


Supplementation of Foals with a Saccharomyces cerevisiae Fermentation Product Alters the Early Response to Vaccination


Abstract
:Simple Summary
Abstract
1. Introduction
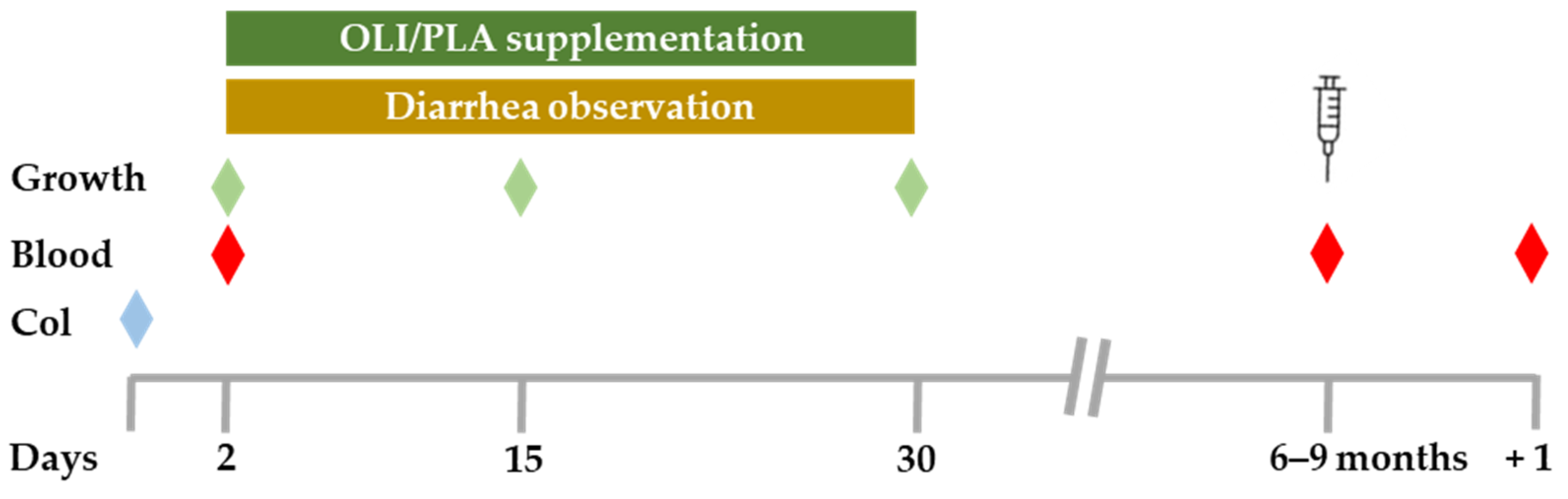
2. Materials and Methods
2.1. Animals and Diet
2.2. Growth and Diarrhea Observation
2.3. Colostrum and Blood Sample Collection
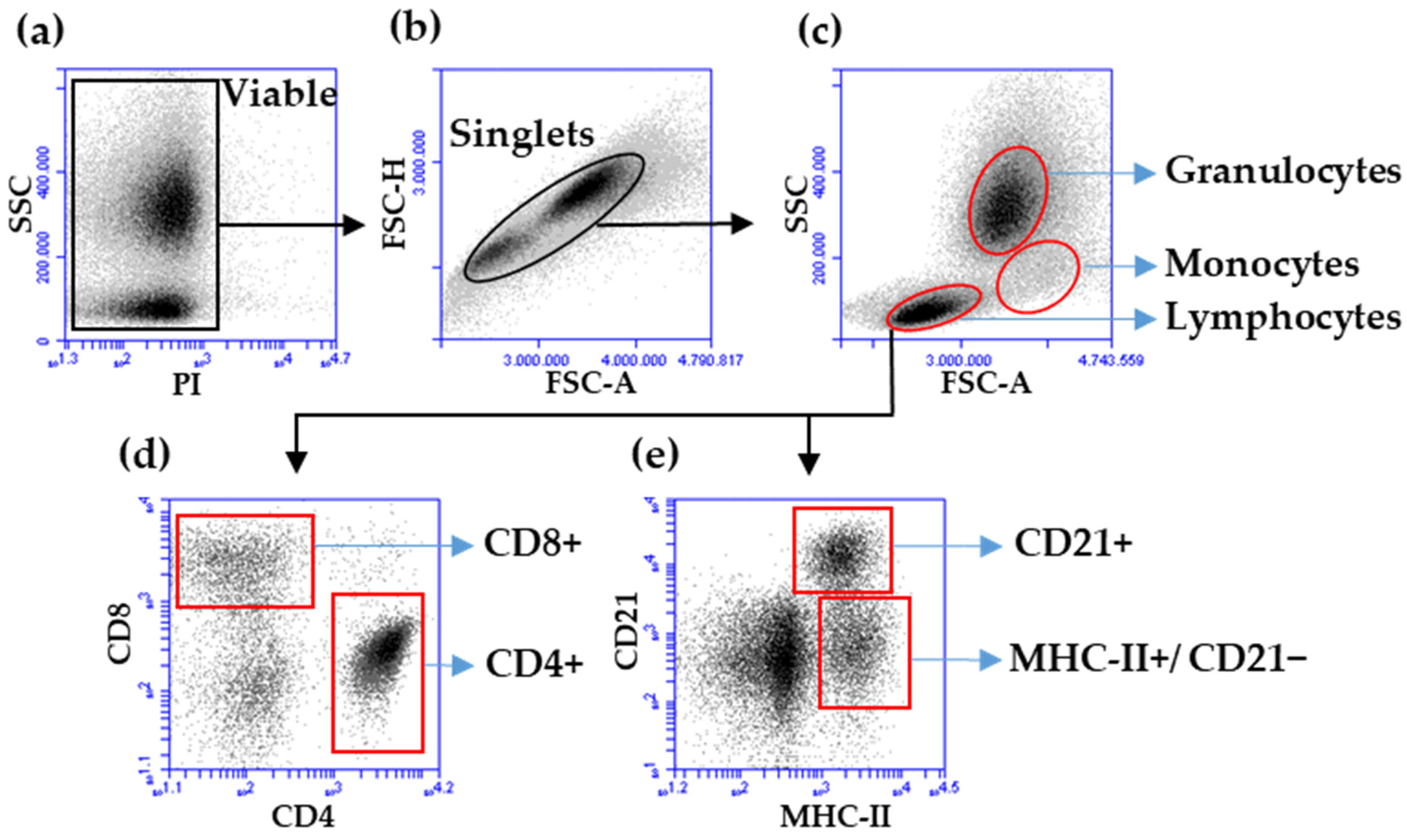
2.4. Determination of Total Leukocyte Counts and Leukocyte Subpopulations
2.5. Membrane Immunofluorescence
2.6. Vaccination
2.7. Statistical Analysis
3. Results
3.1. SCFP Feeding Did Not Affect the Growth of the Foals
3.2. SCFP Feeding Did Not Alter the Duration and the Severity of Foal Heat Diarrhea
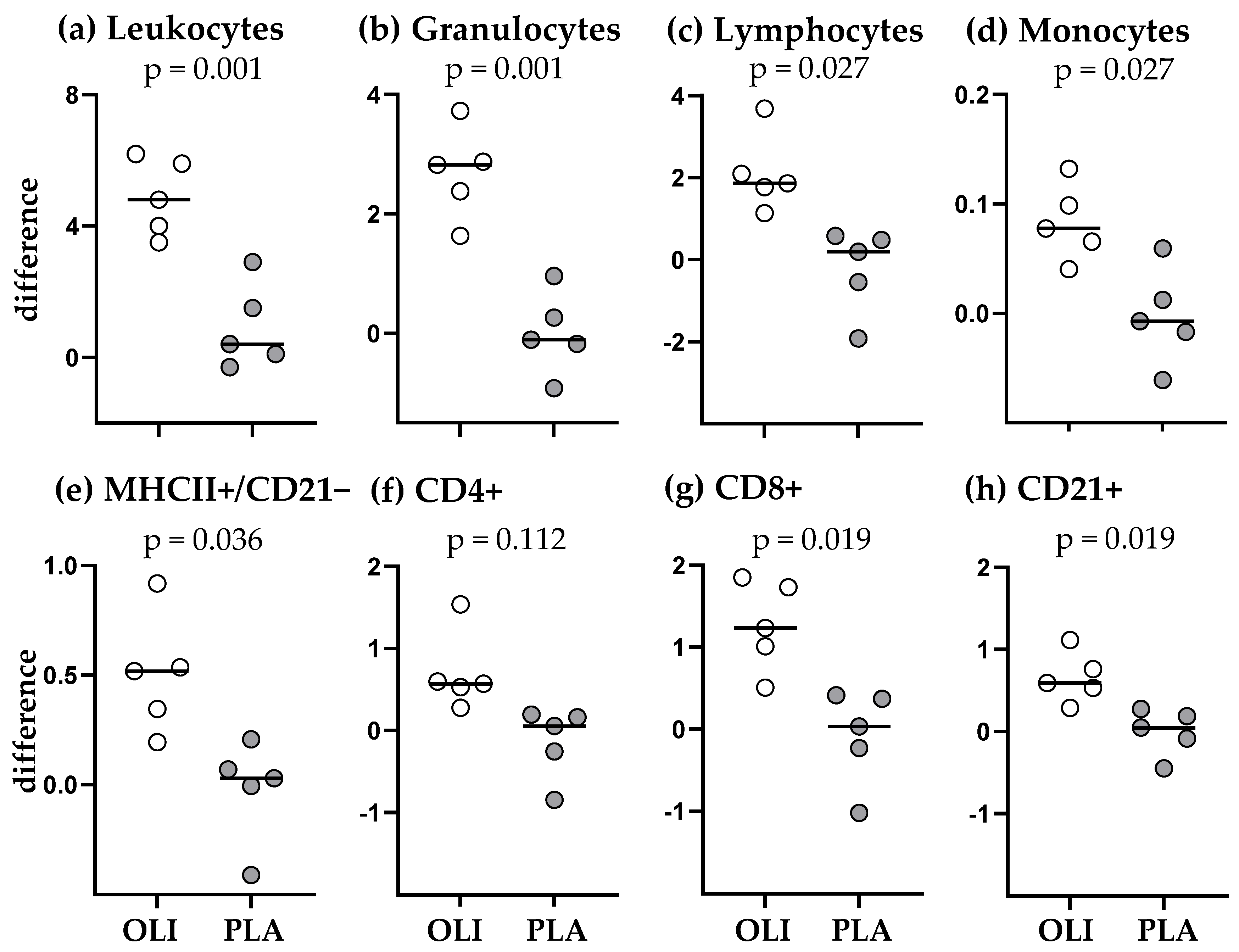
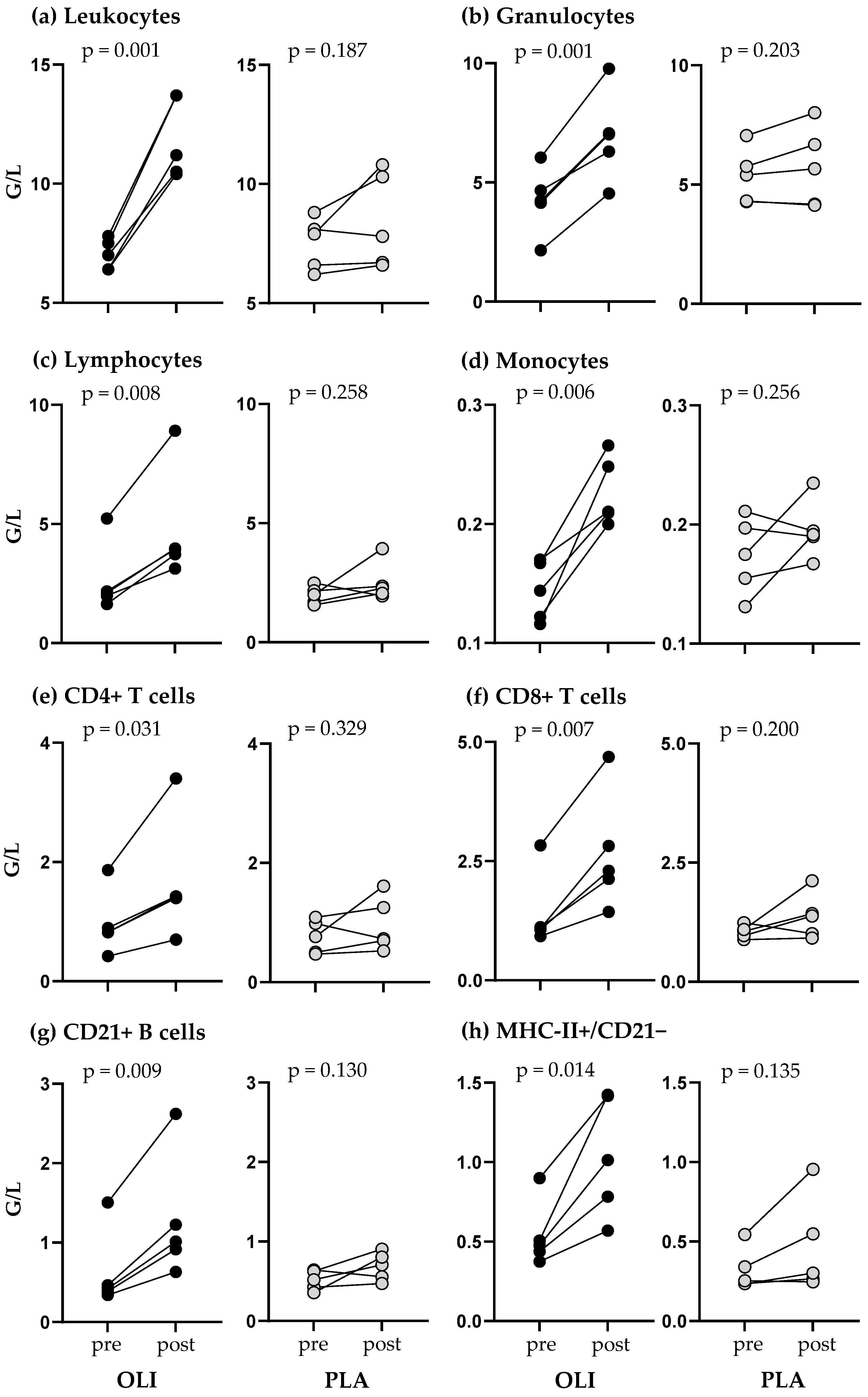
3.3. SCFP Feeding Altered the Immediate Response to Vaccination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ströbel, C.; Günther, E.; Romanowski, K.; Büsing, K.; Urubschurov, V.; Zeyner, A. Effects of oral supplementation of probiotic strains of Lactobacillus rhamnosus and Enterococcus faecium on diarrhoea events of foals in their first weeks of life. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1357–1365. [Google Scholar] [CrossRef]
- Kuhl, J.; Winterhoff, N.; Wulf, M.; Schweigert, F.J.; Schwendenwein, I.; Bruckmaier, R.M.; Aurich, J.E.; Kutzer, P.; Aurich, C. Changes in faecal bacteria and metabolic parameters in foals during the first six weeks of life. Vet. Microbiol. 2011, 151, 321–328. [Google Scholar] [CrossRef]
- Schoster, A.; Staempfli, H.R.; Guardabassi, L.G.; Jalali, M.; Weese, J.S. Comparison of the fecal bacterial microbiota of healthy and diarrheic foals at two and four weeks of life. BMC Vet. Res. 2017, 13, 144. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, U.; Henderson, J.D.; Furtado, K.L.; Pedroja, M.; Elenamarie, O.; Mora, A.; Pechanec, M.Y.; Maga, E.A.; Mienaltowski, M.J. Utilizing the fecal microbiota to understand foal gut transitions from birth to weaning. PLoS ONE 2019, 14, e0216211. [Google Scholar] [CrossRef]
- Masri, M.D.; Merritt, A.M.; Gronwall, R.; Burrows, C.F. Faecal composition in foal heat diarrhoea. Equine Vet. J. 1986, 18, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Magdesian, K.G. Neonatal foal diarrhea. Vet. Clin. N. Am. Equine Pract. 2005, 21, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Oliver-Espinosa, O. Foal Diarrhea: Established and Postulated Causes, Prevention, Diagnostics, and Treatments. Vet. Clin. N. Am. Equine Pract. 2018, 34, 55–68. [Google Scholar] [CrossRef]
- Tremblay, A.; Bronner, S.; Binda, S. Review and Perspectives on Bifidobacterium lactis for Infants’ and Children’s Health. Microorganisms 2023, 11, 2501. [Google Scholar] [CrossRef]
- Kesavelu, D.; Jog, P. Current understanding of antibiotic-associated dysbiosis and approaches for its management. Ther. Adv. Infect. Dis. 2023, 10, 20499361231154443. [Google Scholar] [CrossRef]
- Lynn, M.A.; Tumes, D.J.; Choo, J.M.; Sribnaia, A.; Blake, S.J.; Leong, L.E.X.; Young, G.P.; Marshall, H.S.; Wesselingh, S.L.; Rogers, G.B.; et al. Early-Life Antibiotic-Driven Dysbiosis Leads to Dysregulated Vaccine Immune Responses in Mice. Cell Host Microbe 2018, 23, 653–660.e5. [Google Scholar] [CrossRef]
- John, J.; Roediger, K.; Schroedl, W.; Aldaher, N.; Vervuert, I. Development of intestinal microflora and occurrence of diarrhoea in sucking foals: Effects of Bacillus cereus var. toyoi supplementation. BMC Vet. Res. 2015, 11, 34. [Google Scholar] [CrossRef]
- Schoster, A.; Staempfli, H.R.; Abrahams, M.; Jalali, M.; Weese, J.S.; Guardabassi, L. Effect of a probiotic on prevention of diarrhea and Clostridium difficile and Clostridium perfringens shedding in foals. J. Vet. Intern. Med. 2015, 29, 925–931. [Google Scholar] [CrossRef]
- Urubschurov, V.; Stroebel, C.; Günther, E.; Romanowski, K.; Büsing, K.; Zeyner, A. Effect of oral supplementation of probiotic strains of Lactobacillus rhamnosus and Enterococcus faecium on the composition of the faecal microbiota of foals. J. Anim. Physiol. Anim. Nutr. 2019, 103, 915–924. [Google Scholar] [CrossRef]
- Yuyama, T.; Yusa, S.; Takai, S.; Tsubaki, S.; Kado, Y.; Morotomi, M. Evaluation of a host-specific Lactobacillus probiotic in neonatal foals. J. Appl. Res. Vet. Med. 2004, 2, 26. [Google Scholar]
- Tanabe, S.; Suzuki, T.; Wasano, Y.; Nakajima, F.; Kawasaki, H.; Tsuda, T.; Nagamine, N.; Tsurumachi, T.; Sugaya, K.; Akita, H.; et al. Anti-inflammatory and Intestinal Barrier-protective Activities of Commensal Lactobacilli and Bifidobacteria in Thoroughbreds: Role of Probiotics in Diarrhea Prevention in Neonatal Thoroughbreds. J. Equine Sci. 2014, 25, 37–43. [Google Scholar] [CrossRef]
- Rivulgo, V.M.; Ceci, M.; Haeublin, G.; Sparo, M.; Sanchez Bruni, S. Efficacy of the probiotic strain Enterococcus faecalis CECT7121 in diarrhoea prevention in newborn foals. Rev. Vet. 2016, 27, 3–6. [Google Scholar]
- Alugongo, G.M.; Xiao, J.; Wu, Z.; Li, S.; Wang, Y.; Cao, Z. Review: Utilization of yeast of Saccharomyces cerevisiae origin in artificially raised calves. J. Anim. Sci. Biotechnol. 2017, 8, 34. [Google Scholar] [CrossRef]
- Kiarie, E.; Scott, M.; Krause, D.O.; Khazanehei, H.; Khafipour, E.; Nyachoti, C.M. Interactions of Saccharomyces cerevisiae fermentation product and in-feed antibiotic on gastrointestinal and immunological responses in piglets challenged with Escherichia coli K88+. J. Anim. Sci. 2012, 90 (Suppl. S4), 1–3. [Google Scholar] [CrossRef]
- Shen, Y.B.; Carroll, J.A.; Yoon, I.; Mateo, R.D.; Kim, S.W. Effects of supplementing Saccharomyces cerevisiae fermentation product in sow diets on performance of sows and nursing piglets1,2. J. Anim. Sci. 2011, 89, 2462–2471. [Google Scholar] [CrossRef]
- Yan, H.; Xing, Q.; Xiao, X.; Yu, B.; He, J.; Mao, X.; Yu, J.; Zheng, P.; Luo, Y.; Wu, A.; et al. Effect of Saccharomyces cerevisiae Postbiotics and Essential Oil on Growth Performance and Intestinal Health of Weanling Pigs during K88 ETEC Infection. J. Anim. Sci. 2024, 102, skae007. [Google Scholar] [CrossRef]
- Magalhães, V.J.; Susca, F.; Lima, F.S.; Branco, A.F.; Yoon, I.; Santos, J.E. Effect of feeding yeast culture on performance, health, and immunocompetence of dairy calves. J. Dairy Sci. 2008, 91, 1497–1509. [Google Scholar] [CrossRef]
- Pisoni, L.; Relling, A.E. The effects of supplementing yeast fermentation products on gut permeability, hormone concentration, and growth in newborn dairy calves. Transl. Anim. Sci. 2020, 4, txaa004. [Google Scholar] [CrossRef]
- Burdick Sanchez, N.C.; Carroll, J.A.; Broadway, P.R.; Bass, B.E.; Frank, J.W. Modulation of the acute phase response following a lipopolysaccharide challenge in pigs supplemented with an all-natural Saccharomyces cerevisiae fermentation product. Livest. Sci. 2018, 208, 1–4. [Google Scholar] [CrossRef]
- Chou, W.K.; Park, J.; Carey, J.B.; McIntyre, D.R.; Berghman, L.R. Immunomodulatory Effects of Saccharomyces cerevisiae Fermentation Product Supplementation on Immune Gene Expression and Lymphocyte Distribution in Immune Organs in Broilers. Front. Vet. Sci. 2017, 4, 37. [Google Scholar] [CrossRef]
- Lin, C.Y.; Alexander, C.; Steelman, A.J.; Warzecha, C.M.; de Godoy, M.R.C.; Swanson, K.S. Effects of a Saccharomyces cerevisiae fermentation product on fecal characteristics, nutrient digestibility, fecal fermentative end-products, fecal microbial populations, immune function, and diet palatability in adult dogs1. J. Anim. Sci. 2019, 97, 1586–1599. [Google Scholar] [CrossRef]
- Mahmoud, A.H.A.; Slate, J.R.; Hong, S.; Yoon, I.; McGill, J.L. Supplementing a Saccharomyces cerevisiae fermentation product modulates innate immune function and ameliorates bovine respiratory syncytial virus infection in neonatal calves. J. Anim. Sci. 2020, 98, skaa252. [Google Scholar] [CrossRef]
- Klopp, R.N.; Yoon, I.; Eicher, S.; Boerman, J.P. Effects of feeding Saccharomyces cerevisiae fermentation products on the health of Holstein dairy calves following a lipopolysaccharide challenge. J. Dairy Sci. 2022, 105, 1469–1479. [Google Scholar] [CrossRef]
- Cortés-Coronado, R.F.; Gómez-Rosales, S.; Angeles, M.d.L.; Casaubon-Huguenin, M.T.; Sørensen-Dalgaard, T. Influence of a yeast fermented product on the serum levels of the mannan-binding lectin and the antibodies against the Newcastle disease virus in Ross broilers. J. Appl. Poult. Res. 2017, 26, 38–49. [Google Scholar] [CrossRef]
- Lucassen, A.; Finkler-Schade, C.; Schuberth, H.J. A Saccharomyces cerevisiae Fermentation Product (Olimond BB) Alters the Early Response after Influenza Vaccination in Racehorses. Animals 2021, 11, 2726. [Google Scholar] [CrossRef]
- Deters, E.L.; Stokes, R.S.; Genther-Schroeder, O.N.; Hansen, S.L. Effects of a Saccharomyces cerevisiae fermentation product in receiving diets of newly weaned beef steers. II. Digestibility and response to a vaccination challenge1. J. Anim. Sci. 2018, 96, 3906–3915. [Google Scholar] [CrossRef]
- Valigura, H.C.; Leatherwood, J.L.; Martinez, R.E.; Norton, S.A.; White-Springer, S.H. Dietary supplementation of a Saccharomyces cerevisiae fermentation product attenuates exercise-induced stress markers in young horses. J. Anim. Sci. 2021, 99, skab199. [Google Scholar] [CrossRef]
- Lucassen, A.; Hankel, J.; Finkler-Schade, C.; Osbelt, L.; Strowig, T.; Visscher, C.; Schuberth, H.J. Feeding a Saccharomyces cerevisiae Fermentation Product (Olimond BB) Does Not Alter the Fecal Microbiota of Thoroughbred Racehorses. Animals 2022, 12, 1496. [Google Scholar] [CrossRef]
- Trckova, M.; Faldyna, M.; Alexa, P.; Zajacova, Z.S.; Gopfert, E.; Kumprechtova, D.; Auclair, E.; D’Inca, R. The effects of live yeast Saccharomyces cerevisiae on postweaning diarrhea, immune response, and growth performance in weaned piglets1,2. J. Anim. Sci. 2014, 92, 767–774. [Google Scholar] [CrossRef]
- Lynn, D.J.; Benson, S.C.; Lynn, M.A.; Pulendran, B. Modulation of immune responses to vaccination by the microbiota: Implications and potential mechanisms. Nat. Rev. Immunol. 2022, 22, 33–46. [Google Scholar] [CrossRef]
- Schneider, F.; Wehrend, A. Quality Assessment of Bovine and Equine Colostrum—An Overview. Schweiz. Arch. Tierheilkd. 2019, 161, 287–297. [Google Scholar] [CrossRef]
- Quigley, J.D., 3rd; Wallis, L.B.; Dowlen, H.H.; Heitmann, R.N. Sodium bicarbonate and yeast culture effects on ruminal fermentation, growth, and intake in dairy calves. J. Dairy Sci. 1992, 75, 3531–3538. [Google Scholar] [CrossRef]
- Seymour, W.M.; Nocek, J.E.; Siciliano-Jones, J. Effects of a colostrum substitute and of dietary brewer’s yeast on the health and performance of dairy calves. J. Dairy Sci. 1995, 78, 412–420. [Google Scholar] [CrossRef]
- Brewer, M.T.; Anderson, K.L.; Yoon, I.; Scott, M.F.; Carlson, S.A. Amelioration of salmonellosis in pre-weaned dairy calves fed Saccharomyces cerevisiae fermentation products in feed and milk replacer. Vet. Microbiol. 2014, 172, 248–255. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand Timothy, W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef]
- Andersen-Nissen, E.; Fiore-Gartland, A.; Ballweber Fleming, L.; Carpp, L.N.; Naidoo, A.F.; Harper, M.S.; Voillet, V.; Grunenberg, N.; Laher, F.; Innes, C.; et al. Innate immune signatures to a partially-efficacious HIV vaccine predict correlates of HIV-1 infection risk. PLoS Pathog. 2021, 17, e1009363. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.Y.; Hanson, N.Q.; Straka, R.J.; Hoke, T.R.; Ordovas, J.M.; Peacock, J.M.; Arends, V.L.; Arnett, D.K. Effect of influenza vaccine on markers of inflammation and lipid profile. J. Lab. Clin. Med. 2005, 145, 323–327. [Google Scholar] [CrossRef]
- Arunachalam, P.S.; Scott, M.K.D.; Hagan, T.; Li, C.; Feng, Y.; Wimmers, F.; Grigoryan, L.; Trisal, M.; Edara, V.V.; Lai, L.; et al. Systems biological assessment of human immunity to BNT162b2 mRNA vaccination. Res. Sq. 2021. preprint. [Google Scholar] [CrossRef]
- Pabst, O.; Hornef, M. Gut microbiota: A natural adjuvant for vaccination. Immunity 2014, 41, 349–351. [Google Scholar] [CrossRef]
- Valdez, Y.; Brown, E.M.; Finlay, B.B. Influence of the microbiota on vaccine effectiveness. Trends Immunol. 2014, 35, 526–537. [Google Scholar] [CrossRef]
- Wu, W.; Chen, Z.; Han, J.; Qian, L.; Wang, W.; Lei, J.; Wang, H. Endocrine, genetic, and microbiome nexus of obesity and potential role of postbiotics: A narrative review. Eat Weight Disord. 2023, 28, 84. [Google Scholar] [CrossRef]
- Di Costanzo, M.; De Paulis, N.; Capra, M.E.; Biasucci, G. Nutrition during Pregnancy and Lactation: Epigenetic Effects on Infants’ Immune System in Food Allergy. Nutrients 2022, 14, 1766. [Google Scholar] [CrossRef]
- Cuevas-Sierra, A.; Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, and Obesity: Links with Host Genetics and Epigenetics and Potential Applications. Adv. Nutr. 2019, 10, S17–S30. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | OLI 1 (10 g) | PLA 2 (10 g) |
|---|---|---|
| Tocopherol extract | 0.045 g | 0.045 g |
| Coconut oil (rapeseed oil) | - | 0.3 g |
| Vitamin C | 0.292 g | 0.292 g |
| Dextrose | - | 2.00 g |
| Corn cob meal | - | 1.5 g |
| Linseed cake | - | 1.5 g |
| Microcrystalline cellulose | - | 2.00 g |
| Minerals | 0.063 g | 2.363 g |
| Inactivated yeasts | 9.6 g | - |
| Group 1 (OLI) (n = 11) | Group 2 (PLA) (n = 10) | p c | ||
|---|---|---|---|---|
| Brix (%) | 23.5 (17–32) | 25.0 (22–33) | 0.1923 | |
| Serum protein (g/dL) a | 6.0 (4.6–6.6) | 6.4 (5.6–7.2) | 0.3567 | |
| Height (cm) b | d2 | 105.0 ± 1.0 | 104.0 ± 4.25 | 0.6690 |
| d15 | 110.0 ± 2.5 | 111.0 ± 2.5 | 0.5685 | |
| d30 | 115.0 ± 2.5 | 115.5 ± 5.25 | 0.5708 | |
| Weight (kg) b | d2 | 59.6 ± 7.45 | 58.4 ± 8.55 | 0.6985 |
| d15 | 82.7 ± 8.35 | 80.9 ± 10.925 | 0.8602 | |
| d30 | 101.0 ± 12.85 | 98.1 ± 16.25 | 0.5974 | |
| Group 1 (OLI) (n = 11) | Group 2 (PLA) (n = 10) | p | |
|---|---|---|---|
| Diarrhea days a | 2.0 ± 4.5 | 6.0 ± 5.5 | 0.1347 |
| Diarrhea severity b | 8.0 ± 16.0 | 23.5 ± 21.25 | 0.1288 |
| Diarrhea Group (DG) a | Group 1 (OLI) (n = 11) | Group 2 (PLA) (n = 10) |
|---|---|---|
| DG1 | 8 | 4 |
| DG2 | 3 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terpeluk, E.R.; Schäfer, J.; Finkler-Schade, C.; Schuberth, H.-J. Supplementation of Foals with a Saccharomyces cerevisiae Fermentation Product Alters the Early Response to Vaccination. Animals 2024, 14, 960. https://doi.org/10.3390/ani14060960
Terpeluk ER, Schäfer J, Finkler-Schade C, Schuberth H-J. Supplementation of Foals with a Saccharomyces cerevisiae Fermentation Product Alters the Early Response to Vaccination. Animals. 2024; 14(6):960. https://doi.org/10.3390/ani14060960
Chicago/Turabian StyleTerpeluk, Eva Ronja, Jana Schäfer, Christa Finkler-Schade, and Hans-Joachim Schuberth. 2024. "Supplementation of Foals with a Saccharomyces cerevisiae Fermentation Product Alters the Early Response to Vaccination" Animals 14, no. 6: 960. https://doi.org/10.3390/ani14060960
APA StyleTerpeluk, E. R., Schäfer, J., Finkler-Schade, C., & Schuberth, H. -J. (2024). Supplementation of Foals with a Saccharomyces cerevisiae Fermentation Product Alters the Early Response to Vaccination. Animals, 14(6), 960. https://doi.org/10.3390/ani14060960