Supplementation of Extender with Melatonin Improves the Motility, Mitochondrial Membrane Potential, and Fertilization Ability of Cryopreserved Brown-Marbled Grouper Sperm


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
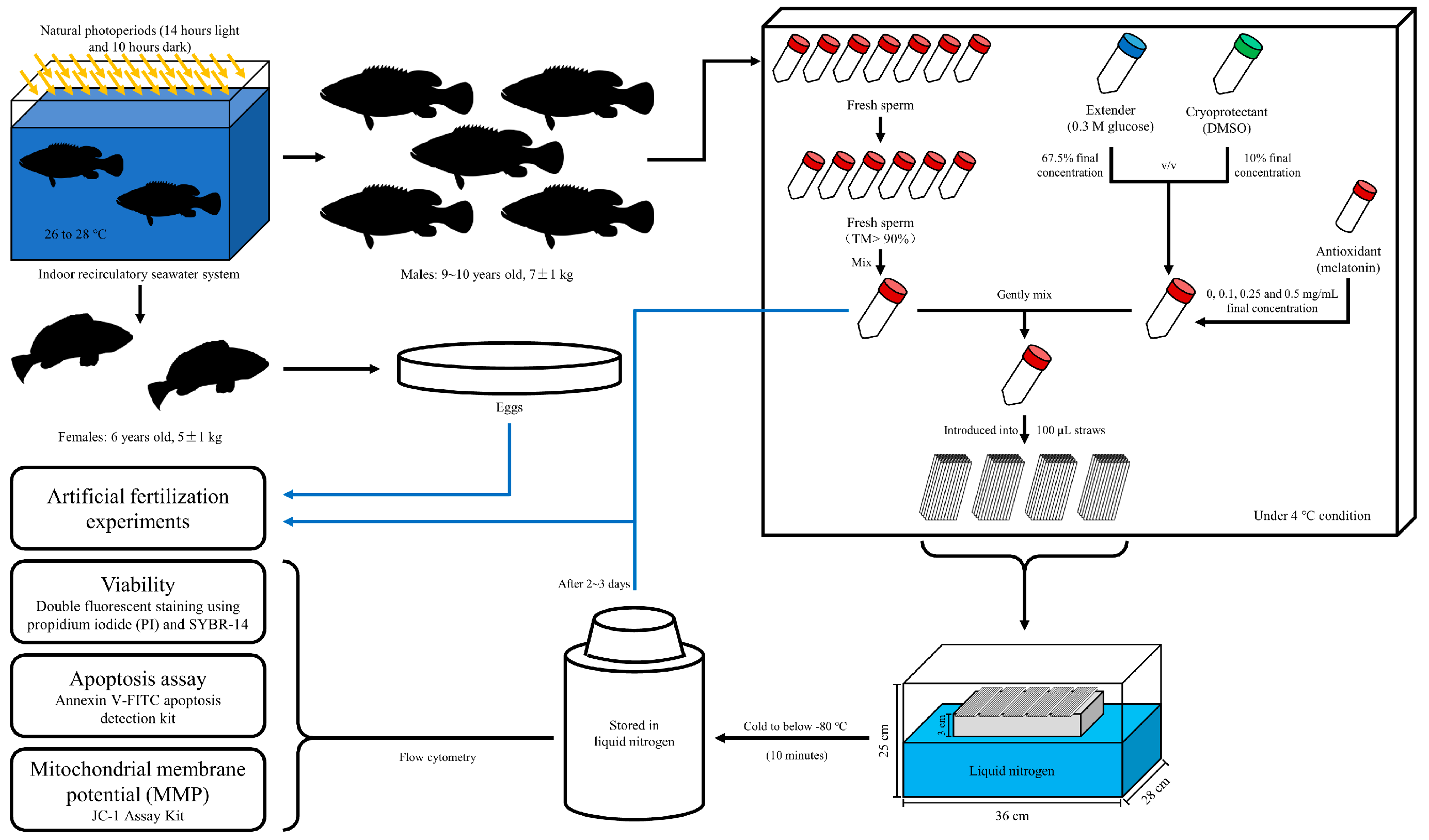
2.1. Broodstock and Gamete Collection
2.2. Preparation and Cryopreservation of Samples
2.3. Sperm Quality Evaluation
2.3.1. Motility Parameters
2.3.2. Viability
2.3.3. Sperm Apoptosis Assay
2.3.4. Mitochondrial Membrane Potential (MMP)
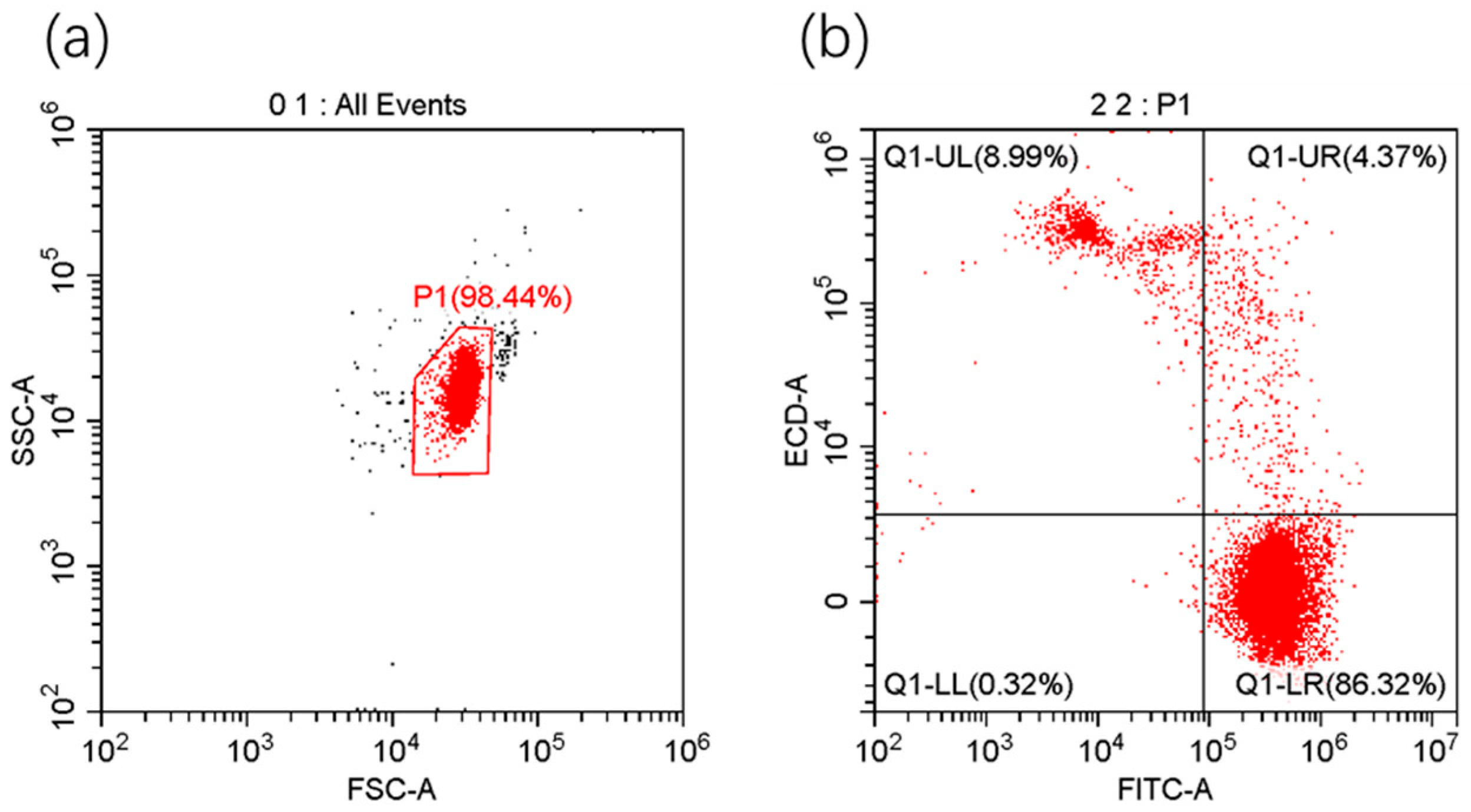
2.3.5. Flow Cytometry
2.4. Artificial Fertilization Experiments
2.5. Statistical Analysis
3. Results
3.1. Motility Parameters
3.2. Viability, Apoptosis Rate, and MMP
3.3. Artificial Fertilization Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rhodes, K.; Sadovy, Y.; Samoilys, M. Epinephelus fuscoguttatus; e.T44673A100468078; The IUCN Red List of Threatened Species: Cambridge, UK, 2018. [Google Scholar] [CrossRef]
- Che-Zulkifli, C.I.; Koh, I.C.C.; Shahreza, M.S.; Ikhwanuddin, M. Cryopreservation of spermatozoa on grouper species: A review. Rev. Aquac. 2020, 12, 26–32. [Google Scholar] [CrossRef]
- Yang, S.; Fan, B.; Chen, X.; Meng, Z. Cryopreservation of sperm in brown-marbled grouper (Epinephelus fuscoguttatus). Aquac. Int. 2020, 28, 1501–1516. [Google Scholar] [CrossRef]
- Yang, S.; Fan, B.; Chen, X.; Meng, Z. Supplementation of the freezing medium with Coenzyme Q10 attenuates oxidative stress and improves function of frozen-thawed giant grouper (Epinephelus lanceolatus) spermatozoa. Theriogenology 2021, 175, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Yang, S.; Wang, L.; Chen, X.; Liu, X.; Zhang, Y.; Li, S.; Zhang, H.; Meng, Z.; Lin, H. Hybridization of tiger grouper (Epinephelus fuscoguttatus female symbol) x giant grouper (Epinephelus lanceolatus male symbol) using cryopreserved sperm. Cryobiology 2020, 95, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Jiang, J.; Wang, N.; Qi, W.; Zhai, J.; Li, B.; Liang, Y.; Chen, Y.; Yang, C.; Chen, S. Sperm of the giant grouper: Cryopreservation, physiological and morphological analysis and application in hybridizations with red-spotted grouper. J. Reprod. Dev. 2015, 61, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, T.; Chen, J.F.; Wu, X.; Wu, L.N.; Zhang, W.W.; Luo, J.; Xia, J.H.; Meng, Z.N.; Liu, X.C. First construction of interspecific backcross grouper and genome-wide identification of their genetic variants associated with early growth. Aquaculture 2021, 545, 737221. [Google Scholar] [CrossRef]
- Wang, D.; Wu, F. 2021 China Fisher Statistical Yearbook; China Agriculture Press: Beijing, China, 2021. [Google Scholar]
- Yang, Y.; Wu, L.N.; Chen, J.F.; Wu, X.; Xia, J.H.; Meng, Z.N.; Liu, X.C.; Lin, H.-R. Whole-genome sequencing of leopard coral grouper (Plectropomus leopardus) and exploration of regulation mechanism of skin color and adaptive evolution. Zool. Res. 2020, 41, 328. [Google Scholar] [CrossRef]
- Guo, L.; Xia, J.; Yang, S.; Li, M.; You, X.; Meng, Z.; Lin, H. GHRH, PRP-PACAP and GHRHR target sequencing via an ion torrent personal genome machine reveals an association with growth in orange-spotted grouper (Epinephelus coioides). Int. J. Mol. Sci. 2015, 16, 26137–26150. [Google Scholar] [CrossRef]
- Sandoval-Vargas, L.; Silva Jiménez, M.; Risopatrón González, J.; Villalobos, E.F.; Cabrita, E.; Valdebenito Isler, I. Oxidative stress and use of antioxidants in fish semen cryopreservation. Rev. Aquac. 2020, 13, 365–387. [Google Scholar] [CrossRef]
- Chauvigné, F.; Boj, M.; Finn, R.N.; Cerdà, J. Mitochondrial aquaporin-8-mediated hydrogen peroxide transport is essential for teleost spermatozoon motility. Sci. Rep. 2015, 5, 7789. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Mansour, N. A comparative study on antioxidant systems in semen of species of the Percidae, Salmonidae, Cyprinidae, and Lotidae for improving semen storage techniques. Aquaculture 2010, 307, 130–140. [Google Scholar] [CrossRef]
- Li, P.; Xi, M.D.; Du, H.; Qiao, X.M.; Liu, Z.G.; Wei, Q.W. Antioxidant supplementation, effect on post-thaw spermatozoan function in three sturgeon species. Reprod. Domest. Anim. 2018, 53, 287–295. [Google Scholar] [CrossRef]
- Eriani, K.; Mustaqim, M.; Hasri, I.; Amalia, R.; Azhar, A.; Nur, F.M.; Muchlisin, Z.A. Effect of fetal bovine serum on the sperm quality of depik (Rasbora tawarensis) after short-term cryopreservation. Indian J. Anim. Res. 2021, 56, 843–847. [Google Scholar] [CrossRef]
- Ogretmen, F.; Inanan, B.E. Effect of butylated hydroxytoluene (BHT) on the cryopreservation of common carp (Cyprinus carpio) spermatozoa. Anim. Reprod. Sci. 2014, 151, 269–274. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Mansour, N.; Kunz, F.A. The effect of antioxidants on the quality of cryopreserved semen in two salmonid fish, the brook trout (Salvelinus fontinalis) and the rainbow trout (Oncorhynchus mykiss). Theriogenology 2011, 76, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Ekici, A.; Baran, A.; Yamaner, G.; Ozdas, O.B.; Sandal, A.I.; Guven, E.; Baltaci, M.A. Effects of different doses of taurine in the flucose-based extender during cryopreservation of rainbow trout (Oncorhynchus mykiss) semen. Biotechnol. Biotechnol. Equip. 2012, 26, 3113–3115. [Google Scholar] [CrossRef]
- Sarosiek, B.; Judycka, S.; Kowalski, R.K. Influence of antioxidants on spermatozoa in the short-term storage of salmonidae milt. Pol. J. Nat. Sci. 2013, 28, 379–384. [Google Scholar]
- Muthmainnah, C.R.; Eriani, K.; Hasri, I.; Irham, M.; Batubara, A.S.; Muchlisin, Z.A. Effect of glutathione on sperm quality after short-term cryopreservation in seurukan fish Osteochilus vittatus (Cyprinidae). Theriogenology 2018, 122, 30–34. [Google Scholar] [CrossRef] [PubMed]
- El-Raey, M.; Badr, M.R.; Rawash, Z.M.; Darwish, G.M. Evidences for the role of melatonin as a protective additive during buffalo semen freezing. Am. J. Anim. Vet. Sci. 2014, 9, 252–262. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, R.; Lv, Y.; Zeng, W. Melatonin protects rabbit spermatozoa from cryo-damage via decreasing oxidative stress. Cryobiology 2019, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Jozaki, M.; Tanabe, M.; Shirafuta, Y.; Mihara, Y.; Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Taketani, T.; et al. Importance of melatonin in assisted reproductive technology and ovarian aging. Int. J. Mol. Sci. 2020, 21, 1135. [Google Scholar] [CrossRef]
- Lemasters, J.J. Selective mitochondrial autophagy, or mitophagy, as a targeted defense against oxidative stress, mitochondrial dysfunction, and aging. Rejuvenation Res. 2005, 8, 3–5. [Google Scholar] [CrossRef]
- Shahandeh, E.; Ghorbani, M.; Mokhlesabadifarahani, T.; Bardestani, F. Melatonin and selenium supplementation in extenders improves the post-thaw quality parameters of rat sperm. Clin. Exp. Reprod. Med. 2022, 49, 87–92. [Google Scholar] [CrossRef]
- Chandra Chaudhary, S.; Aeksiri, N.; Wanangkarn, A.; Liao, Y.-J.; Inyawilert, W. Effects of melatonin on cryopreserved semen parameters and apoptosis of Thai swamp buffalo bull (Bubalus bubalis) in different thawing conditions. Adv. Anim. Vet. Sci. 2020, 9, 238–245. [Google Scholar] [CrossRef]
- Tanhaei Vash, N.; Nadri, P.; Karimi, A. Synergistic effects of myo-inositol and melatonin on cryopreservation of goat spermatozoa. Reprod. Domest. Anim. 2022, 57, 876–885. [Google Scholar] [CrossRef]
- Mehaisen, G.M.K.; Partyka, A.; Ligocka, Z.; Nizanski, W. Cryoprotective effect of melatonin supplementation on post-thawed rooster sperm quality. Anim. Reprod. Sci. 2020, 212, 106238. [Google Scholar] [CrossRef]
- Najafi, A.; Adutwum, E.; Yari, A.; Salehi, E.; Mikaeili, S.; Dashtestani, F.; Abolhassani, F.; Rashki, L.; Shiasi, S.; Asadi, E. Melatonin affects membrane integrity, intracellular reactive oxygen species, caspase3 activity and AKT phosphorylation in frozen thawed human sperm. Cell Tissue Res. 2018, 372, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.-L.; Sun, T.-C.; Yu, K.; Wang, Z.-P.; Zhang, B.-L.; Zhang, Y.; Wang, X.-X.; Lian, Z.-X.; Liu, Y.-X. Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, C.; Xiang, H.; Jia, G.; Zhong, R. Melatonin improves cryopreservation of ram sperm by inhibiting mitochondrial permeability transition pore opening. Reprod. Domest. Anim. 2020, 55, 1240–1249. [Google Scholar] [CrossRef]
- Lançoni, R.; Celeghini, E.C.C.; Alves, M.B.R.; Lemes, K.M.; Gonella-Diaza, A.M.; Oliveira, L.Z.; Arruda, R.P.d. Melatonin added to cryopreservation extenders improves the mitochondrial membrane potential of postthawed equine sperm. J. Equine Vet. Sci. 2018, 69, 78–83. [Google Scholar] [CrossRef]
- Fadl, A.M.; Ghallab, A.R.M.; Abou-Ahmed, M.M.; Moawad, A.R. Melatonin can improve viability and functional integrity of cooled and frozen/thawed rabbit spermatozoa. Reprod. Domest. Anim. 2021, 56, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Felix, F.; Oliveira, C.C.V.; Cabrita, E. Antioxidants in fish sperm and the potential role of melatonin. Antioxidants 2020, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Alevra, A.I.; Exadactylos, A.; Mente, E.; Papadopoulos, S. The protective role of melatonin in sperm cryopreservation of farm animals and human: Lessons for male fish cryopreservation. Antioxidants 2022, 12, 791. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fan, B.; Chen, X.; Meng, Z. Evaluation of different sugar extenders and large volume cryotube for sperm cryopreservation of brown-marbled grouper ( Epinephelus fuscoguttatus). Aquac. Res. 2022, 53, 4069–4075. [Google Scholar] [CrossRef]
- Yang, S.; Huang, W.; Guo, W.; Chen, X.; Fan, B.; Meng, Z. Effects of gametes post-activation, egg storage and sperm-egg ratio on in vitro fertilization outcomes in the brown-marbled grouper (Epinephelus fuscoguttatus). Anim. Reprod. Sci. 2020, 219, 106529. [Google Scholar] [CrossRef] [PubMed]
- Palhares, P.C.; Assis, I.L.; Machado, G.J.; de Freitas, R.M.P.; de Freitas, M.B.D.; Paula, D.A.J.; Carneiro, W.F.; Motta, N.C.; Murgas, L.D.S. Sperm characteristics, peroxidation lipid and antioxidant enzyme activity changes in milt of Brycon orbignyanus cryopreserved with melatonin in different freezing curves. Theriogenology 2021, 176, 18–25. [Google Scholar] [CrossRef]
- Ofosu, J.; Qazi, I.H.; Fang, Y.; Zhou, G. Use of melatonin in sperm cryopreservation of farm animals: A brief review. Anim. Reprod. Sci. 2021, 233, 106850. [Google Scholar] [CrossRef]
- Cabrita, E.; Ma, S.; Diogo, P.; Martínez-Páramo, S.; Sarasquete, C.; Dinis, M.T. The influence of certain aminoacids and vitamins on post-thaw fish sperm motility, viability and DNA fragmentation. Anim. Reprod. Sci. 2011, 125, 189–195. [Google Scholar] [CrossRef]
- Yang, S.; Zeng, Z.; Chen, X.; Fan, B.; Meng, Z. Melatonin supplementation to the freezing medium enhances post-thaw sperm quality and fertility of giant grouper (Epinephelus lanceolatus). Aquac. Rep. 2023, 31, 101662. [Google Scholar] [CrossRef]
- Gallego, V.; Asturiano, J.F. Fish sperm motility assessment as a tool for aquaculture research: A historical approach. Rev. Aquac. 2019, 11, 697–724. [Google Scholar] [CrossRef]
- Motta, N.C.; Egger, R.C.; Monteiro, K.S.; Vogel de Oliveira, A.; Solis Murgas, L.D. Effects of melatonin supplementation on the quality of cryopreserved sperm in the neotropical fish Prochilodus lineatus. Theriogenology 2022, 179, 14–21. [Google Scholar] [CrossRef]
- Medrano, A.; Contreras, C.; Herrera, F.; Alcantar-Rodriguez, A. Melatonin as an antioxidant preserving sperm from domestic animals. Asian Pac. J. Reprod. 2017, 6, 241–246. [Google Scholar] [CrossRef]
- Leon, J.; Acuna-Castroviejo, D.; Sainz, R.M.; Mayo, J.C.; Tan, D.X.; Reiter, R.J. Melatonin and mitochondrial function. Life Sci. 2004, 75, 765–790. [Google Scholar] [CrossRef]
- Treulen, F.; Arias, M.E.; Aguila, L.; Uribe, P.; Felmer, R. Cryopreservation induces mitochondrial permeability transition in a bovine sperm model. Cryobiology 2018, 83, 65–74. [Google Scholar] [CrossRef]
- Crompton, M. The mitochondrial permeability transition pore and its role in cell death. Biochem. J. 1999, 341, 233–249. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, C.; Xiang, H.; Zhao, X.; Zhong, R. Melatonin inhibits formation of mitochondrial permeability transition pores and improves oxidative phosphorylation of frozen-thawed ram sperm. Front. Endocrinol. 2020, 10, 491736. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.-Y.; Li, Q.; Ren, F.; Xi, H.-M.; Lv, D.-L.; Li, Y.; Hu, J.-H. Melatonin Protects Goat Spermatogonial Stem Cells against Oxidative Damage during Cryopreservation by Improving Antioxidant Capacity and Inhibiting Mitochondrial Apoptosis Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 5954635. [Google Scholar] [CrossRef] [PubMed]
- Palhares, P.C.; Assis, I.L.; Souza, J.; Franca, T.S.; Egger, R.C.; Paula, D.A.J.; Murgas, L.D.S. Effect of melatonin supplementation to a cytoprotective medium on post-thawed Brycon orbignyanus sperm quality preserved during different freezing times. Cryobiology 2020, 96, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin protects ram spermatozoa from cryopreservation injuries in a dose-dependent manner. J. Pineal Res. 2011, 50, 310–318. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Melatonin (mg/mL) | TM (%) | PM (%) | VCL (μm/s) | VSL (μm/s) | VAP (μm/s) |
|---|---|---|---|---|---|
| Fresh | 93.85 ± 0.94 a | 28.12 ± 3.10 | 124.62 ± 6.01 a | 44.29 ± 2.91 a | 67.66 ± 4.25 ab |
| 0 (Control) | 76.50 ± 1.10 c | 23.88 ± 2.35 | 85.81 ± 5.01 c | 49.23 ± 6.89 ac | 62.56 ± 6.54 b |
| 0.1 | 78.86 ± 3.59 bc | 28.28 ± 3.77 | 91.90 ± 2.65 b | 58.55 ± 3.32 b | 70.63 ± 2.86 a |
| 0.25 | 81.16 ± 4.08 b | 29.49 ± 1.98 | 92.26 ± 2.13 b | 51.96 ± 6.02 c | 67.00 ± 4.73 ab |
| 0.5 | 76.49 ± 1.23 c | 26.56 ± 3.01 | 90.35 ± 3.35 ab | 50.39 ± 5.22 ac | 65.27 ± 4.49 ab |
| Melatonin (mg/mL) | Viability (%) | Apoptosis Rate (%) | High MMP (%) |
|---|---|---|---|
| 0 | 85.8 ± 1.5 | 16.8 ± 2.1 | 77.1 ± 6.6 a |
| 0.1 | 86.4 ± 1.6 | 15.3 ± 0.6 | 89.0 ± 5.0 ab |
| 0.25 | 87.4 ± 1.3 | 15.7 ± 1.1 | 91.5 ± 3.9 b |
| 0.5 | 83.6 ± 1.4 | 15.9 ± 2.9 | 86.1 ± 4.8 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, Q.; Yang, S.; Hua, S.; Zhang, W.; Li, D.; Yang, Y.; Wang, X.; Wang, Q.; Meng, Z. Supplementation of Extender with Melatonin Improves the Motility, Mitochondrial Membrane Potential, and Fertilization Ability of Cryopreserved Brown-Marbled Grouper Sperm. Animals 2024, 14, 995. https://doi.org/10.3390/ani14070995
Ruan Q, Yang S, Hua S, Zhang W, Li D, Yang Y, Wang X, Wang Q, Meng Z. Supplementation of Extender with Melatonin Improves the Motility, Mitochondrial Membrane Potential, and Fertilization Ability of Cryopreserved Brown-Marbled Grouper Sperm. Animals. 2024; 14(7):995. https://doi.org/10.3390/ani14070995
Chicago/Turabian StyleRuan, Qingxin, Sen Yang, Sijie Hua, Weiwei Zhang, Duo Li, Yang Yang, Xi Wang, Qinghua Wang, and Zining Meng. 2024. "Supplementation of Extender with Melatonin Improves the Motility, Mitochondrial Membrane Potential, and Fertilization Ability of Cryopreserved Brown-Marbled Grouper Sperm" Animals 14, no. 7: 995. https://doi.org/10.3390/ani14070995
APA StyleRuan, Q., Yang, S., Hua, S., Zhang, W., Li, D., Yang, Y., Wang, X., Wang, Q., & Meng, Z. (2024). Supplementation of Extender with Melatonin Improves the Motility, Mitochondrial Membrane Potential, and Fertilization Ability of Cryopreserved Brown-Marbled Grouper Sperm. Animals, 14(7), 995. https://doi.org/10.3390/ani14070995