
Effects of Individual Essential Amino Acids on Growth Rates of Young Rats Fed a Low-Protein Diet

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Diets
2.2. Sample Collection and Measure
2.3. Statistics and Analysis
3. Results and Discussion
3.1. Determination of Crude Protein Concentration of Low-Protein Diet
(p < 0.001, R2 = 0.73);
3.2. Growth Performance
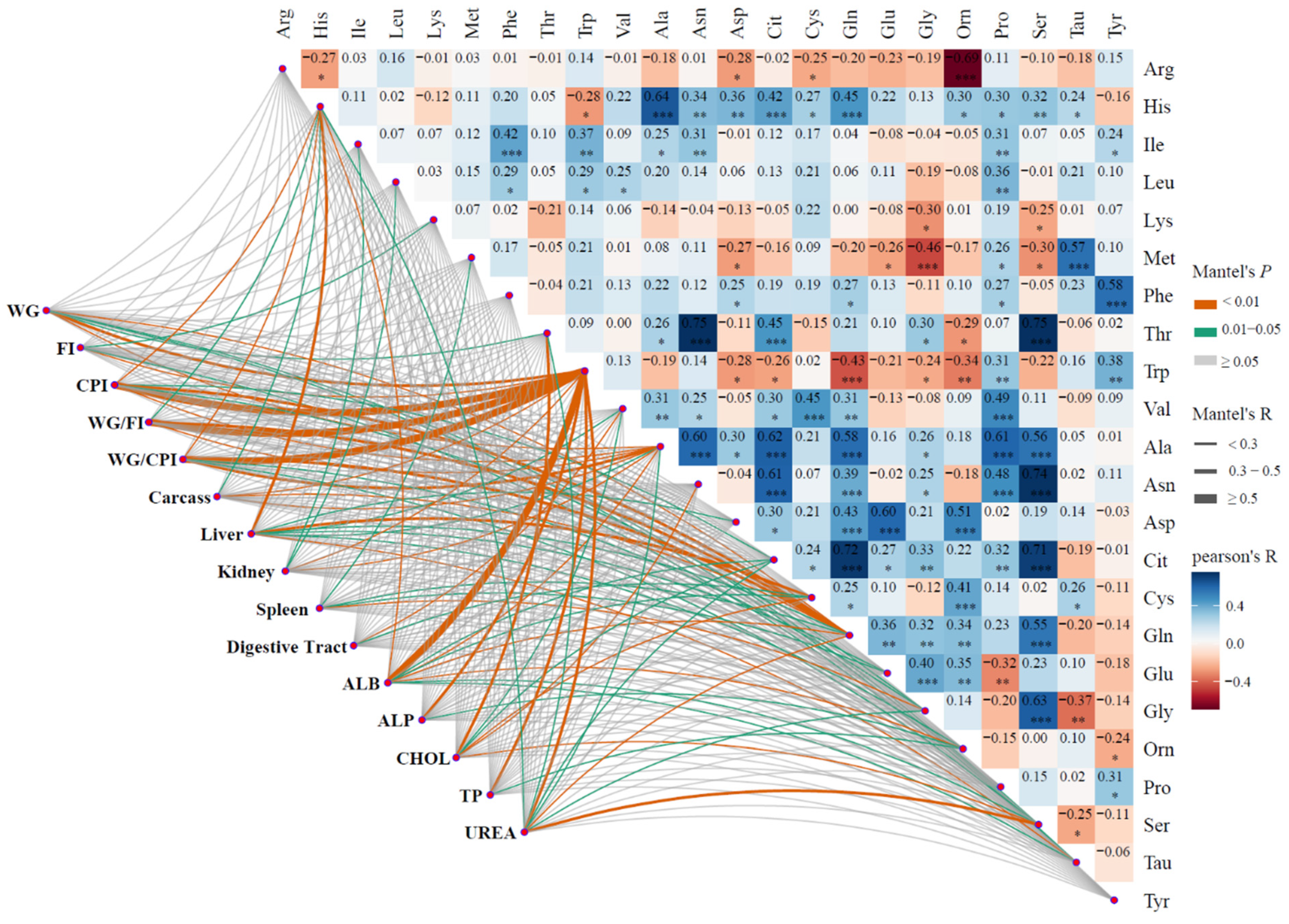
3.3. Plasma Free Amino Acids Concentrations
3.4. Plasma Biochemical Indexes
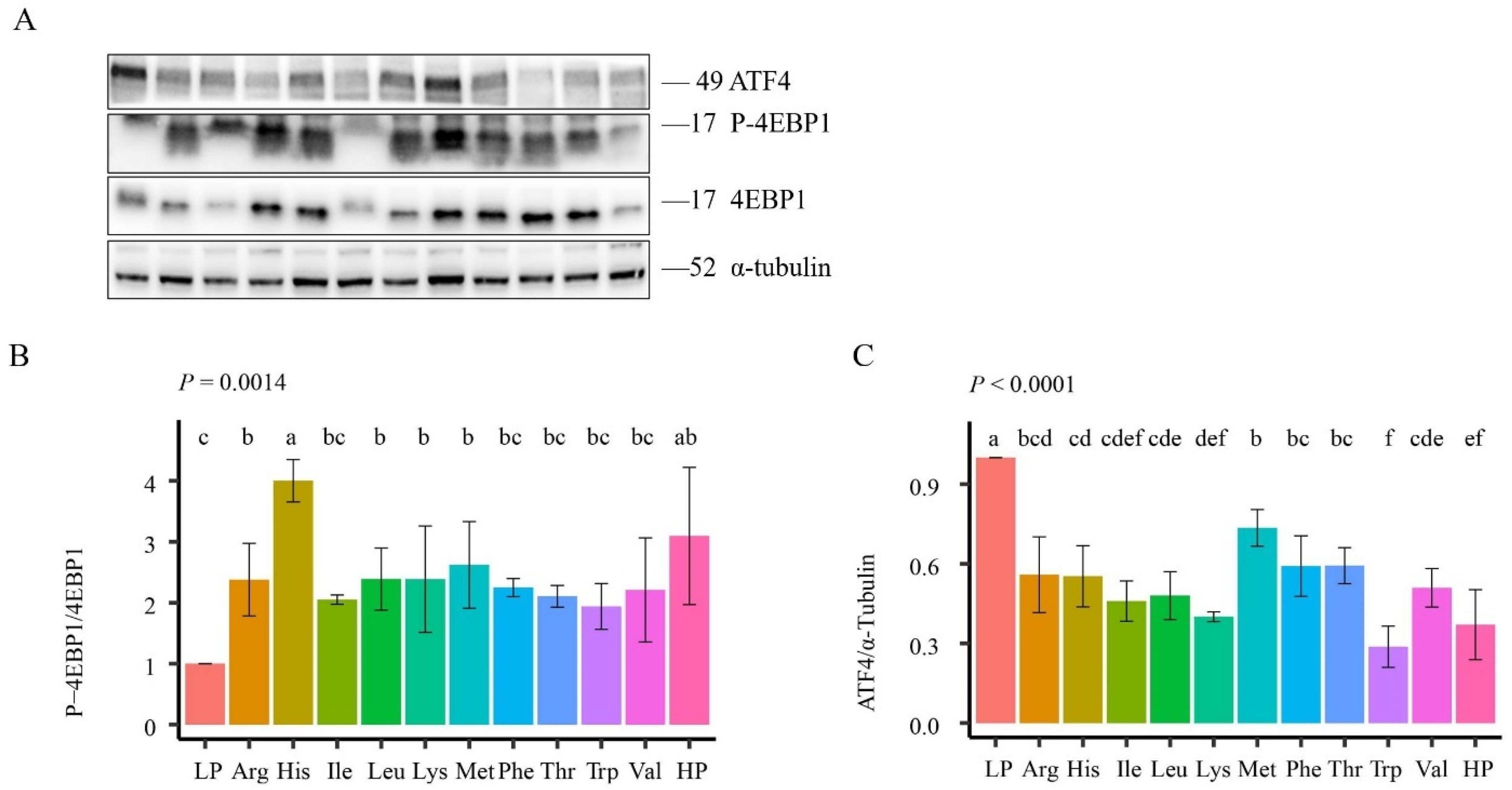
3.5. mTORC1 and ATF4 Signaling Pathways’ Activity in Rat Muscle
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arriola Apelo, S.I.; Knapp, J.R.; Hanigan, M.D. Invited review: Current representation and future trends of predicting amino acid utilization in the lactating dairy cow. J. Dairy Sci. 2014, 97, 4000–4017. [Google Scholar] [CrossRef]
- Wolfe, A.H.; Patz, J.A. Reactive nitrogen and human health: Acute and long-term implications. Ambio 2002, 31, 120–125. [Google Scholar] [CrossRef]
- Mitchell, H.H.; Block, R.J. Some relationships between the amino acid contents of proteins and their nutritive values for the rat. J. Biol. Chem. 1946, 163, 599–620. [Google Scholar] [CrossRef] [PubMed]
- von Liebig, J.F. The Natural Laws of Husbandry; Walton & Maberly: London, UK, 1863. [Google Scholar]
- Yoder, P.S.; Huang, X.; Teixeira, I.A.; Cant, J.P.; Hanigan, M.D. Effects of jugular infused methionine, lysine, and histidine as a group or leucine and isoleucine as a group on production and metabolism in lactating dairy cows. J. Dairy Sci. 2020, 103, 2387–2404. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Liu, W.; Lin, X.Y.; Hu, Z.Y.; Yan, Z.G.; Wang, Y.; Shi, K.R.; Liu, G.M.; Wang, Z.H. Effects of rumen-protected methionine and other essential amino acid supplementation on milk and milk component yields in lactating Holstein cows. J. Dairy Sci. 2019, 102, 7936–7947. [Google Scholar] [CrossRef] [PubMed]
- Arriola Apelo, S.I.; Singer, L.M.; Ray, W.K.; Helm, R.F.; Lin, X.Y.; McGilliard, M.L.; St-Pierre, N.R.; Hanigan, M.D. Casein synthesis is independently and additively related to individual essential amino acid supply. J. Dairy Sci. 2014, 97, 2998–3005. [Google Scholar] [CrossRef] [PubMed]
- Appuhamy, J.A.; Bell, A.L.; Nayananjalie, W.A.; Escobar, J.; Hanigan, M.D. Essential amino acids regulate both initiation and elongation of mRNA translation independent of insulin in MAC-T cells and bovine mammary tissue slices. J. Nutr. 2011, 141, 1209–1215. [Google Scholar] [CrossRef]
- Pszczolkowski, V.L.; Arriola Apelo, S.I. The market for amino acids: Understanding supply and demand of substrate for more efficient milk protein synthesis. J. Anim. Sci. Biotechnol. 2020, 11, 108. [Google Scholar] [CrossRef]
- National Academy of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle: Eighth Revised Edition; The National Academies Press: Washington, DC, USA, 2021. [Google Scholar]
- Shelton, N.W.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; Nelssen, J.L.; DeRouchey, J.M. Effects of increasing dietary standardized ileal digestible lysine for gilts grown in a commercial finishing environment. J. Anim. Sci. 2011, 89, 3587–3595. [Google Scholar] [CrossRef]
- Figueroa, J.L.; Lewis, A.J.; Miller, P.S.; Fischer, R.L.; Diedrichsen, R.M. Growth, carcass traits, and plasma amino acid concentrations of gilts fed low-protein diets supplemented with amino acids including histidine, isoleucine, and valine. J. Anim. Sci. 2003, 81, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Yao, K.; Liu, Z.; Gong, M.; Ruan, Z.; Deng, D.; Tan, B.; Liu, Z.; Wu, G. Supplementing L-leucine to a low-protein diet increases tissue protein synthesis in weanling pigs. Amino Acids 2010, 39, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Kerr, B.J.; Southern, L.L.; Bidner, T.D.; Friesen, K.G.; Easter, R.A. Influence of dietary protein level, amino acid supplementation, and dietary energy levels on growing-finishing pig performance and carcass composition. J. Anim. Sci. 2003, 81, 3075–3087. [Google Scholar] [CrossRef] [PubMed]
- Corzo, A.; Fritts, C.A.; Kidd, M.T.; Kerr, B.J. Response of broiler chicks to essential and non-essential amino acid supplementation of low crude protein diets. Anim. Feed. Sci. Technol. 2005, 118, 319–327. [Google Scholar] [CrossRef]
- Abdel-Maksoud, A.; Yan, F.; Cerrate, S.; Coto, C.; Waldroup, P.W. Effect of Dietary Crude Protein, Lysine Level and Amino Acid Balance on Performance of Broilers 0 to 18 Days of Age. Int. J. Poult. Sci. 2010, 9, 21–27. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in low-protein diets for swine. J. Anim. Sci. Biotechnol. 2018, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Sato, Y.; Obeng, K.A.; Suzuki, D.; Komiya, Y.; Adachi, S.I.; Yoshizawa, F.; Sato, Y. Energy metabolism profile of the effects of amino acid treatment on skeletal muscle cells: Leucine inhibits glycolysis of myotubes. Nutrition 2020, 77, 110794. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Yin, Y.L.; Chu, W.; Liu, Z.; Deng, D.; Li, T.; Huang, R.; Zhang, J.; Tan, B.; Wang, W.; et al. Dietary arginine supplementation increases mTOR signaling activity in skeletal muscle of neonatal pigs. J. Nutr. 2008, 138, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.L.; Ye, J.L.; Yang, J.; Gao, C.Q.; Yan, H.C.; Li, H.C.; Wang, X.Q. mTORC1 Mediates Lysine-Induced Satellite Cell Activation to Promote Skeletal Muscle Growth. Cells 2019, 8, 1549. [Google Scholar] [CrossRef]
- Wen, C.; Chen, X.; Chen, G.Y.; Wu, P.; Chen, Y.P.; Zhou, Y.M.; Wang, T. Methionine improves breast muscle growth and alters myogenic gene expression in broilers. J. Anim. Sci. 2014, 92, 1068–1073. [Google Scholar] [CrossRef]
- Kim, S.W.; Chen, H.; Parnsen, W. Regulatory Role of Amino Acids in Pigs Fed on Protein-restricted Diets. Curr. Protein Pept. Sci. 2019, 20, 132–138. [Google Scholar] [CrossRef]
- Dean, E.D. A Primary Role for α-Cells as Amino Acid Sensors. Diabetes 2020, 69, 542–549. [Google Scholar] [CrossRef]
- Sancak, Y.; Bar-Peled, L.; Zoncu, R.; Markhard, A.L.; Nada, S.; Sabatini, D.M. Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 2010, 141, 290–303. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; Solon-Biet, S.M.; Cogger, V.C.; Ribeiro, R.; de Cabo, R.; Raubenheimer, D.; Cooney, G.J.; Simpson, S.J. Branched chain amino acids, aging and age-related health. Ageing Res. Rev. 2020, 64, 101198. [Google Scholar] [CrossRef] [PubMed]
- Bifari, F.; Nisoli, E. Branched-chain amino acids differently modulate catabolic and anabolic states in mammals: A pharmacological point of view. Br. J. Pharmacol. 2017, 174, 1366–1377. [Google Scholar] [CrossRef]
- Kandasamy, P.; Gyimesi, G.; Kanai, Y.; Hediger, M.A. Amino acid transporters revisited: New views in health and disease. Trends Biochem. Sci. 2018, 43, 752–789. [Google Scholar] [CrossRef]
- Marion, V.; Sankaranarayanan, S.; de Theije, C.; van Dijk, P.; Lindsey, P.; Lamers, M.C.; Harding, H.P.; Ron, D.; Lamers, W.H.; Köhler, S.E. Arginine deficiency causes runting in the suckling period by selectively activating the stress kinase GCN2. J. Biol. Chem. 2011, 286, 8866–8874. [Google Scholar] [CrossRef]
- Marques-Ramos, A.; Candeias, M.M.; Menezes, J.; Lacerda, R.; Willcocks, M.; Teixeira, A.; Locker, N.; Romão, L. Cap-independent translation ensures mTOR expression and function upon protein synthesis inhibition. Rna 2017, 23, 1712–1728. [Google Scholar] [CrossRef]
- Klann, K.; Tascher, G.; Münch, C. Functional Translatome Proteomics Reveal Converging and Dose-Dependent Regulation by mTORC1 and eIF2α. Mol. Cell 2020, 77, 913–925.e914. [Google Scholar] [CrossRef] [PubMed]
- Castilho, B.A.; Shanmugam, R.; Silva, R.C.; Ramesh, R.; Himme, B.M.; Sattlegger, E. Keeping the eIF2 alpha kinase Gcn2 in check. Biochim. Biophys. Acta 2014, 1843, 1948–1968. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Reyna-Neyra, A.; Philippe, L.; Thoreen, C.C. mTORC1 Balances Cellular Amino Acid Supply with Demand for Protein Synthesis through Post-transcriptional Control of ATF4. Cell Rep. 2017, 19, 1083–1090. [Google Scholar] [CrossRef]
- Bröer, S.; Bröer, A. Amino acid homeostasis and signalling in mammalian cells and organisms. Biochem. J. 2017, 474, 1935–1963. [Google Scholar] [CrossRef]
- Jin, H.O.; Hong, S.E.; Kim, J.Y.; Jang, S.K.; Park, I.C. Amino acid deprivation induces AKT activation by inducing GCN2/ATF4/REDD1 axis. Cell Death Dis. 2021, 12, 1127. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Nelson, G.M.; Watson, J.L.; Morf, J.; Dalglish, M.; Luh, L.M.; Weber, A.; Bertolotti, A. Protein Stability Buffers the Cost of Translation Attenuation following eIF2α Phosphorylation. Cell Rep. 2020, 32, 108154. [Google Scholar] [CrossRef]
- Ojha, R.; Leli, N.M.; Onorati, A.; Piao, S.; Verginadis, I.I.; Tameire, F.; Rebecca, V.W.; Chude, C.I.; Murugan, S.; Fennelly, C.; et al. ER Translocation of the MAPK Pathway Drives Therapy Resistance in BRAF-Mutant Melanoma. Cancer Discov. 2019, 9, 396–415. [Google Scholar] [CrossRef]
- Fougeray, S.; Bouvier, N.; Beaune, P.; Legendre, C.; Anglicheau, D.; Thervet, E.; Pallet, N. Metabolic stress promotes renal tubular inflammation by triggering the unfolded protein response. Cell Death Dis. 2011, 2, e143. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Koumenis, C. ATF4, an ER stress and hypoxia-inducible transcription factor and its potential role in hypoxia tolerance and tumorigenesis. Curr. Mol. Med. 2009, 9, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Yang, Q.; Jeong, M.H.; Curukovic, A.; Tiwary, S.; Melick, C.H.; Lama-Sherpa, T.D.; Wang, H.; Huerta-Rosario, M.; Urquhart, G.; et al. SNAT7 regulates mTORC1 via macropinocytosis. Proc. Natl. Acad. Sci. USA 2022, 119, e2123261119. [Google Scholar] [CrossRef]
- Meng, D.; Yang, Q.; Wang, H.; Melick, C.H.; Navlani, R.; Frank, A.R.; Jewell, J.L. Glutamine and asparagine activate mTORC1 independently of Rag GTPases. J. Biol. Chem. 2020, 295, 2890–2899. [Google Scholar] [CrossRef]
- Luo, C.; Zhao, S.; Zhang, M.; Gao, Y.; Wang, J.; Hanigan, M.D.; Zheng, N. SESN2 negatively regulates cell proliferation and casein synthesis by inhibition the amino acid-mediated mTORC1 pathway in cow mammary epithelial cells. Sci. Rep. 2018, 8, 3912. [Google Scholar] [CrossRef]
- Hara, K.; Yonezawa, K.; Weng, Q.P.; Kozlowski, M.T.; Belham, C.; Avruch, J. Amino acid sufficiency and mTOR regulate p70 S6 kinase and eIF-4E BP1 through a common effector mechanism. J. Biol. Chem. 1998, 273, 14484–14494. [Google Scholar] [CrossRef]
- Nicklin, P.; Bergman, P.; Zhang, B.; Triantafellow, E.; Wang, H.; Nyfeler, B.; Yang, H.; Hild, M.; Kung, C.; Wilson, C.; et al. Bidirectional transport of amino acids regulates mTOR and autophagy. Cell 2009, 136, 521–534. [Google Scholar] [CrossRef]
- Battaglioni, S.; Benjamin, D.; Wälchli, M.; Maier, T.; Hall, M.N. mTOR substrate phosphorylation in growth control. Cell 2022, 185, 1814–1836. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Edick, A.M.; Audette, J.; Burgos, S.A. CRISPR-Cas9-mediated knockout of GCN2 reveals a critical role in sensing amino acid deprivation in bovine mammary epithelial cells. J. Dairy Sci. 2021, 104, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Mazor, K.M.; Stipanuk, M.H. GCN2- and eIF2α-phosphorylation-independent, but ATF4-dependent, induction of CARE-containing genes in methionine-deficient cells. Amino Acids 2016, 48, 2831–2842. [Google Scholar] [CrossRef] [PubMed]
- Gray, N.; Zia, R.; King, A.; Patel, V.C.; Wendon, J.; McPhail, M.J.; Coen, M.; Plumb, R.S.; Wilson, I.D.; Nicholson, J.K. High-Speed Quantitative UPLC-MS Analysis of Multiple Amines in Human Plasma and Serum via Precolumn Derivatization with 6-Aminoquinolyl-N-hydroxysuccinimidyl Carbamate: Application to Acetaminophen-Induced Liver Failure. Anal. Chem. 2017, 89, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.; Mayntz, D.; Raubenheimer, D.; Simpson, S.J. Protein-leverage in mice: The geometry of macronutrient balancing and consequences for fat deposition. Obesity 2008, 16, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. Obesity: The protein leverage hypothesis. Obes. Rev. 2005, 6, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Gietzen, D.W.; Hao, S.; Anthony, T.G. Mechanisms of food intake repression in indispensable amino acid deficiency. Annu. Rev. Nutr. 2007, 27, 63–78. [Google Scholar] [CrossRef]
- Leung, P.M.; Rogers, Q.R. Food intake: Regulation by plasma amino acid pattern. Life Sci. 1969, 8, 1–9. [Google Scholar] [CrossRef]
- Yap, Y.W.; Rusu, P.M.; Chan, A.Y.; Fam, B.C.; Jungmann, A.; Solon-Biet, S.M.; Barlow, C.K.; Creek, D.J.; Huang, C.; Schittenhelm, R.B.; et al. Restriction of essential amino acids dictates the systemic metabolic response to dietary protein dilution. Nat. Commun. 2020, 11, 2894. [Google Scholar] [CrossRef]
- Yao, B.; Xu, J.; Qi, Z.; Harris, R.C.; Zhang, M.Z. Role of renal cortical cyclooxygenase-2 expression in hyperfiltration in rats with high-protein intake. Am. J. Physiol. Renal Physiol. 2006, 291, F368–F374. [Google Scholar] [CrossRef]
- Cangelosi, A.L.; Puszynska, A.M.; Roberts, J.M.; Armani, A.; Nguyen, T.P.; Spinelli, J.B.; Kunchok, T.; Wang, B.; Chan, S.H.; Lewis, C.A.; et al. Zonated leucine sensing by Sestrin-mTORC1 in the liver controls the response to dietary leucine. Science 2022, 377, 47–56. [Google Scholar] [CrossRef]
- Fuller, M.F.; Livingstone, R.M.; Baird, B.A.; Atkinson, T. The optimal amino acid supplementation of barley for the growing pig. 1. Response of nitrogen metabolism to progressive supplementation. Br. J. Nutr. 1979, 41, 321–331. [Google Scholar] [CrossRef]
- Wiedmer, P.; Jung, T.; Castro, J.P.; Pomatto, L.C.D.; Sun, P.Y.; Davies, K.J.A.; Grune, T. Sarcopenia-Molecular mechanisms and open questions. Ageing Res. Rev. 2021, 65, 101200. [Google Scholar] [CrossRef] [PubMed]
- Holeček, M. Influence of Histidine Administration on Ammonia and Amino Acid Metabolism: A Review. Physiol. Res. 2020, 69, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Waterland, R.A. Assessing the effects of high methionine intake on DNA methylation. J. Nutr. 2006, 136, 1706s–1710s. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Chakraborty, A.; Huang, S.C.; Wang, P.Y.; Hsieh, Y.J.; Chien, K.Y.; Lee, Y.H.; Chang, C.C.; Tang, H.Y.; Lin, Y.T.; et al. Histidine-Dependent Protein Methylation Is Required for Compartmentalization of CTP Synthase. Cell Rep. 2018, 24, 2733–2745.e2739. [Google Scholar] [CrossRef]
- Sarwar, G.; Peace, R.W.; Botting, H.G. Influence of high dietary threonine on growth and amino acids in blood and tissues of rats. Amino Acids 1995, 8, 69–78. [Google Scholar] [CrossRef]
- Rosell, V.L.; Zimmerman, D.R. Threonine requirement of pigs weighing 5 to 15 kg and the effect of excess methionine in diets marginal in threonine. J. Anim. Sci. 1985, 60, 480–486. [Google Scholar] [CrossRef]
- Tom, A.; Nair, K.S. Assessment of branched-chain amino Acid status and potential for biomarkers. J. Nutr. 2006, 136, 324s–330s. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Lamers, W.H.; Broer, S.; van de Graaf, S.F.J. Amino acid metabolism, transport and signalling in the liver revisited. Biochem. Pharmacol. 2022, 201, 115074. [Google Scholar] [CrossRef]
- Soukoulis, V.; Dihu, J.B.; Sole, M.; Anker, S.D.; Cleland, J.; Fonarow, G.C.; Metra, M.; Pasini, E.; Strzelczyk, T.; Taegtmeyer, H.; et al. Micronutrient deficiencies an unmet need in heart failure. J. Am. Coll. Cardiol. 2009, 54, 1660–1673. [Google Scholar] [CrossRef]
- Yang, H.B.; Xu, Y.Y.; Zhao, X.N.; Zou, S.W.; Zhang, Y.; Zhang, M.; Li, J.T.; Ren, F.; Wang, L.Y.; Lei, Q.Y. Acetylation of MAT IIα represses tumour cell growth and is decreased in human hepatocellular cancer. Nat. Commun. 2015, 6, 6973. [Google Scholar] [CrossRef]
- Parkhitko, A.A.; Filine, E.; Mohr, S.E.; Moskalev, A.; Perrimon, N. Targeting metabolic pathways for extension of lifespan and healthspan across multiple species. Ageing Res. Rev. 2020, 64, 101188. [Google Scholar] [CrossRef]
- Siegert, W.; Rodehutscord, M. The relevance of glycine and serine in poultry nutrition: A review. Br. Poult. Sci. 2019, 60, 579–588. [Google Scholar] [CrossRef]
- Kim, B.; Cho, E.J.; Yoon, J.H.; Kim, S.S.; Cheong, J.Y.; Cho, S.W.; Park, T. Pathway-Based Integrative Analysis of Metabolome and Microbiome Data from Hepatocellular Carcinoma and Liver Cirrhosis Patients. Cancers 2020, 12, 2705. [Google Scholar] [CrossRef] [PubMed]
- Keshet, R.; Szlosarek, P.; Carracedo, A.; Erez, A. Rewiring urea cycle metabolism in cancer to support anabolism. Nat. Rev. Cancer 2018, 18, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M., Jr. Regulation of enzymes of the urea cycle and arginine metabolism. Annu. Rev. Nutr. 2002, 22, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, R.; Caraceni, P.; Bernardi, M.; Gines, P.; Arroyo, V.; Jalan, R. Albumin: Pathophysiologic basis of its role in the treatment of cirrhosis and its complications. Hepatology 2013, 58, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Jagdish, R.K.; Maras, J.S.; Sarin, S.K. Albumin in Advanced Liver Diseases: The Good and Bad of a Drug! Hepatology 2021, 74, 2848–2862. [Google Scholar] [CrossRef]
- Hou, Y.; Hu, S.; Li, X.; He, W.; Wu, G. Amino Acid Metabolism in the Liver: Nutritional and Physiological Significance. Adv. Exp. Med. Biol. 2020, 1265, 21–37. [Google Scholar] [CrossRef]
- Tsien, C.; Davuluri, G.; Singh, D.; Allawy, A.; Ten Have, G.A.; Thapaliya, S.; Schulze, J.M.; Barnes, D.; McCullough, A.J.; Engelen, M.P.; et al. Metabolic and molecular responses to leucine-enriched branched chain amino acid supplementation in the skeletal muscle of alcoholic cirrhosis. Hepatology 2015, 61, 2018–2029. [Google Scholar] [CrossRef] [PubMed]
- Adegoke, O.A.; Abdullahi, A.; Tavajohi-Fini, P. mTORC1 and the regulation of skeletal muscle anabolism and mass. Appl. Physiol. Nutr. Metab. 2012, 37, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.M.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G.M. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhong, J.; Vehkamäki, H.; Kurtén, T.; Wang, W.; Ge, M.; Zhang, S.; Li, Z.; Zhang, X.; Francisco, J.S.; et al. Self-Catalytic Reaction of SO3 and NH3 To Produce Sulfamic Acid and Its Implication to Atmospheric Particle Formation. J. Am. Chem. Soc. 2018, 140, 11020–11028. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredients (g/kg) | Dietary Protein Concentration | ||||||
|---|---|---|---|---|---|---|---|
| 2% | 6% | 10% | 14% | 18% | 22% | 26% | |
| Casein 1 | 22 | 67 | 112 | 156 | 200 | 246 | 291 |
| L-Methionine 2 | 0.30 | 1.00 | 1.65 | 2.34 | 3.00 | 3.70 | 4.38 |
| Corn starch | 578 | 532 | 487 | 442 | 397 | 351 | 305 |
| Maltodextrin | 132 | 132 | 132 | 132 | 132 | 132 | 132 |
| Sucrose | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Cellulose | 50 | 50 | 50 | 50 | 50 | 50 | 50 |
| Soybean oil | 70 | 70 | 70 | 70 | 70 | 70 | 70 |
| Mineral mix | 35 | 35 | 35 | 35 | 35 | 35 | 35 |
| Vitamin mix | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Choline | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| Butyl hydroquinone | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 |
| Nutrient level (%) | |||||||
| Protein | 1.97 | 6.00 | 9.97 | 13.96 | 17.90 | 22.02 | 26.05 |
| Carbohydrate | 82.01 | 77.44 | 82.01 | 68.41 | 63.95 | 59.27 | 54.71 |
| Fat | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 |
| Total energy (Mcal/kg) | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 |
| Ingredients (g/kg) | LP | HP | LP + Val | LP + Ile | LP + Trp | LP + Thr | LP + Lys | LP + Phe | LP + Leu | LP + arg | LP + His | LP + Met |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Casein 1 | 67 | 200 | 67 | 67 | 67 | 67 | 67 | 67 | 67 | 67 | 67 | 67 |
| L-Valine 2 | 4.0 | 11.8 | 11.8 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 |
| L-Isoleucine | 3.1 | 9.3 | 3.1 | 9.3 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 |
| L-Tryptophan | 0.8 | 2.3 | 0.8 | 0.8 | 2.3 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| L-Threonine | 2.5 | 7.6 | 2.5 | 2.5 | 2.5 | 7.6 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| L-Lysine | 4.7 | 14.1 | 4.7 | 4.7 | 4.7 | 4.7 | 14.1 | 4.7 | 4.7 | 4.7 | 4.7 | 4.7 |
| L-Phenylalanine | 3.1 | 9.2 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 | 9.2 | 3.1 | 3.1 | 3.1 | 3.1 |
| L-Leucine | 5.5 | 16.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 5.5 | 16.5 | 5.5 | 5.5 | 5.5 |
| L-Arginine | 2.2 | 6.7 | 2.2 | 2.2 | 2.2 | 2.2 | 2.2 | 2.2 | 2.2 | 6.7 | 6.7 | 2.2 |
| L-Histidine | 1.7 | 4.9 | 1.7 | 1.7 | 1.7 | 1.7 | 1.7 | 1.7 | 1.7 | 1.7 | 4.9 | 1.7 |
| L-Methionine | 1.8 | 5.3 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 | 5.3 |
| L-Cystine | 1.0 | 3.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Corn starch | 533 | 398 | 525 | 526 | 531 | 527 | 523 | 526 | 521 | 538 | 529 | 529 |
| Maltodextrin | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 |
| Sucrose | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Cellulose | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 |
| Soybean oil | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 | 70 |
| Mineral mix | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 |
| Vitamin mix | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Choline | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| Butyl hydroquinone | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 |
| Nutrients (% of DM) | ||||||||||||
| Protein | 6.0 | 17.9 | 6.8 | 6.6 | 6.2 | 6.5 | 6.9 | 6.6 | 7.1 | 6.4 | 6.3 | 6.4 |
| Carbohydrate | 77.5 | 64.0 | 76.7 | 76.8 | 77.3 | 76.9 | 76.5 | 76.8 | 76.3 | 77 | 77.1 | 77.1 |
| Fat | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 |
| Total energy (Mcal/kg) | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 |
| Item | Protein Concentration | |||||
|---|---|---|---|---|---|---|
| 6% | 10% | 14% | 18% | 22% | 26% | |
| WG, g | 52 ± 10 1 | 65 ± 9 | 60 ± 17 | 62 ± 9 | 70 ± 12 | 71 ± 11 |
| Intake, g | 236 ± 18 | 220 ± 22 | 192 ± 31 | 185 ± 11 | 197 ± 27 | 197 ± 12 |
| CPI, g | 14 ± 1 | 22 ± 2 | 27 ± 4 | 33 ± 2 | 43 ± 6 | 51 ± 3 |
| MEIDMI, Kcal | 942 | 879 | 766 | 739 | 786 | 788 |
| MEICP, Kcal | 81 | 125 | 153 | 189 | 246 | 292 |
| MEINCP, Kcal | 862 | 754 | 613 | 549 | 540 | 496 |
| CPI, g | MEINCP, Kcal | WG, g | δ, g | CP |
|---|---|---|---|---|
| 11.7 | 905 | 52.2 | 10.8 | 5% |
| 13.5 | 867 | 53.5 | 9.5 | 6% |
| 15.3 | 831 | 54.8 | 8.2 | 7% |
| 17.2 | 797 | 56.1 | 6.9 | 8% |
| 19.0 | 765 | 57.3 | 5.7 | 9% |
| 20.8 | 734 | 58.4 | 4.6 | 10% |
| Item | LP | LP + Arg | LP + His | LP + Ile | LP + Leu | LP + Lys | LP + Met | LP + Phe | LP + Thr | LP + Trp | LP + Val | HP | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WG, g | 50.3 e | 53.0 de | 62.0 bc | 58.7 bcd | 55.5 cde | 61.9 bc | 53.0 de | 55.3 cde | 66.3 b | 59.1 bcd | 52.9 de | 79.8 a | 2.3 | <0.01 |
| FI, g | 224 def | 234 bcde | 250 ab | 239 abcde | 236 abcd | 259 a | 220 ef | 232 bcdef | 246 abc | 237 bcde | 228 cdef | 213 f | 5.8 | <0.01 |
| CPI, g | 13.4 g | 15.1 def | 15.8 de | 15.9 cde | 16.7 bc | 17.9 b | 14.0 fg | 15.3 de | 16.0 cd | 14.6 efg | 15.4 de | 38.0 a | 0.4 | <0.01 |
| WG/FI, g/g | 0.22 d | 0.23 d | 0.25 bc | 0.25 bc | 0.23 cd | 0.24 cd | 0.24 cd | 0.24 cd | 0.27 b | 0.25 bc | 0.23 cd | 0.38 a | 0.007 | <0.01 |
| WG/CPI, g/g | 3.8 cd | 3.5 def | 3.9 abc | 3.8 bcd | 3.2 f | 3.5 ef | 3.8 bcd | 3.6 de | 4.1 a | 4.1 ab | 3.4 ef | 2.1 g | 0.095 | <0.01 |
| Carcass, g | 54.9 c | 58.2 bc | 58.1 bc | 57.4 bc | 54.2 c | 63.1 ab | 56.5 c | 54.8 c | 59.7 bc | 57.3 bc | 53.7 c | 66.7 a | 1.9 | <0.01 |
| Liver 1, % | 5.6 abc | 4.9 bc | 6.0 ab | 5.3 bc | 5.6 abc | 5.1 bc | 6.6 a | 5.4 bc | 4.7 c | 5.2 bc | 5.9 ab | 4.6 c | 0.003 | <0.01 |
| Kidney 1, % | 0.87 ab | 0.78 b | 0.88 ab | 0.75 b | 0.90 ab | 0.77 b | 0.79 b | 0.78 b | 0.84 b | 0.82 b | 0.83 b | 1.01 a | 0.0004 | <0.01 |
| Spleen, % | 0.22 a | 0.24 a | 0.24 a | 0.23 a | 0.22 a | 0.21 a | 0.26 a | 0.21 a | 0.22 a | 0.2 a | 0.22 a | 0.27 a | 0.0002 | 0.05 |
| Heart 1, % | 0.54 | 0.54 | 0.57 | 0.54 | 0.54 | 0.55 | 0.52 | 0.57 | 0.56 | 0.57 | 0.51 | 0.49 | 0.0003 | 0.68 |
| Limbs 1, % | 6.9 | 7.1 | 6.5 | 6.3 | 6.3 | 6.5 | 6.3 | 6.5 | 7.4 | 6.6 | 6.9 | 7.3 | 0.004 | 0.74 |
| Digestive Tract 1, % | 4.9 ab | 4.7 ab | 5.0 a | 4.9 ab | 5.2 a | 4.7 ab | 4.2 b | 4.7 ab | 4.8 ab | 4.8 ab | 4.8 ab | 4.6 ab | 0.002 | 0.04 |
| Dependent Variable | Independent Variable | Estimate | SE | Variable p-Value | R2 | Model p-Value |
|---|---|---|---|---|---|---|
| Weight Gain, g | Intercept | 63.31 | 6.32 | <0.001 | 0.59 | <0.001 |
| Glu | 0.03 | 0.01 | 0.006 | |||
| Ile | 0.15 | 0.06 | 0.020 | |||
| Lys | 0.01 | 0.002 | 0.028 | |||
| Ser | −0.08 | 0.02 | <0.001 | |||
| Thr | 0.01 | 0.001 | <0.001 | |||
| Trp | 0.13 | 0.05 | 0.009 | |||
| Feed Intake, g | Intercept | 245.90 | 9.87 | <0.001 | 0.34 | <0.001 |
| Asp | −0.57 | 0.26 | 0.032 | |||
| Glu | 0.08 | 0.04 | 0.026 | |||
| Lys | 0.02 | 0.01 | 0.006 | |||
| Trp | −0.39 | 0.11 | 0.000 | |||
| Tau | −0.32 | 0.11 | 0.005 | |||
| Weight Gain/Feed Intake, | Intercept | 0.27 | 0.02 | <0.001 | 0.75 | <0.001 |
| Glu | 0.0001 | 0.0000 | 0.002 | |||
| Ile | 0.0005 | 0.0002 | 0.022 | |||
| Ser | −0.0004 | 0.0001 | <0.001 | |||
| Thr | 0.00003 | 0.000005 | <0.001 | |||
| Trp | 0.001 | 0.0002 | <0.001 | |||
| Weight Gain /Protein Intake, % | Intercept | 3.43 | 0.30 | <0.001 | 0.75 | <0.001 |
| Ala | 0.002 | 0.001 | <0.001 | |||
| Asn | −0.016 | 0.01 | 0.006 | |||
| His | 0.01 | 0.002 | 0.010 | |||
| Leu | −0.007 | 0.001 | <0.001 | |||
| Thr | 0.0002 | 0.0001 | 0.003 | |||
| Trp | −0.0098 | 0.002 | <0.001 | |||
| Gly | 0.001 | 0.001 | 0.031 | |||
| Liver 1 | Intercept | 0.13 | 0.02 | <0.001 | 0.43 | <0.001 |
| Arg | −0.00024 | 0.0001 | 0.018 | |||
| His | 0.001 | 0.0001 | <0.001 | |||
| Thr | −0.000006 | 0.000003 | 0.025 | |||
| Gly | −0.00008 | 0.00004 | 0.042 | |||
| Albumen | Intercept | 14.76 | 2.02 | <0.001 | 0.7 | <0.001 |
| Arg | 0.04 | 0.01 | 0.002 | |||
| Cys | −0.15 | 0.05 | 0.002 | |||
| Thr | 0.001 | 0.0003 | <0.001 | |||
| Trp | 0.11 | 0.01 | <0.001 | |||
| Cit | −0.08 | 0.02 | 0.001 | |||
| Orn | 0.02 | 0.01 | 0.002 |
| EAA, μmol/L | LP | LP + Arg | LP + His | LP + Ile | LP + Leu | LP + Lys | LP + Met | LP + Phe | LP + Thr | LP + Trp | LP + Val | HP | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arg | 67.42 | 112 | 84 | 85 | 93 | 91 | 69 | 79 | 83 | 83 | 85 | 105 | 10 | 0.11 |
| His | 97.51 cde | 80 efg | 137 a | 107 bcd | 74 fg | 76 fg | 119 ab | 90 def | 104 bcd | 91 def | 113 bc | 63 g | 5.9 | <0.01 |
| Ile | 23.1 de | 23.5 cde | 27.8 cde | 63.0 a | 18.3 e | 27.3 cde | 28.7 cd | 26.5 cde | 32.2 c | 25.8 cde | 29.5 cd | 41.6 b | 2.7 | <0.01 |
| Leu | 125 d | 120 d | 138 bcd | 120 d | 180 a | 128 cd | 136 bcd | 124 cd | 136 bcd | 125 cd | 156 abc | 165 ab | 8.9 | <0.01 |
| Lys | 726 d | 634 d | 737 cd | 894 bcd | 752 bcd | 1667 a | 849 bcd | 775 bcd | 634 d | 748 bcd | 901 bc | 994 b | 71 | <0.01 |
| Met | 22.1 b | 22.4 b | 20.1 b | 25.7 b | 21.1 b | 27.3 b | 75.5 a | 22.8 b | 23.9 b | 20.5 b | 26.5 b | 43.1 b | 6.8 | <0.01 |
| Phe | 68.4 | 73.9 | 76.3 | 82.9 | 71.0 | 72.0 | 80.0 | 84.4 | 73.2 | 73.4 | 79.5 | 83.8 | 4.8 | 0.28 |
| Thr | 59 c | 53 c | 61 c | 62 c | 50 c | 52 c | 55 c | 55 c | 3310 a | 53 c | 58 c | 486 b | 40 | <0.01 |
| Trp | 36.7 bc | 25.4 c | 32.6 bc | 38.8 bc | 27.6 c | 34.0 bc | 34.6 bc | 31.8 bc | 37.6 bc | 43.9 b | 39.0 bc | 87.0 a | 4.0 | <0.01 |
| Val | 24 d | 20 de | 26 d | 28 cd | 14 e | 26 d | 27 d | 26 d | 36 bc | 25 d | 145 a | 38 b | 2.5 | <0.01 |
| NEAA, μmol/L | LP | LP + Arg | LP + His | LP + Ile | LP + Leu | LP + Lys | LP + Met | LP + Phe | LP + Thr | LP + Trp | LP + Val | HP | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ala | 547 abc | 474 cd | 550 abc | 615 ab | 507 bcd | 473 cd | 593 abc | 517 abc | 648 a | 540 abc | 637 ab | 373 d | 37 | <0.01 |
| Asp | 31 | 30 | 33 | 29 | 28 | 21 | 24 | 28 | 24 | 23 | 26 | 20 | 3.5 | 0.19 |
| Asn | 43.9 bcd | 37.6 cd | 40.3 bcd | 48.1 bc | 35.0 d | 43.2 bcd | 46.9 bcd | 41.8 bcd | 74.4 a | 40.2 bcd | 51.1 bcd | 43.7 b | 3.2 | <0.01 |
| Cys | 9.1 b | 7.4 b | 9.9 b | 12.4 ab | 8.0 b | 10.2 b | 12.6 ab | 7.2 b | 7.4 b | 7.4 b | 17.0 a | 10.4 b | 1.6 | <0.01 |
| Gln | 605 abc | 585 bc | 616 abc | 637 abc | 592 abc | 632 abc | 560 c | 580 bc | 696 ab | 599 abc | 714 a | 422 d | 34 | <0.01 |
| Glu | 187 | 188 | 244 | 149 | 174 | 156 | 144 | 185 | 200 | 212 | 152 | 160 | 25 | 0.16 |
| Gly | 321 ab | 239 cd | 234 cd | 273 bc | 240 cd | 195 de | 174 e | 270 c | 322 ab | 330 a | 234 cd | 171 e | 15 | <0.01 |
| Pro | 23.3 ab | 18.7 b | 22.6 ab | 28.1 ab | 22.7 ab | 23.8 ab | 25.3 ab | 21.0 b | 25.1 ab | 19.9 b | 32.6 a | 24.7 ab | 2.4 | 0.01 |
| Ser | 371 b | 313 cde | 313 cde | 350 bcd | 321 bcde | 299 de | 286 e | 322 bcde | 549 a | 348 bcd | 362 bc | 233 f | 16 | <0.01 |
| Tyr | 11.4 c | 12.3 c | 11.8 c | 16.7 bc | 11.5 c | 15.3 c | 12.7 c | 29.0 a | 15.1 c | 13.1 c | 15.9 bc | 21.8 b | 1.8 | <0.01 |
| Tau | 25.1 c | 21.5 c | 27.4 c | 18.8 c | 20.5 c | 17.5 c | 72.0 a | 20.8 c | 22.0 c | 20.3 c | 20.2 c | 42.1 b | 3.2 | <0.01 |
| Cit | 76.1 bc | 76.3 bc | 81.1 bc | 80.9 bc | 74.1 cd | 79.3 bc | 72.7 cd | 77.0 bc | 96.6 a | 74.3 cd | 89.2 ab | 62.6 d | 3.8 | <0.01 |
| Orn | 147 | 154 | 135 | 127 | 116 | 110 | 130 | 126 | 82 | 107 | 145 | 72 | 18 | 0.06 |
| Item | LP | LP + Arg | LP + His | LP + Ile | LP + Leu | LP + Lys | LP + Met | LP + Phe | LP + Thr | LP + Trp | LP + Val | HP | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ALB, g/L | 18 cd | 19 bc | 17 cde | 17 cde | 16 cde | 18 de | 16 e | 18 bcd | 20 b | 18 bcd | 16 de | 25 a | 0.66 | <0.01 |
| ALP, U/L | 641 ab | 805 a | 717 a | 720 a | 708 ab | 778 a | 640 ab | 755 a | 619 ab | 738 a | 680 ab | 467 b | 59 | 0.01 |
| ALT, U/L | 41 | 44 | 41 | 33 | 39 | 45 | 32 | 41 | 40 | 40 | 30 | 33 | 4.7 | 0.33 |
| AST, U/L | 177 | 144 | 177 | 135 | 132 | 164 | 170 | 141 | 157 | 143 | 154 | 115 | 15 | 0.10 |
| Glucose, mmol/L | 9.3 | 9.2 | 9.4 | 9.0 | 8.9 | 9.2 | 8.7 | 9.2 | 9.5 | 9.6 | 9.2 | 9.5 | 0.44 | 0.97 |
| CHOL, mmol/L | 1.5 bc | 1.5 bc | 1.7 b | 1.5 bc | 1.5 bc | 1.9 b | 1.6 bc | 1.7 bc | 2.4 a | 1.8 b | 1.2 c | 1.9 b | 0.13 | <0.01 |
| TG, mmol/L | 0.75 | 0.64 | 0.83 | 0.67 | 0.93 | 0.80 | 0.66 | 0.76 | 0.68 | 0.92 | 0.67 | 0.65 | 0.10 | 0.45 |
| TP, g/L | 53 b | 55 b | 52 b | 52 b | 48 b | 54 b | 50 b | 52 b | 55 b | 49 b | 49 b | 65 a | 2.0 | <0.01 |
| Urea, mmol/L | 2.2 cd | 3.4 bc | 3.7 b | 3.4 bc | 2.7 bcd | 3.0 bcd | 2.3 cd | 2.4 cd | 1.7 d | 2.4 cd | 3.5 bc | 6.8 a | 0.37 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Wang, T.; Zhao, K.; Hanigan, M.D.; Lin, X.; Hu, Z.; Hou, Q.; Wang, Y.; Wang, Z. Effects of Individual Essential Amino Acids on Growth Rates of Young Rats Fed a Low-Protein Diet. Animals 2024, 14, 959. https://doi.org/10.3390/ani14060959
Liu W, Wang T, Zhao K, Hanigan MD, Lin X, Hu Z, Hou Q, Wang Y, Wang Z. Effects of Individual Essential Amino Acids on Growth Rates of Young Rats Fed a Low-Protein Diet. Animals. 2024; 14(6):959. https://doi.org/10.3390/ani14060959
Chicago/Turabian StyleLiu, Wei, Tianyi Wang, Kai Zhao, Mark D. Hanigan, Xueyan Lin, Zhiyong Hu, Qiuling Hou, Yun Wang, and Zhonghua Wang. 2024. "Effects of Individual Essential Amino Acids on Growth Rates of Young Rats Fed a Low-Protein Diet" Animals 14, no. 6: 959. https://doi.org/10.3390/ani14060959
APA StyleLiu, W., Wang, T., Zhao, K., Hanigan, M. D., Lin, X., Hu, Z., Hou, Q., Wang, Y., & Wang, Z. (2024). Effects of Individual Essential Amino Acids on Growth Rates of Young Rats Fed a Low-Protein Diet. Animals, 14(6), 959. https://doi.org/10.3390/ani14060959