
The Detection of Circulating Antigen Glutathione S-Transferase in Sheep Infected with Fasciola hepatica with Double-Antibody Sandwich Signal Amplification Enzyme-Linked Immunosorbent Assay

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum and Fecal Samples
2.2. Screening of Candidate Detection Antigen Genes of F. hepatica
2.3. Expression, Purification, and Identification of the Recombinant Protein rFhGST
2.4. Preparation of the Anti-rFhGST Polyclonal Antibody
2.5. Immunolocalization of FhGST in F. hepatica
2.6. Preparation of the Anti-rFhGST Monoclonal Antibody
2.7. Preparation and Optimization of Biotin-Labeled Rabbit Anti-rFhGST Polyclonal Antibody
2.8. Establishment of the SA-ELISA
2.9. Performance Test of the SA-ELISA
2.10. Comparison with Fecal Test
2.11. Comparison with Commercial Kits
2.12. Application of the SA-ELISA
2.13. Statistical Analysis
3. Results
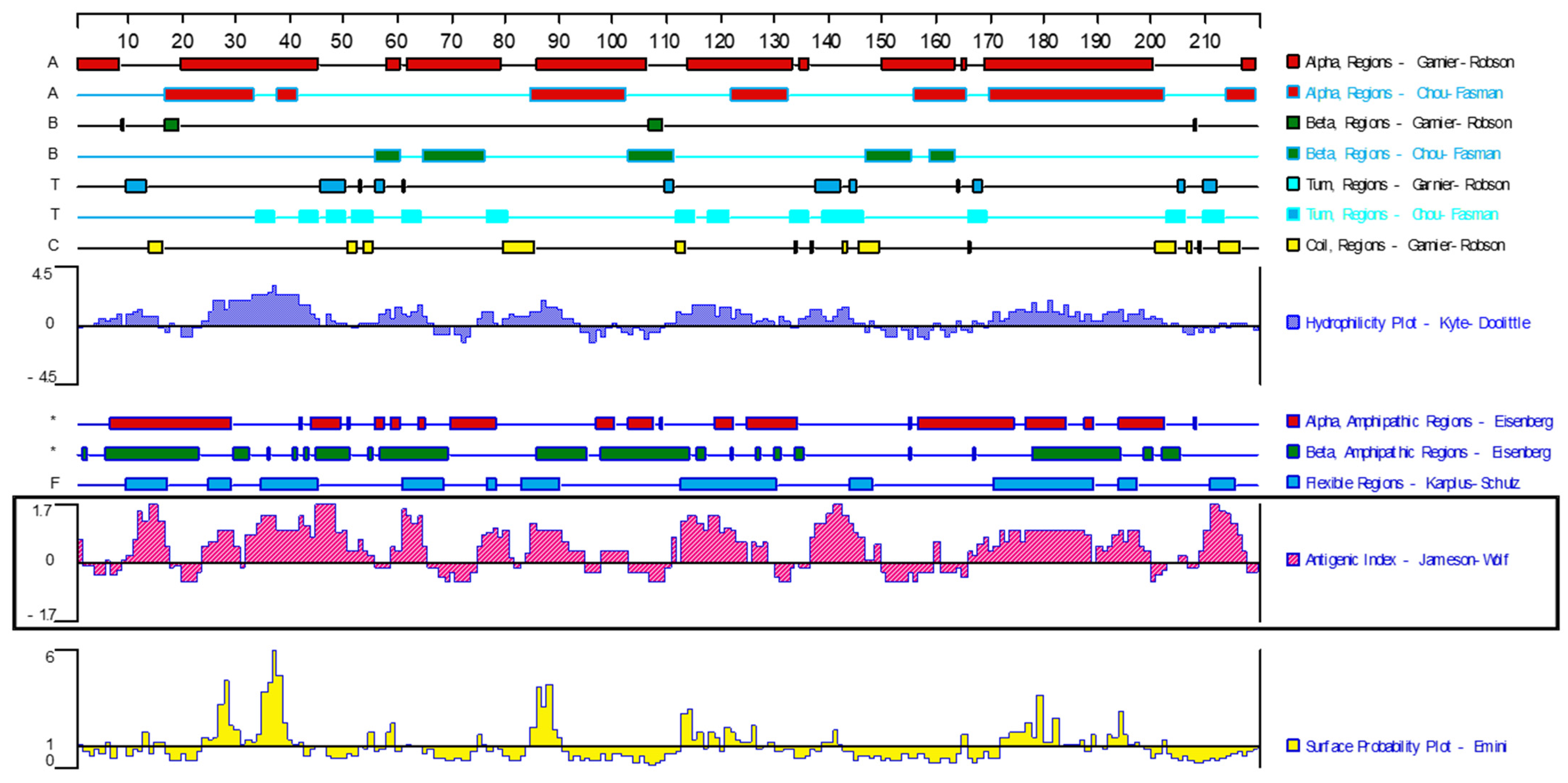
3.1. Screening of Immunodominant Antigen Genes of F. hepatica
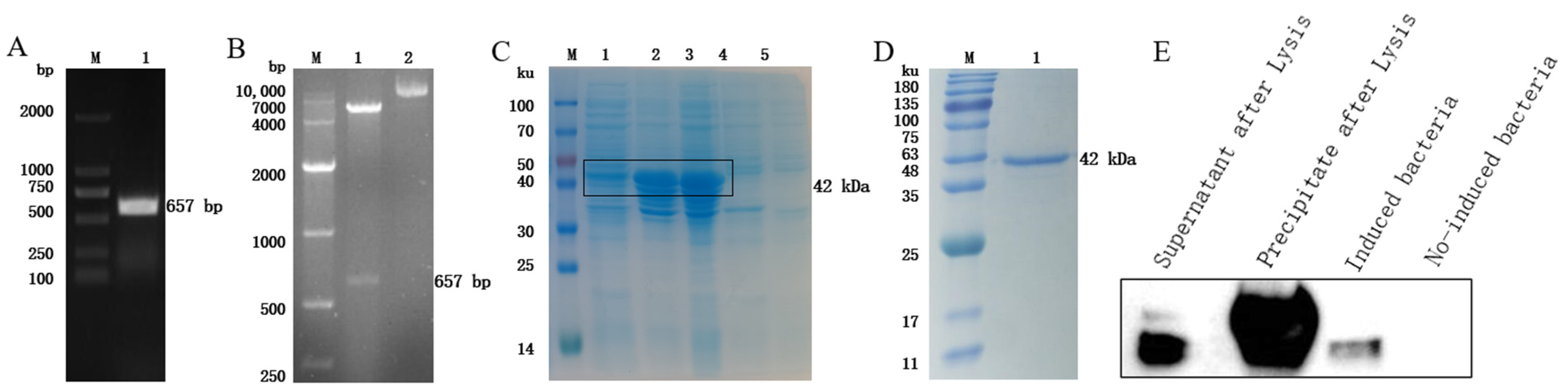
3.2. Expression, Purification, and Identification of the Recombinant Protein rFhGST
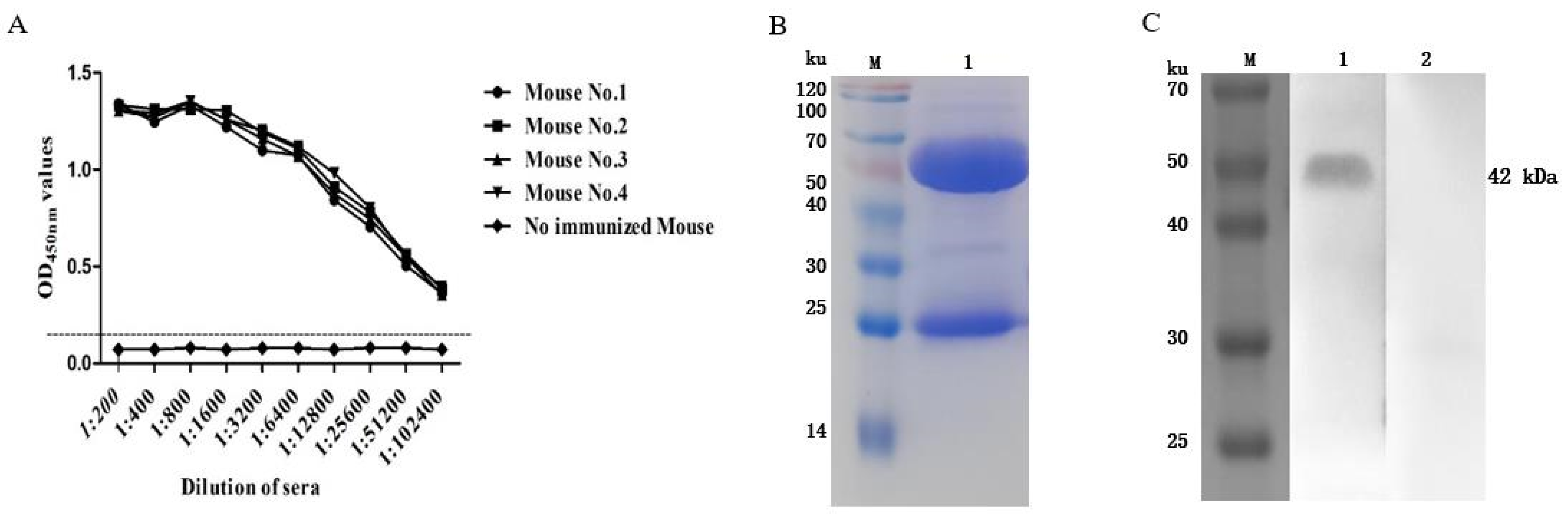
3.3. Preparation of the Anti-rFhGST Polyclonal Antibody
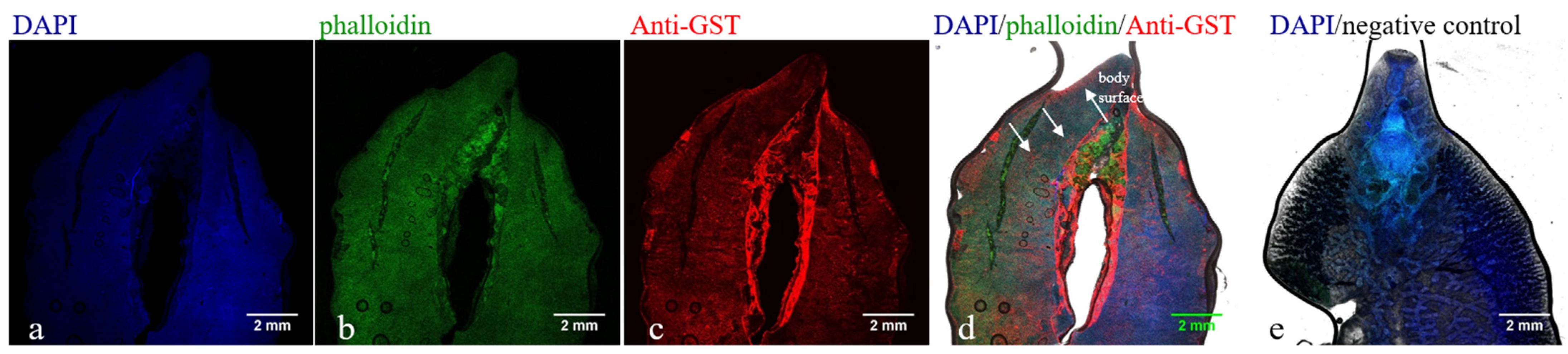
3.4. Immunolocalization of FhGST in F. hepatica
3.5. Preparation of the Mouse Anti-rFhGST Monoclonal Antibody
3.6. Optimization of the Molar Feed Ratio of the Rabbit Anti-rFhGST Polyclonal Antibody to BNHS
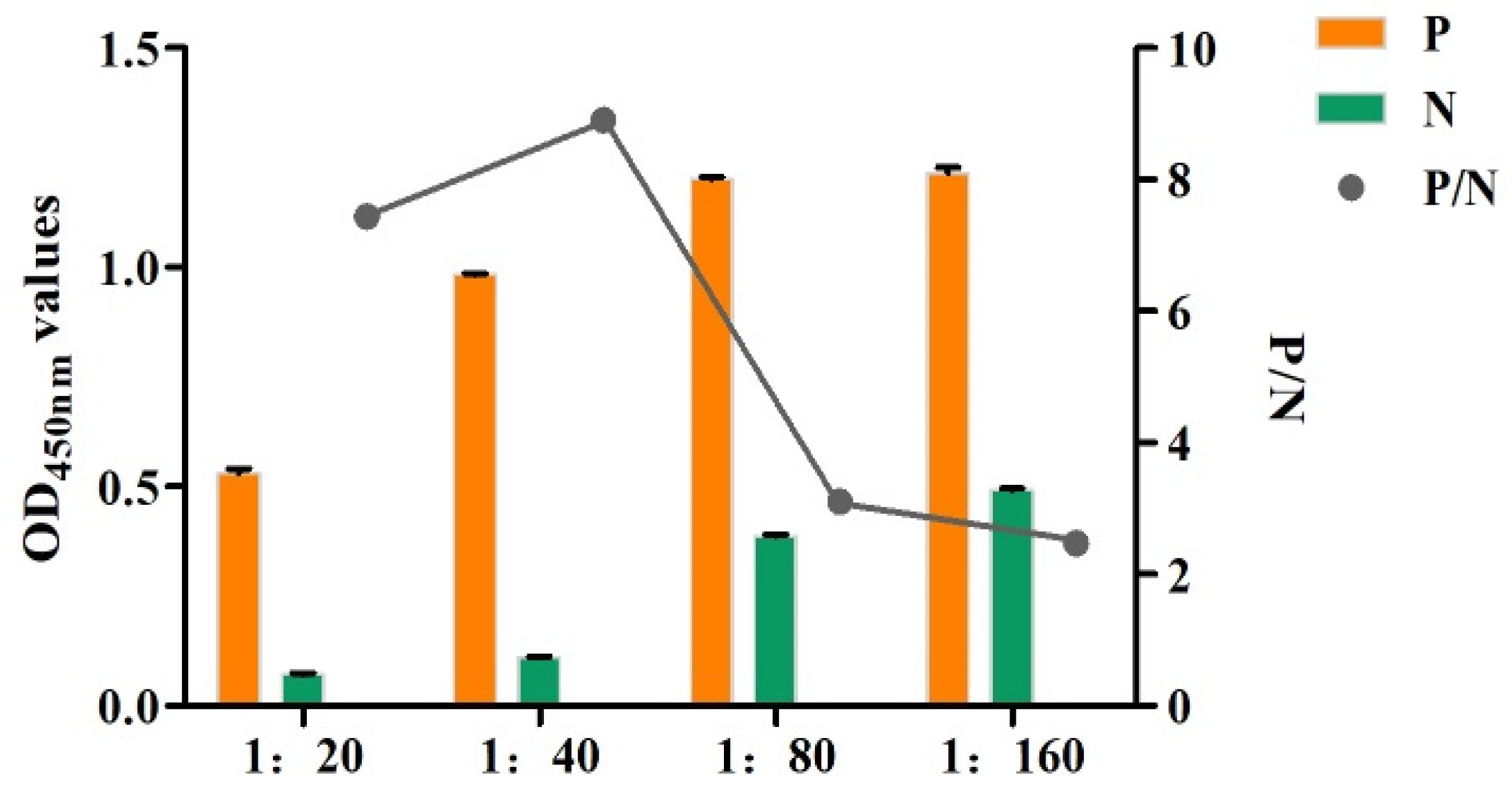
3.7. Establishment and Optimization of the SA-ELISA
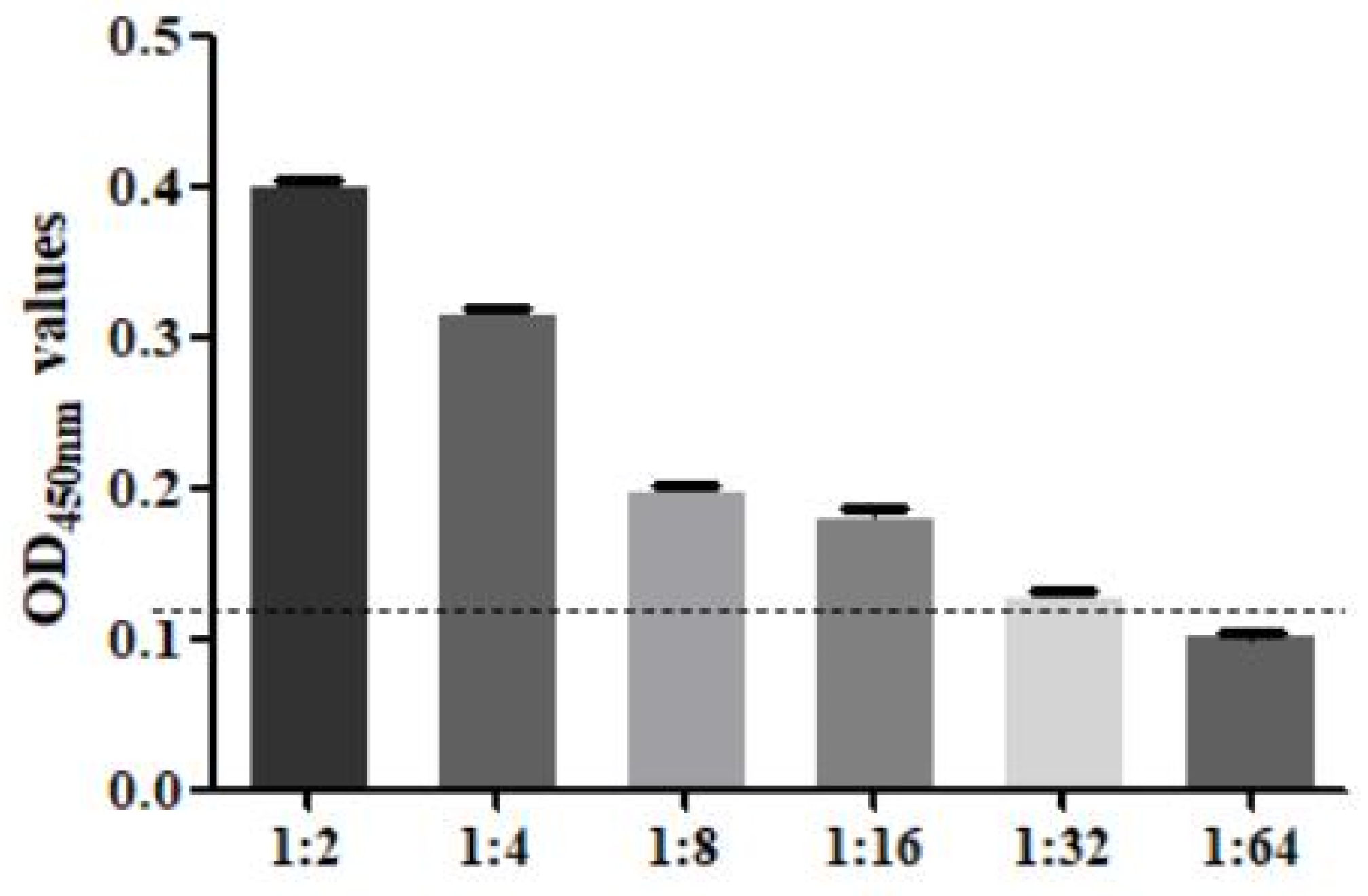
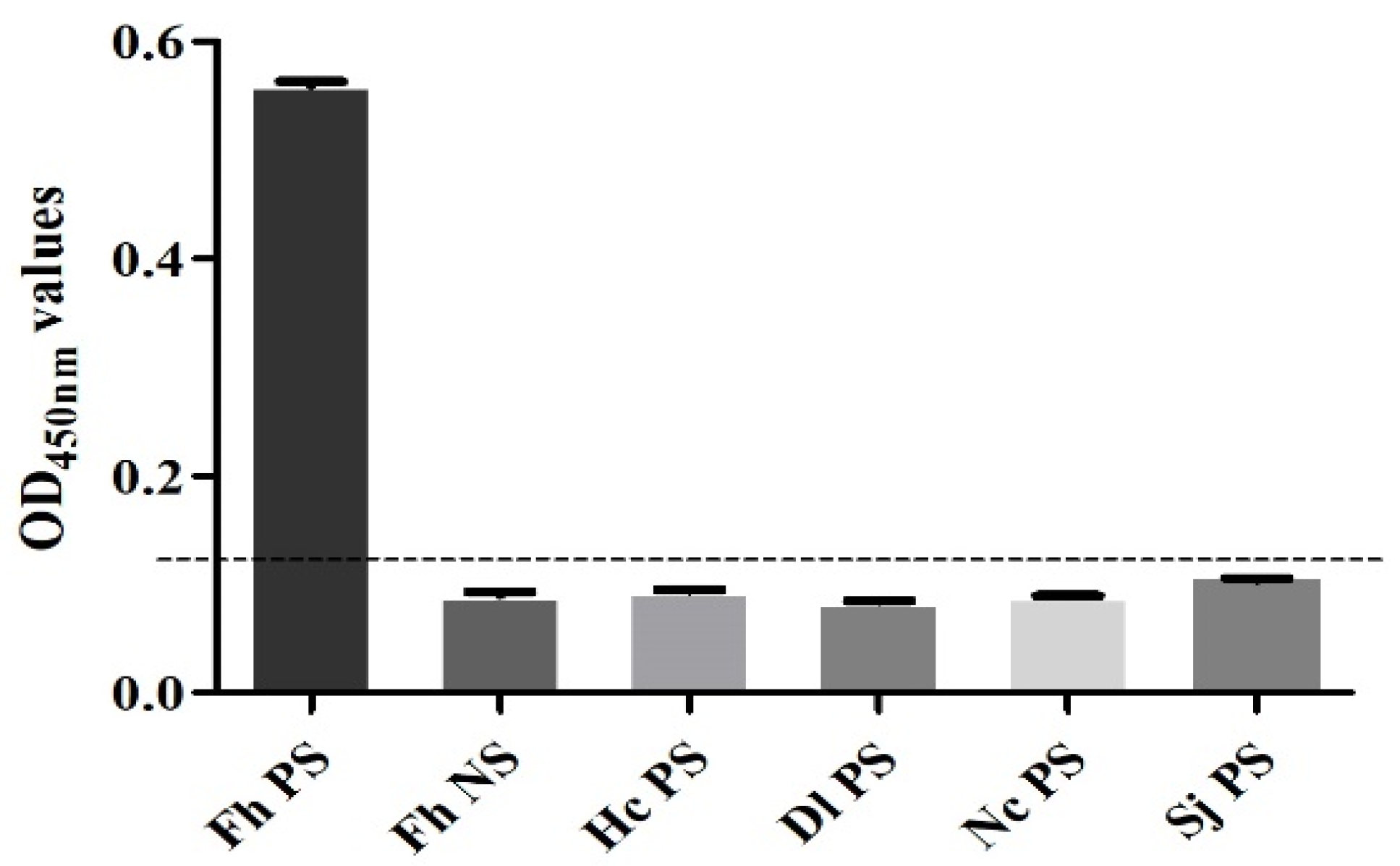
3.8. Performance Test of the SA-ELISA
3.9. Comparison with Fecal Test
3.10. Comparison with Commercial Kit Test Results
3.11. Application of the SA-ELISA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Afshan, K.; Valero, M.A.; Qayyum, M.; Peixoto, R.V.; Magraner, A.; Mas-Coma, S. Phenotypes of intermediate forms of Fasciola hepatica and F. gigantica in buffaloes from Central Punjab, Pakistan. J. Helminthol. 2014, 88, 417–426. [Google Scholar] [CrossRef]
- Dolay, K.; Hasbahçeci, M.; Hatipoğlu, E.; Ümit Malya, F.; Akçakaya, A. Endoscopic diagnosis and treatment of biliary obstruction due to acute cholangitis and acute pancreatitis secondary to Fasciola hepatica infection. Ulus. Travma Acil Cerrahi Derg. 2018, 24, 71–73. [Google Scholar]
- Lefryekh, R.; Bensaad, A.; Bensardi, F.; Elhattabi, K.; Bouali, M.; Daif, B.; Fadil, A.; Jaouhari, Z.; Hicham, T.; Hamdani, A.; et al. Hepatic fascioliasis presenting with bile duct obstruction: A case report. Pan Afr. Med. J. 2017, 28, 44. [Google Scholar] [PubMed]
- Rawla, P.; Bandaru, S.S.; Vellipuram, A.R. Review of Infectious Etiology of Acute Pancreatitis. Gastroenterol. Res. 2017, 10, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Sumruayphol, S.; Siribat, P.; Dujardin, J.P.; Dujardin, S.; Komalamisra, C.; Thaenkham, U. Fasciola gigantica, F. hepatica and Fasciola intermediate forms: Geometric morphometrics and an artificial neural network to help morphological identification. PeerJ 2020, 8, e8597. [Google Scholar] [CrossRef] [PubMed]
- Fürst, T.; Duthaler, U.; Sripa, B.; Utzinger, J.; Keiser, J. Trematode infections: Liver and lung flukes. Infect. Dis. Clin. N. Am. 2012, 26, 399–419. [Google Scholar] [CrossRef] [PubMed]
- Khoramian, H.; Arbabi, M.; Osqoi, M.M.; Delavari, M.; Hooshyar, H.; Asgari, M. Prevalence of ruminants fascioliasis and their economic effects in Kashan, center of Iran. Asian Pac. J. Trop. Biomed. 2014, 4, 918–922. [Google Scholar] [CrossRef]
- Mehmood, K.; Zhang, H.; Sabir, A.J.; Abbas, R.Z.; Ijaz, M.; Durrani, A.Z.; Saleem, M.H.; Ur Rehman, M.; Iqbal, M.K.; Wang, Y.; et al. A review on epidemiology, global prevalence and economical losses of fasciolosis in ruminants. Microb. Pathog. 2017, 109, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Dalton, J.P. Omics tools enabling vaccine discovery against fasciolosis. Trends Parasitol. 2022, 38, 1068–1079. [Google Scholar] [CrossRef]
- Kelley, J.M.; Elliott, T.P.; Beddoe, T.; Anderson, G.; Skuce, P.; Spithill, T.W. Current Threat of Triclabendazole Resistance in Fasciola hepatica. Trends Parasitol. 2016, 32, 458–469. [Google Scholar] [CrossRef]
- Heidari, H.; Zahiri, H.; Gharekhani, J.; Hosseini, A.; Aeineh, S. Comparison of Dot-ELISA and ELISA Techniques for Detection of Fasciola hepatica in Sheep Using Excretory-Secretory Antigens. Istanb. Univ. Vet. Fak. Dergisi. 2015, 41, 21–25. [Google Scholar]
- Khabisi, S.A.; Sarkari, B.; Moshfe, A.; Jalali, S. Production of Monoclonal Antibody Against Excretory-Secretory Antigen of Fasciola hepatica and Evaluation of Its Efficacy in the Diagnosis of Fascioliasis. Monoclon. Antibodies Immunodiagn. Immunother. 2017, 65, 171–174. [Google Scholar]
- López Corrales, J.; Cwiklinski, K.; De Marco Verissimo, C.; Dorey, A.; Lalor, R.; Jewhurst, H.; McEvoy, A.; Diskin, M.; Duffy, C.; Cosby, S.L.; et al. Diagnosis of sheep fasciolosis caused by Fasciola hepatica using cathepsin L enzyme-linked immunosorbent assays (ELISA). Vet. Parasitol. 2021, 298, 109517. [Google Scholar] [CrossRef] [PubMed]
- Afshan, K.; Qayyum, M.; Rizvi, S.S.R.; Mukhtar, M.; Mushtaq, M.; Miller, J.E. Serological and coprological comparison for rapid diagnosis of Fasciola hepatica infection in small ruminants from sub-tropical area of Pakistan. Small Rumin. Res. 2013, 113, 267–272. [Google Scholar] [CrossRef]
- Mazeri, S.; Sargison, N.; Kelly, R.F.; Bronsvoort, B.M.D.; Handel, I. Evaluation of the Performance of Five Diagnostic Tests for Fasciola hepatica Infection in Naturally Infected Cattle Using a Bayesian No Gold Standard Approach. PLoS ONE 2016, 11, e0161621. [Google Scholar] [CrossRef]
- Espino, A.M.; Finlay, C.M. Sandwich enzyme-linked immunosorbent assay for detection of excretory secretory antigens in humans with fascioliasis. J. Clin. Microbiol. 1994, 32, 190–193. [Google Scholar] [CrossRef]
- Gordon, D.K.; Zadoks, R.N.; Stevenson, H.; Sargison, N.D.; Skuce, P.J. On farm evaluation of the coproantigen ELISA and coproantigen reduction test in Scottish sheep naturally infected with Fasciola hepatica. Vet. Parasitol. 2012, 187, 436–444. [Google Scholar] [CrossRef]
- Aghamolaei, S.; Kazemi, B.; Bandehpour, M.; Ranjbar, M.M.; Rouhani, S.; Javadi Mamaghani, A.; Tabaei, S.J.S. Design and expression of polytopic construct of cathepsin-L1, SAP-2 and FhTP16.5 proteins of Fasciola hepatica. J. Helminthol. 2020, 94, e134. [Google Scholar] [CrossRef]
- Mokhtarian, K.; Meamar, A.R.; Khoshmirsafa, M.; Razmjou, E.; Masoori, L.; Khanmohammadi, M.; Akhlaghi, L.; Falak, R. Comparative assessment of recombinant and native immunogenic forms of Fasciola hepatica proteins for serodiagnosis of sheep fasciolosis. Parasitol. Res. 2018, 117, 225–232. [Google Scholar] [CrossRef]
- Vasti, A.; Bianca, V.; Espino, A.M. Assessment of Fasciola hepatica glutathione S-transferase as an antigen for serodiagnosis of human chronic fascioliasis. Acta Trop. 2018, 186, 41–49. [Google Scholar]
- Xifeng, W.; Mengfan, Q.; Kai, Z.; Guowu, Z.; Xuepeng, C. Development and evaluation of a colloidal gold immunochromatographic assay based on recombinant protein CatL1D for serodiagnosis of sheep fasciolosis. J. Helminthol. 2019, 94, e98. [Google Scholar] [CrossRef]
- Anuracpreeda, P.; Chawengkirttikul, R.; Sobhon, P. Immunodiagnostic monoclonal antibody-based sandwich ELISA of fasciolosis by detection of Fasciola gigantica circulating fatty acid binding protein. Parasitology 2016, 143, 1369–1381. [Google Scholar] [CrossRef]
- Martínez-Sernández, V.; Orbegozo-Medina, R.A.; González-Warleta, M.; Mezo, M.; Ubeira, F.M. Rapid Enhanced MM3-COPRO ELISA for Detection of Fasciola Coproantigens. PLoS Negl. Trop. Dis. 2016, 10, e0004872. [Google Scholar] [CrossRef]
- Liu, T.; Li, X.; Gong, P.T.; Zhang, X.C.; Yang, J.; Li, H.; Li, J.H. Construction of a Fasciola hepatica cDNA library and screening of genes encoding potential diagnostic antigens. J. Pathog. Biol. 2019, 14, 27–31. [Google Scholar]
- Beatty, J.D.; Beatty, B.G.; Vlahos, W.G. Measurement of monoclonal antibody affinity by non-competitive enzyme immunoassay. J. Immunol. Methods 1987, 100, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, H.; Zheng, X.; Jiao, Y.; Ning, L.; Zhou, E.M.; Mu, Y. A Double-Antibody Sandwich ELISA for Sensitive and Specific Detection of Swine Fibrinogen-Like Protein 1. Front. Immunol. 2021, 12, 670626. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.J.; Board, P.G.; Boray, J.C. Glutathione S-transferases in Fasciola hepatica. J. Parasitol. 1988, 74, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Zerna, G.; Cameron, T.C.; Toet, H.; Spithill, T.W.; Beddoe, T. Bovine Natural Antibody Relationships to Specific Antibodies and Fasciola hepatica Burdens after Experimental Infection and Vaccination with Glutathione S-Transferase. Vet. Sci. 2022, 9, 58. [Google Scholar] [CrossRef]
- Wijffels, G.L.; Sexton, J.L.; Salvatore, L.; Pettitt, J.M.; Humphris, D.C.; Panaccio, M.; Spithill, T.W. Primary sequence heterogeneity and tissue expression of glutathione S-transferases of Fasciola hepatica. Exp. Parasitol. 1992, 74, 87–99. [Google Scholar] [CrossRef]
- Wilson, R.A.; Wright, J.M.; de Castro-Borges, W.; Parker-Manuel, S.J.; Dowle, A.A.; Ashton, P.D.; Young, N.D.; Gasser, R.B.; Spithill, T.W. Exploring the Fasciola hepatica tegument proteome. Int. J. Parasitol. 2011, 41, 1347–1359. [Google Scholar] [CrossRef] [PubMed]
- Munita, M.P.; Rea, R.; Martinez-Ibeas, A.M.; Byrne, N.; Kennedy, A.; Sekiya, M.; Mulcahy, G.; Sayers, R. Comparison of four commercially available ELISA kits for diagnosis of Fasciola hepatica in Irish cattle. BMC Vet. Res. 2019, 15, 414. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Jayasena, D.D.; Jung, Y.; Jung, S.; Jo, C. Differential Proteome Analysis of Breast and Thigh Muscles between Korean Native Chickens and Commercial Broilers. Asian-Australas. J. Anim. Sci. 2012, 25, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Jefferies, J.R.; Campbell, A.M.; van Rossum, A.J.; Barrett, J.; Brophy, P.M. Proteomic analysis of Fasciola hepatica excretory-secretory products. Proteomics 2001, 1, 1128–1132. [Google Scholar] [CrossRef]
- Pérez-Sánchez, R.; Ramajo-Hernández, A.; Ramajo-Martín, V.; Oleaga, A. Proteomic analysis of the tegument and excretory-secretory products of adult Schistosoma bovis worms. Proteomics 2006, 6 (Suppl. S1), S226–S236. [Google Scholar] [CrossRef]
- LaCourse, E.J.; Perally, S.; Morphew, R.M.; Moxon, J.V.; Prescott, M.; Dowling, D.J.; O’Neill, S.M.; Kipar, A.; Hetzel, U.; Hoey, E.; et al. The Sigma class glutathione transferase from the liver fluke Fasciola hepatica. PLoS Neglected. Trop. Dis. 2012, 6, e1666. [Google Scholar] [CrossRef] [PubMed]
- Brophy, P.M.; Barrett, J. Glutathione transferase in helminths. Parasitology 1990, 100, 345–349. [Google Scholar] [CrossRef]
- Wang, Y.K.; Yan, Y.X.; Ji, W.H.; Wang, H.A.; Zou, Q.; Sun, J.H. Novel Chemiluminescence Immunoassay for the Determination of Zearalenone in Food Samples Using Gold Nanoparticles Labeled with Streptavidin–Horseradish Peroxidase. J. Agric. Food Chem. 2013, 61, 4250–4256. [Google Scholar] [CrossRef]
- Bustos, J.A.; Ninaquispe, B.E.; Rodriguez, S.; Castillo, Y.; Yang, S.Y.; Gilman, R.H.; Dorny, P.; Gabriël, S.; García, H.H.; Gonzalez, A.E.; et al. Performance of a Sandwich Antigen-Detection ELISA for the Diagnosis of Porcine Cysticercosis. Am. J. Trop. Med. Hyg. 2019, 100, 604–608. [Google Scholar] [CrossRef]
- Lan, Z.; Liu, X.L.; Lv, Q.B.; Zeng, M.H.; Gao, J.F.; Chang, Q.C.; Chen, Y.Y.; Wang, C.R. Proteomic Analysis of Fasciola hepatica Excretory and Secretory Products Co-Immunoprecipitated Using Time Course Infection Sera. Pathogens 2021, 10, 749. [Google Scholar] [CrossRef]
- Moazeni, M.; Ahmadi, A. Controversial aspects of the life cycle of Fasciola hepatica. Exp. Parasitol. 2016, 169, 81–89. [Google Scholar] [CrossRef]
- Kueakhai, P.; Chaithirayanon, K.; Chaiwichien, A.; Samrit, T.; Osotprasit, S.; Suksomboon, P.; Jaikua, W.; Sobhon, P.; Changklungmoa, N. Monoclonal antibody against Fasciola gigantica glutathione peroxidase and their immunodiagnosis potential for fasciolosis. Vet. Parasitol. 2019, 276, 108979. [Google Scholar] [CrossRef] [PubMed]
- Valero, M.A.; Ubeira, F.M.; Khoubbane, M.; Artigas, P.; Mas-Coma, S. MM3-ELISA evaluation of coproantigen release and serum antibody production in sheep experimentally infected with Fasciola hepatica and F. gigantica. Vet. Parasitol. 2009, 159, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Alba, A.; Grech-Angelini, S.; Vázquez, A.A.; Alda, P.; Blin, Q.; Lemmonier, L.; Chauvin, A.; Chartier, C.; Douchet, P.; Hurtrez-Boussès, S.; et al. Fasciolosis in the Mediterranean island of Corsica (France): Insights from epidemiological and malacological investigations. Food Waterborne Parasitol. 2023, 19, e00188. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Number | Size/bp |
|---|---|---|
| Glutathione S transferase | 2WRT_A | 657 |
| Vitellin protein B2 | THD21181.1 | 810 |
| Cathepsin L | AAR99519.1 | 717 |
| Flagellar radial spoke protein 3 | TPP67460.1 | 1320 |
| Hypothetical protein D915_006170 | THD22877.1 | 4380 |
| Hypothetical protein D915_007826 | THD21079.1 | 1383 |
| Hypothetical protein D915_007355 | THD21541.1 | 672 |
| Hypothetical protein D915_010250 | THD19012.1 | 447 |
| Hypothetical protein D915_009223 | THD19808.1 | 375 |
| Hypothetical protein D915_005856 | THD23564.1 | 3357 |
| Hypothetical protein D915_003548 | THD25798.1 | 2520 |
| Hypothetical protein D915_007543 | THD21302.1 | 402 |
| Hypothetical protein D915_009822 | THD19402.1 | 300 |
| Hypothetical protein D915_009429 | THD19927.1 | 7968 |
| Hypothetical protein D915_008999 | THD20139.1 | 1884 |
| Hypothetical protein D915_004302 | THD24864.1 | 489 |
| Hypothetical protein D915_006291 | THD22951.1 | 1086 |
| Number | 2D3 | 3D8 | 3E5 | 4E3 | 6F3 |
|---|---|---|---|---|---|
| Affinity Constant | 1.71 × 1010 | 4.52 × 109 | 1.06 × 1010 | 1.48 × 1010 | 2.74 × 109 |
| Sample No. | Intra Batch CV% | Inter Batch CV% | ||
|---|---|---|---|---|
| ± s | CV | CV | ||
| 1 | 0.087 ± 0.004 | 4.14 | 0.089 ± 0.005 | 5.29 |
| 2 | 0.085 ± 0.002 | 2.43 | 0.09 ± 0.004 | 3.93 |
| 3 | 0.091 ± 0.004 | 4.79 | 0.088 ± 0.005 | 5.60 |
| 4 | 0.093 ± 0.002 | 1.86 | 0.090 ± 0.004 | 4.70 |
| 5 | 0.095 ± 0.001 | 1.05 | 0.090 ± 0.005 | 5.03 |
| 6 | 0.105 ± 0.003 | 2.92 | 0.101 ± 0.006 | 5.58 |
| 7 | 0.084 ± 0.004 | 4.76 | 0.088 ± 0.005 | 5.65 |
| 8 | 0.085 ± 0.003 | 3.77 | 0.086 ± 0.002 | 2.48 |
| 9 | 0.257 ± 0.003 | 1.19 | 0.259 ± 0.005 | 1.94 |
| 10 | 0.325 ± 0.004 | 1.28 | 0.331 ± 0.003 | 0.85 |
| 11 | 0.194 ± 0.004 | 1.81 | 0.199 ± 0.002 | 1.07 |
| 12 | 0.273 ± 0.002 | 0.76 | 0.277 ± 0.003 | 1.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, J.; Zhang, N.; Liu, S.; Li, J.; Gong, P.; Wang, X.; Li, X.; Zhang, X.; Tang, B.; Zhang, X. The Detection of Circulating Antigen Glutathione S-Transferase in Sheep Infected with Fasciola hepatica with Double-Antibody Sandwich Signal Amplification Enzyme-Linked Immunosorbent Assay. Animals 2024, 14, 506. https://doi.org/10.3390/ani14030506
Duan J, Zhang N, Liu S, Li J, Gong P, Wang X, Li X, Zhang X, Tang B, Zhang X. The Detection of Circulating Antigen Glutathione S-Transferase in Sheep Infected with Fasciola hepatica with Double-Antibody Sandwich Signal Amplification Enzyme-Linked Immunosorbent Assay. Animals. 2024; 14(3):506. https://doi.org/10.3390/ani14030506
Chicago/Turabian StyleDuan, Jiahui, Nan Zhang, Shaoxiong Liu, Jianhua Li, Pengtao Gong, Xiaocen Wang, Xin Li, Xu Zhang, Bo Tang, and Xichen Zhang. 2024. "The Detection of Circulating Antigen Glutathione S-Transferase in Sheep Infected with Fasciola hepatica with Double-Antibody Sandwich Signal Amplification Enzyme-Linked Immunosorbent Assay" Animals 14, no. 3: 506. https://doi.org/10.3390/ani14030506
APA StyleDuan, J., Zhang, N., Liu, S., Li, J., Gong, P., Wang, X., Li, X., Zhang, X., Tang, B., & Zhang, X. (2024). The Detection of Circulating Antigen Glutathione S-Transferase in Sheep Infected with Fasciola hepatica with Double-Antibody Sandwich Signal Amplification Enzyme-Linked Immunosorbent Assay. Animals, 14(3), 506. https://doi.org/10.3390/ani14030506