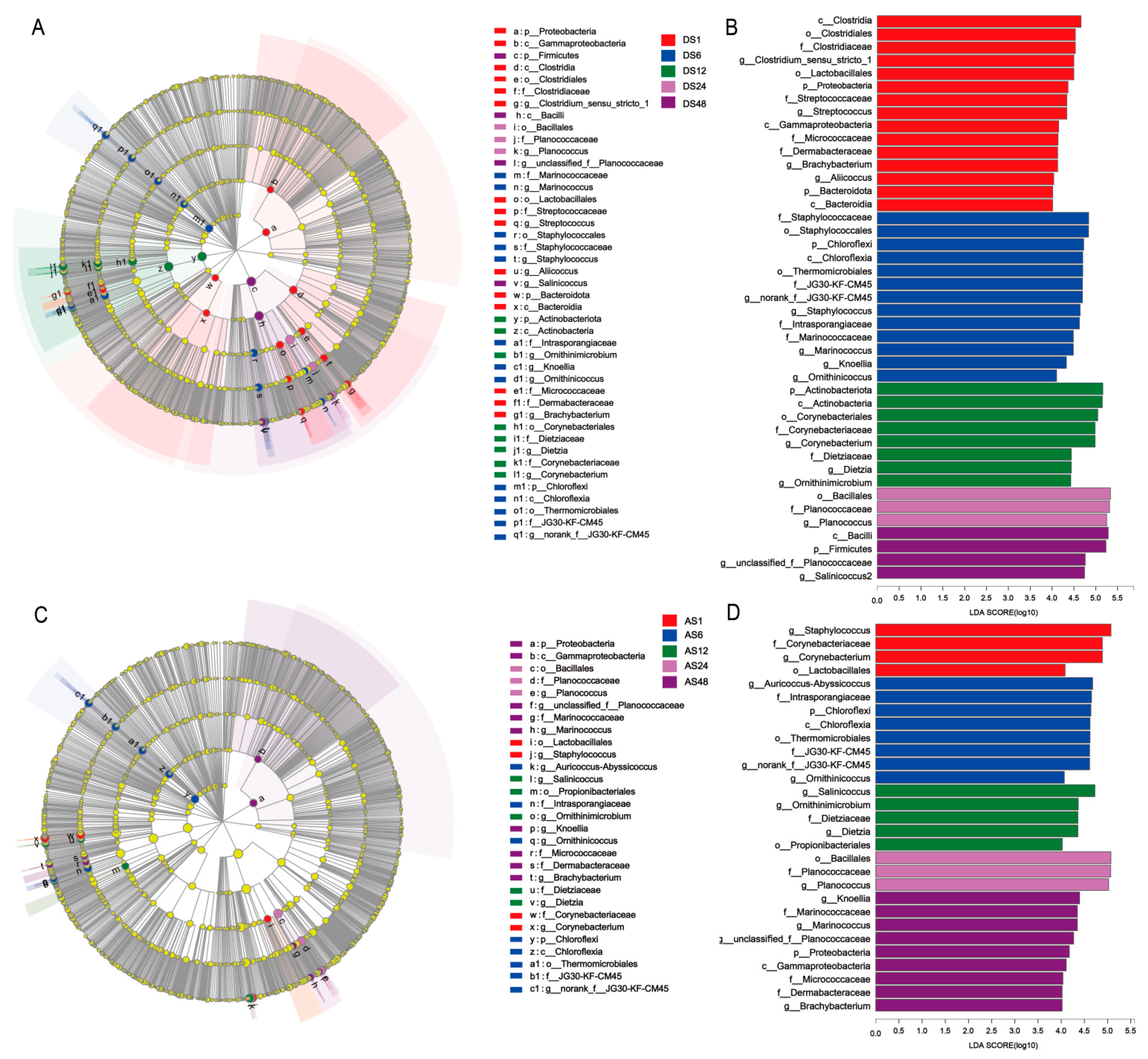
3.5. Taxonomic and Functional Changes in Skin Microbiomes among the Five Age Groups
3.6. Bacterial and Fungal Functional Characteristics
3.7. Correlations between the Skin Microbiota and Breeding Environment Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sanford, J.A.; Gallo, R.L. Functions of the skin microbiota in health and disease. Semin. Immunol. 2013, 25, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Saito, M.; Sugita, T.; Tsuboi, R. Malassezia species and their associated skin diseases. J. Dermatol. 2015, 42, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Barrio, E.; Rickards, K.J.; Thiemann, A.K. Clinical Evaluation and Preventative Care in Donkeys. Vet. Clin. N. Am. Equine Pract. 2019, 35, 545–560. [Google Scholar] [CrossRef] [PubMed]
- White, S.D.; Bourdeau, P.J.; Brément, T.; Vandenabeele, S.I.; Haspeslagh, M.; Bruet, V.; van Oldruitenborgh-Oosterbaan, M.M.S. Skin disease in donkeys (Equus asinus): A retrospective study from four veterinary schools. Vet. Dermatol. 2019, 30, 247-e76. [Google Scholar] [CrossRef] [PubMed]
- Knottenbelt, D.C. Skin Disorders of the Donkey and Mule. Vet. Clin. N. Am. Equine Pract. 2019, 35, 493–514. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Hospenthal, D.R. Uncommon Fungi and Related Species. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2015; pp. 3003–3015.e3003. [Google Scholar]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Lina, G.; Boutite, F.; Tristan, A.; Bes, M.; Etienne, J.; Vandenesch, F. Bacterial competition for human nasal cavity colonization: Role of Staphylococcal agr alleles. Appl. Environ. Microbiol. 2003, 69, 18–23. [Google Scholar] [CrossRef]
- Uçkay, I.; Pittet, D.; Vaudaux, P.; Sax, H.; Lew, D.; Waldvogel, F. Foreign body infections due to Staphylococcus epidermidis. Ann. Med. 2009, 41, 109–119. [Google Scholar] [CrossRef]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef]
- Ikhuoso, O.A.; Monroy, J.C.; Rivas-Caceres, R.R.; Cipriano-Salazar, M.; Barbabosa Pliego, A. Streptococcus equi in Equine: Diagnostic and Healthy Performance Impacts. J. Equine Vet. Sci. 2020, 85, 102870. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mitra, R.; Maitra, A.; Gupta, S.; Kumaran, S.; Chakrabortty, A.; Majumder, P.P. Sebum and Hydration Levels in Specific Regions of Human Face Significantly Predict the Nature and Diversity of Facial Skin Microbiome. Sci. Rep. 2016, 6, 36062. [Google Scholar] [CrossRef]
- Jugé, R.; Rouaud-Tinguely, P.; Breugnot, J.; Servaes, K.; Grimaldi, C.; Roth, M.P.; Coppin, H.; Closs, B. Shift in skin microbiota of Western European women across aging. J. Appl. Microbiol. 2018, 125, 907–916. [Google Scholar] [CrossRef]
- Wu, L.; Zeng, T.; Deligios, M.; Milanesi, L.; Langille, M.G.I.; Zinellu, A.; Rubino, S.; Carru, C.; Kelvin, D.J. Age-Related Variation of Bacterial and Fungal Communities in Different Body Habitats across the Young, Elderly, and Centenarians in Sardinia. mSphere 2020, 5, e00558-19. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Olejniczak-Staruch, I.; Ciążyńska, M.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibińska, M.; Lesiak, A. Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis. Int. J. Mol. Sci. 2021, 22, 3998. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Wanke, I.; Steffen, H.; Christ, C.; Krismer, B.; Götz, F.; Peschel, A.; Schaller, M.; Schittek, B. Skin commensals amplify the innate immune response to pathogens by activation of distinct signaling pathways. J. Investig. Dermatol. 2011, 131, 382–390. [Google Scholar] [CrossRef]
- Lai, Y.; Cogen, A.L.; Radek, K.A.; Park, H.J.; Macleod, D.T.; Leichtle, A.; Ryan, A.F.; Di Nardo, A.; Gallo, R.L. Activation of TLR2 by a small molecule produced by Staphylococcus epidermidis increases antimicrobial defense against bacterial skin infections. J. Investig. Dermatol. 2010, 130, 2211–2221. [Google Scholar] [CrossRef]
- Lima-Junior, D.S.; Krishnamurthy, S.R.; Bouladoux, N.; Collins, N.; Han, S.-J.; Chen, E.Y.; Constantinides, M.G.; Link, V.M.; Lim, A.I.; Enamorado, M.; et al. Endogenous retroviruses promote homeostatic and inflammatory responses to the microbiota. Cell 2021, 184, 3794–3811.e19. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, Q.; Meng, H.; Lv, H.; Liu, Y.; Liu, J.; Wang, H.; He, L.; Qin, J.; Wang, Y.; et al. Staphylococcus epidermidis Contributes to Healthy Maturation of the Nasal Microbiome by Stimulating Antimicrobial Peptide Production. Cell Host Microbe 2020, 27, 68–78.e65. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Jiang, W.; Holyoak, R.; Liu, B.; Li, J. Investigation of Oral Microbiome in Donkeys and the Effect of Dental Care on Oral Microbial Composition. Animals 2020, 10, 2245. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Huang, B.; Zhu, M.; Wang, C. The Fibrolytic Enzyme Profiles and the Composition of Fungal Communities in Donkey Cecum-Colon Ecosystem. Animals 2022, 12, 412. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, R.A.; Elshahed, M.S.; Youssef, N.H. Feramyces austinii, gen. nov., sp. nov., an anaerobic gut fungus from rumen and fecal samples of wild Barbary sheep and fallow deer. Mycologia 2018, 110, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.A. Purification and Characterisation of a Secreted Glycosidase, from the Extreme Xerophile Wallemia Ichthyophaga: A Thesis Presented in Partial Fulfilment of the Requirement for the Degree in Master of Science in Biochemistry at Massey University Palmerston North, New Zealand. Master’s Thesis, Massey University, Palmerston North, New Zealand, 2014. [Google Scholar]
- DeMers, M. Alternaria alternata as endophyte and pathogen. Microbiology 2022, 168, 001153. [Google Scholar] [CrossRef]
- Cao, M.; Fatma, Z.; Song, X.; Hsieh, P.H.; Tran, V.G.; Lyon, W.L.; Sayadi, M.; Shao, Z.; Yoshikuni, Y.; Zhao, H. A genetic toolbox for metabolic engineering of Issatchenkia orientalis. Metab. Eng. 2020, 59, 87–97. [Google Scholar] [CrossRef]
- Júnior, M.C.; de Moraes Arantes, A.; Silva, H.M.; Costa, C.R.; Silva, M.d.R. Acremonium kiliense: Case report and review of published studies. Mycopathologia 2013, 176, 417–421. [Google Scholar] [CrossRef]
- Etienne, K.A.; Roe, C.C.; Smith, R.M.; Vallabhaneni, S.; Duarte, C.; Escadon, P.; Castaneda, E.; Gomez, B.L.; de Bedout, C.; López, L.F.; et al. Whole-Genome Sequencing to Determine Origin of Multinational Outbreak of Sarocladium kiliense Bloodstream Infections. Emerg. Infect. Dis. 2016, 22, 476–481. [Google Scholar] [CrossRef]
- Ayla, S.; Golla, N.; Pallipati, S. Production of ligninolytic enzymes from sp. and its efficiency to decolourise textile dyes. Open Biotechnol. J. 2018, 12, 112–122. [Google Scholar] [CrossRef]
- Singh, P.; Roy, U.; Tsuji, M. Characterisation of yeast and filamentous fungi from Brøggerbreen glaciers, Svalbard. Polar Rec. 2016, 52, 442–449. [Google Scholar] [CrossRef]
- Ederveen, T.H.A.; Smits, J.P.H.; Boekhorst, J.; Schalkwijk, J.; van den Bogaard, E.H.; Zeeuwen, P. Skin microbiota in health and disease: From sequencing to biology. J. Dermatol. 2020, 47, 1110–1118. [Google Scholar] [CrossRef]
- Xu, L.L.; Chen, H.; Zhang, M.; Zhu, W.; Chang, Q.; Lu, G.; Chen, Y.; Jiang, J.; Zhu, L. Changes in the community structure of the symbiotic microbes of wild amphibians from the eastern edge of the Tibetan Plateau. MicrobiologyOpen 2020, 9, e1004. [Google Scholar] [CrossRef]
- Percival, S.L.; Emanuel, C.; Cutting, K.F.; Williams, D.W. Microbiology of the skin and the role of biofilms in infection. Int. Wound J. 2012, 9, 14–32. [Google Scholar] [CrossRef]
- Hussain, A.; Ansari, A.; Ahmad, R. Chapter 4—Microbial biofilms: Human mucosa and intestinal microbiota. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Biofilms; Yadav, M.K., Singh, B.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 47–60. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Callewaert, C.; Ravard Helffer, K.; Lebaron, P. Skin Microbiome and its Interplay with the Environment. Am. J. Clin. Dermatol. 2020, 21, 4–11. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | DS | AS | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 Month | 6 Months | 12 Months | 24 Months | 48 Months | 1 Month | 6 Months | 12 Months | 24 Months | 48 Months | ||
| Bacteria | sobs | 1285.17 ± 29.80 a | 1042.17 ± 63.59 b | 802.83 ± 68.20 c | 757.83 ± 61.29 c | 1069.17 ± 65.18 b | 982.67 ± 140.40 b | 921.33 ± 89.30 b | 884.17 ± 63.26 b | 847.67 ± 47.71 b | 1287.33 ± 82.02 a |
| ACE | 1896.90 ± 153.95 ab | 1963.50 ± 97.40 a | 1586.37 ± 116.74 b | 1573.74 ± 104.50 b | 2204.33 ± 86.12 a | 1680.27 ± 179.90 b | 1730.79 ± 108.02 b | 1754.32 ± 84.43 b | 1652.48 ± 75.89 b | 2087.35 ± 69.63 a | |
| Chao | 1734.37 ± 90.11 a | 1588.41 ± 76.60 a | 1284.37 ± 88.29 b | 1223.46 ± 89.23 b | 1781.74 ± 76.04 a | 1416.23 ± 138.63 b | 1423.90 ± 98.71 b | 1415.06 ± 93.76 b | 1368.06 ± 64.78 b | 1884.82 ± 64.79 a | |
| Shannon | 4.75 ± 0.06 a | 4.32 ± 0.19 ab | 3.84 ± 0.19 c | 3.39 ± 0.17 d | 4.05 ± 0.11 bc | 3.23 ± 0.48 b | 3.76 ± 0.34 ab | 3.71 ± 0.35 ab | 3.49 ± 0.17 ab | 4.40 ± 0.10 a | |
| Simpson | 0.03 ± 0.00 c | 0.04 ± 0.01 bc | 0.07 ± 0.02 b | 0.13 ± 0.02 a | 0.05 ± 0.01 bc | 0.24 ± 0.07 a | 0.08 ± 0.02 b | 0.11 ± 0.05 ab | 0.12 ± 0.03 ab | 0.04 ± 0.00 b | |
| Fungus | sobs | 135.17 ± 35.90 c | 337.33 ± 14.14 ab | 325.50 ± 21.48 ab | 369.33 ± 43.43 a | 277.83 ± 10.94 b | 199.33 ± 58.08 a | 197.83 ± 29.86 a | 90.17 ± 17.62 b | 200.17 ± 17.71 a | 174.17 ± 12.02 ab |
| ACE | 171.53 ± 48.16 c | 424.95 ± 14.91 ab | 376.29 ± 29.97 ab | 448.60 ± 58.15 a | 318.76 ± 15.30 b | 233.73 ± 68.43 | 250.60 ± 35.00 | 192.71 ± 19.46 | 221.37 ± 15.05 | 208.11 ± 11.98 | |
| Chao | 171.10 ± 50.13 c | 417.70 ± 16.45 ab | 377.50 ± 29.59 ab | 448.69 ± 57.52 a | 319.05 ± 15.05 b | 230.40 ± 68.58 | 249.74 ± 34.58 | 136.98 ± 19.78 | 225.36 ± 17.84 | 206.71 ± 10.53 | |
| Shannon | 1.80 ± 0.31 b | 3.19 ± 0.08 a | 2.76 ± 0.34 a | 2.86 ± 0.24 a | 3.02 ± 0.12 a | 2.95 ± 0.37 a | 1.34 ± 0.46 b | 0.63 ± 0.51 b | 3.21 ± 0.18 a | 1.63 ± 0.58 b | |
| Simpson | 0.39 ± 0.08 a | 0.09 ± 0.01 b | 0.23 ± 0.08 b | 0.17 ± 0.03 b | 0.13 ± 0.02 b | 0.16 ± 0.05 b | 0.62 ± 0.13 a | 0.83 ± 0.14 a | 0.13 ± 0.03 b | 0.55 ± 0.16 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Yue, Y.; Kou, X.; Hou, W.; Wang, M.; Yang, X.; Liu, G.; Li, Y.; Wang, C. Dynamic Distribution of Skin Microorganisms in Donkeys at Different Ages and Various Sites of the Body. Animals 2023, 13, 1566. https://doi.org/10.3390/ani13091566
Ma Q, Yue Y, Kou X, Hou W, Wang M, Yang X, Liu G, Li Y, Wang C. Dynamic Distribution of Skin Microorganisms in Donkeys at Different Ages and Various Sites of the Body. Animals. 2023; 13(9):1566. https://doi.org/10.3390/ani13091566
Chicago/Turabian StyleMa, Qingshan, Yunshuang Yue, Xiyan Kou, Wanting Hou, Mingyu Wang, Xihao Yang, Guiqin Liu, Yan Li, and Changfa Wang. 2023. "Dynamic Distribution of Skin Microorganisms in Donkeys at Different Ages and Various Sites of the Body" Animals 13, no. 9: 1566. https://doi.org/10.3390/ani13091566
APA StyleMa, Q., Yue, Y., Kou, X., Hou, W., Wang, M., Yang, X., Liu, G., Li, Y., & Wang, C. (2023). Dynamic Distribution of Skin Microorganisms in Donkeys at Different Ages and Various Sites of the Body. Animals, 13(9), 1566. https://doi.org/10.3390/ani13091566