Effects of Dietary Fatty Acids on Lipid Traits in the Muscle and Perirenal Fat of Growing Rabbits Fed Mixed Diets

Abstract
:Simple Summary
Abstract
1. Introduction
2. Experimental Section

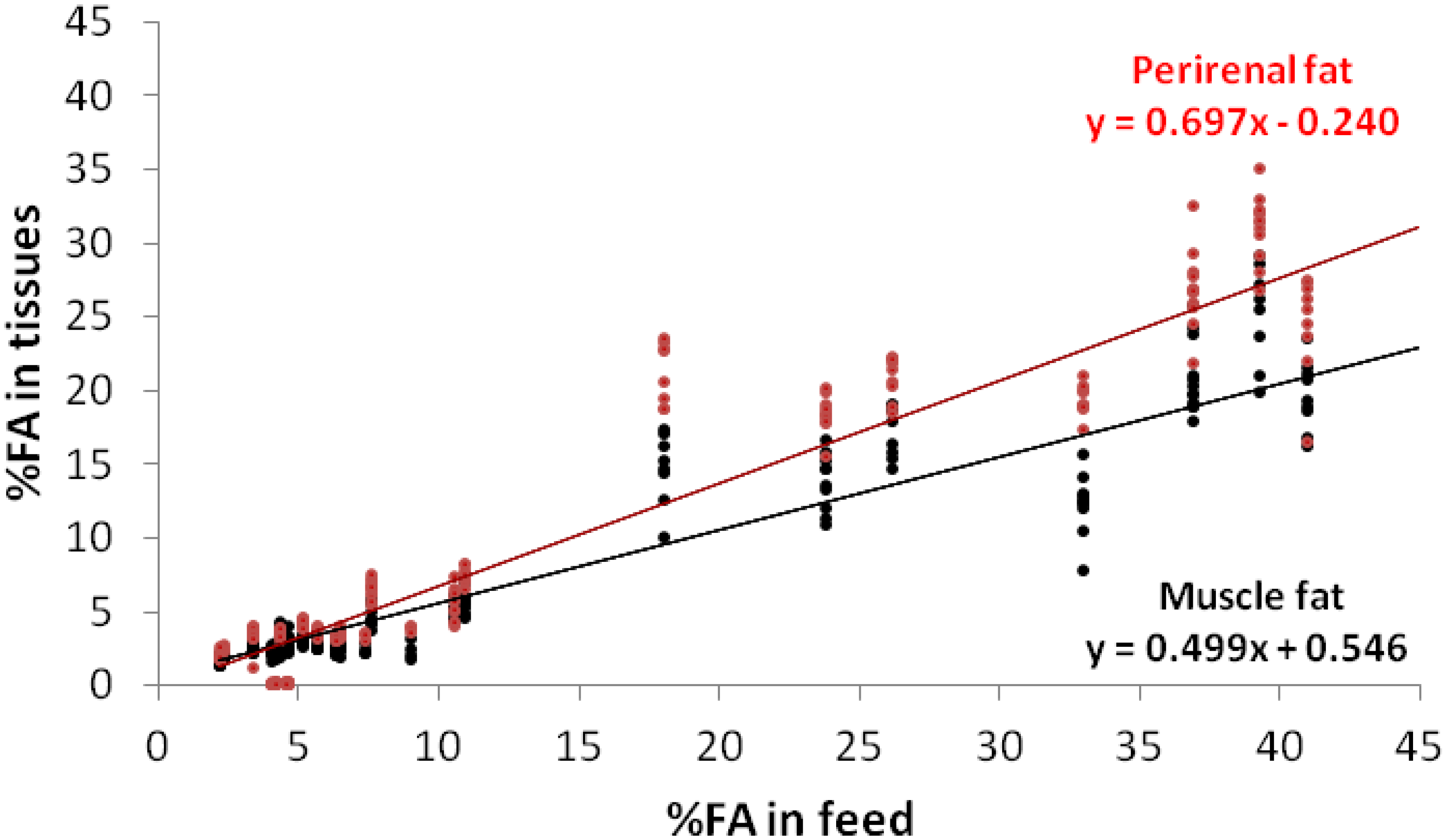{kind=link}
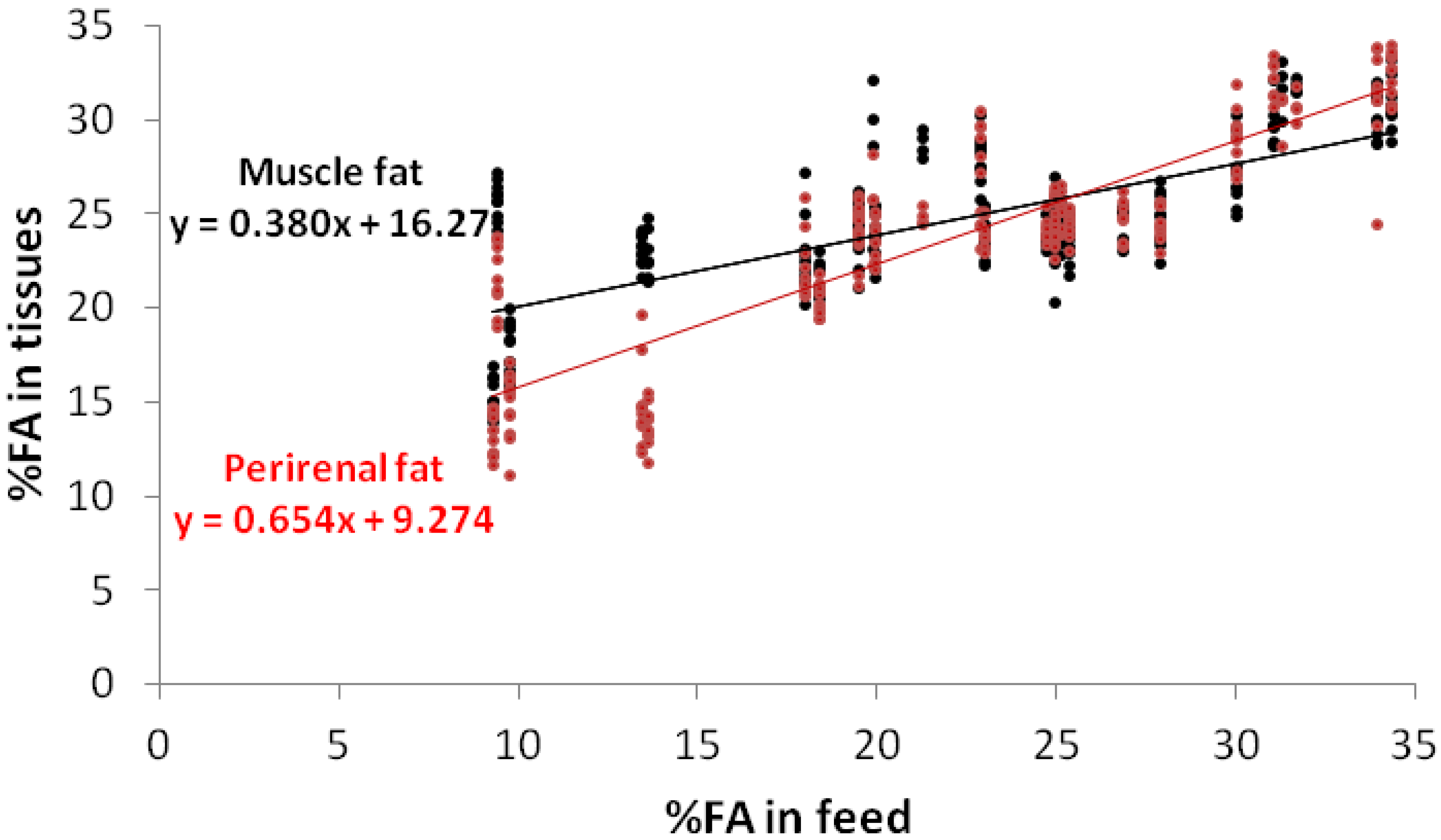{kind=link}
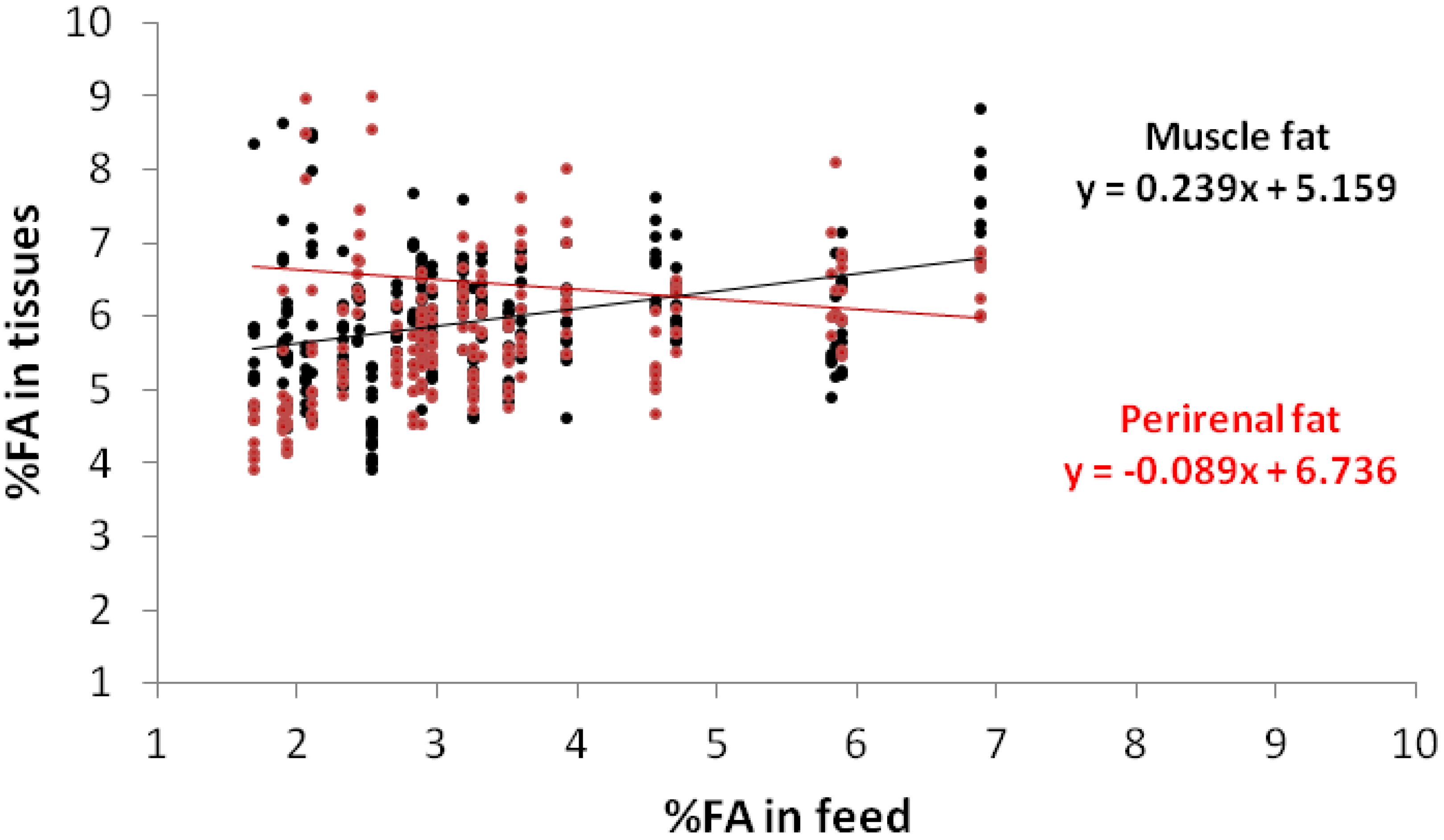{kind=link}
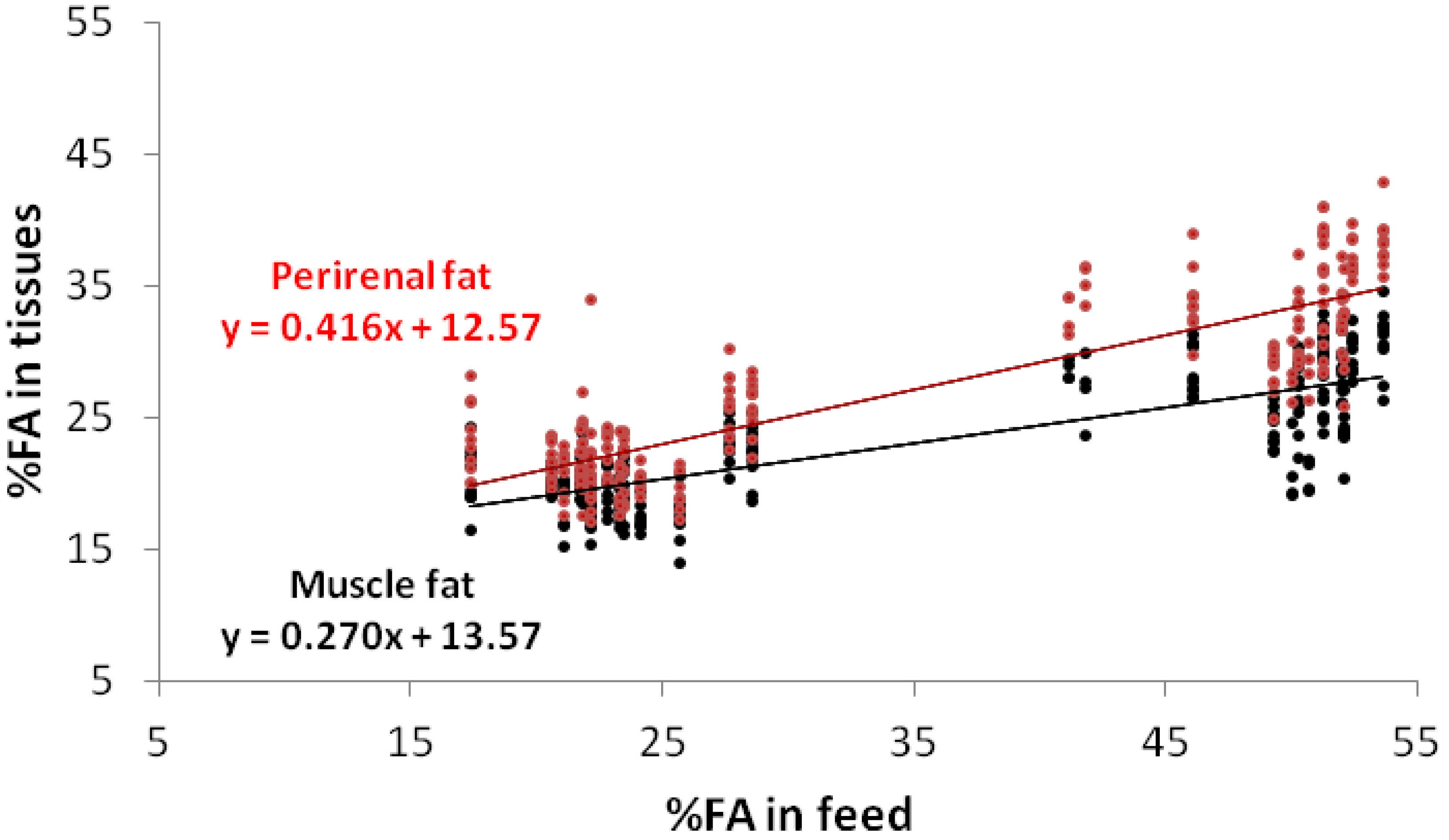{kind=link}
| Reference | Dietsn ∑ 27 | Raw material | Breed | Weight (g) | Animals n. ∑ 246 |
|---|---|---|---|---|---|
| [12] | 4 | Spirulina / fat level1 | New Zealand | 2,805 | 16 |
| [13] | 3 | False flax2 | Crossbred | 2,316 | 30 |
| [14] | 3 | Chia3 | Crossbred | 1,433 | 30 |
| [15] | 3 | Golden flaxseed4 | Crossbred | 2,074 | 30 |
| [16] | 4 | Spirulina5 | Crossbred | 2,034 | 40 |
| [17] | 3 | Perilla6 | Crossbred | 1,120 | 30 |
| [18] | 4 | Curcuma / oils7 | Crossbred | 1,512 | 40 |
| [19] | 3 | Tomato pomace8 | Crossbred | 1,166 | 30 |
| FA | Diets | Muscle | Perirenal | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | VC | Mean | SD | VC | Mean | SD | VC | |
| C10:0 | 0 | 0 | - | 0 | 0 | - | 0.066 | 0.11 | 165% |
| C12:0 | 0.004 | 0.019 | 539% | 0 | 0 | - | 0.12 | 0.13 | 110% |
| C14:0 | 0.43 | 0.66 | 155% | 2.07 | 0.42 | 20% | 1.87 | 0.43 | 23% |
| C14:1 | 0 | 0 | - | 0.063 | 0.12 | 194% | 0.042 | 0.08 | 183% |
| C15:0 | 0 | 0 | - | 0.36 | 0.19 | 53% | 0.48 | 0.10 | 20% |
| C16:0 | 15.81 | 7.16 | 45% | 25.78 | 3.61 | 14% | 23.39 | 4.34 | 19% |
| C16:1 n-7 | 0.040 | 0.085 | 211% | 0.39 | 1.16 | 295% | 0.68 | 1.26 | 185% |
| C16:1 n-9 | 0.24 | 0.25 | 104% | 2.88 | 1.66 | 57% | 1.70 | 1.21 | 71% |
| C16:3 n-4 | 0 | 0 | - | 0.04 | 0.12 | 280% | 0 | 0 | - |
| C17:0 | 0.56 | 1.00 | 180% | 0.44 | 0.25 | 57% | 0.42 | 0.25 | 60% |
| C17:1 | 0.008 | 0.032 | 381% | 0.052 | 0.12 | 224% | 0.10 | 0.15 | 154% |
| C18:0 | 3.43 | 1.48 | 43% | 5.94 | 0.92 | 15% | 6.48 | 2.32 | 36% |
| C18:1 n-7 | 0.45 | 0.42 | 93% | 0.78 | 0.55 | 70% | 0.62 | 0.49 | 79% |
| C18:1 n-9 | 22.81 | 7.25 | 32% | 24.71 | 3.88 | 16% | 23.85 | 5.56 | 23% |
| C18:2 n-6 | 36.43 | 13.66 | 37% | 23.21 | 4.76 | 20% | 27.00 | 6.57 | 24% |
| C18:3 n-3 | 13.15 | 13.67 | 104% | 7.79 | 7.52 | 96% | 9.98 | 10.39 | 104% |
| C18:3 n-6 | 0.10 | 0.33 | 341% | 0.11 | 0.23 | 215% | 0.14 | 0.28 | 205% |
| C18:4 n-3 | 1.97 | 5.37 | 273% | 0 | 0 | - | 0 | 0 | - |
| C20:0 | 0.25 | 0.23 | 91% | 0 | 0 | - | 0.62 | 1.35 | 216% |
| C20:1 n-9 | 0.80 | 2.12 | 264% | 0.33 | 1.03 | 317% | 0.57 | 1.42 | 248% |
| C20:2 n-6 | 0.46 | 1.64 | 354% | 0.060 | 0.19 | 315% | 0.19 | 0.21 | 115% |
| C20:3 n-3 | 0 | 0 | - | 0.029 | 0.08 | 287% | 0.025 | 0.06 | 248% |
| C20:3 n-6 | 0 | 0 | - | 0.24 | 0.72 | 294% | 0 | 0 | - |
| C20:4 n-6 | 0.052 | 0.12 | 230% | 2.04 | 1.20 | 59% | 0.074 | 0.17 | 228% |
| C20:5 n-3 | 0 | 0 | - | 0.023 | 0.08 | 341% | 0 | 0 | - |
| C21:0 | 0.21 | 0.63 | 302% | 0.15 | 0.69 | 458% | 0.034 | 0.07 | 191% |
| C22:1 n-9 | 0.18 | 0.66 | 375% | 0 | 0 | - | 0 | 0 | - |
| C22:4 n-6 | 0 | 0 | - | 0.15 | 0.28 | 185% | 0 | 0 | - |
| C22:6 n-3 | 0 | 0 | - | 0.084 | 0.24 | 286% | 0 | 0 | - |
| Others | 2.64 | 3.19 | 121% | 2.26 | 1.87 | 83% | 1.60 | 0.99 | 62% |
| Raw avg. | 207% | 173% | 123% |
3. Results and Discussion
| FA | R2 | MSE | M | P | Sign. | bM | Sign. | bP | Sign. | b1#b2 |
|---|---|---|---|---|---|---|---|---|---|---|
| C12:0 | 0.29 | 0.10 | 0.00 | 0.12 | <.0001 | 0.00 | 1 | 0.33 | 0.2289 | 0.4163 |
| C14:0 | 0.13 | 0.41 | 2.07 | 1.87 | <.0001 | 0.17 | <.0001 | 0.17 | <.0001 | 0.9727 |
| C16:0 | 0.44 | 3.13 | 25.80 | 23.37 | <.0001 | 0.27 | <.0001 | 0.37 | <.0001 | 0.0071 |
| C16:1 n-7 | 0.59 | 0.79 | 0.38 | 0.70 | 0.042 | 8.83 | <.0001 | 12.41 | <.0001 | 0.0000 |
| C16:1 n-9 | 0.15 | 1.44 | 2.88 | 1.70 | <.0001 | -0.15 | 0.733 | −1.08 | 0.0123 | 0.0000 |
| C17:0 | 0.09 | 0.24 | 0.45 | 0.42 | 0.48 | 0.08 | <.0001 | 0.06 | 0.0001 | 0.2995 |
| C18:0 | 0.04 | 1.77 | 5.93 | 6.48 | 0.0001 | 0.22 | 0.010 | −0.12 | 0.1513 | 0.0046 |
| C18:1 n-7 | 0.88 | 2.61 | 0.76 | 0.64 | 0.22 | 1.30 | <.0001 | 1.15 | <.0001 | 0.0000 |
| C18:1 n-9 | 0.71 | 0.18 | 24.65 | 23.94 | <.0001 | 0.39 | <.0001 | 0.66 | <.0001 | 0.0000 |
| C18:2 n-6 | 0.74 | 3.09 | 23.22 | 26.99 | 0.60 | 0.29 | <.0001 | 0.41 | <.0001 | 0.0000 |
| C18:3 n-3 | 0.91 | 2.74 | 7.85 | 9.91 | 0.0083 | 0.50 | <.0001 | 0.70 | <.0001 | 0.0000 |
| C18:3 n-6 | 0.84 | 0.10 | 0.10 | 0.14 | 0.028 | 0.61 | <.0001 | 0.78 | <.0001 | 0.0000 |
| C20:0 | 0.27 | 0.87 | 0.00 | 0.62 | <.0001 | 0.00 | 1 | −2.54 | <.0001 | <.0001 |
| C20:1 n-9 | 0.98 | 0.16 | 0.33 | 0.57 | <.0001 | 0.47 | <.0001 | 0.65 | <.0001 | 0.0000 |
| C20:2 n-6 | 0.13 | 0.20 | 0.06 | 0.19 | <.0001 | -0.01 | 0.1466 | −0.03 | <.0001 | 0.0295 |
| C20:4 n-6 | 0.67 | 0.76 | 2.03 | 0.00 | <.0001 | 4.04 | <.0001 | 0.00 | 1 | 0.0000 |




| Independent FAs (X) | Muscle (M) | Perirenal (P) | |||||
|---|---|---|---|---|---|---|---|
| R2 | 0.94 | 0.96 | |||||
| SE | 1.86 | 2.06 | |||||
| Mean | 7.96 | 9.98 | |||||
| Intercept | aM | SaM | Sign. | aP | SaP | Sign. | |
| −27 | 10.3 | 0.0136 | −46 | 9.7 | <.0001 | ||
| Means of X | bM | SbM | Sign. | bP | SbP | Sign. | |
| C12:0 | 0.004 | −10.2 | 8.5 | 0.18 | −18.6 | 8.7 | 0.035 |
| C14:0 | 0.43 | 4.5 | 1.5 | 0.0002 | 7.6 | 1.5 | <.0001 |
| C16:0 | 15.81 | 0.2 | 0.1 | 0.25 | 0.4 | 0.1 | <.0001 |
| C16:1 n-7 | 0.040 | 7.4 | 2.4 | 0.0003 | 7.5 | 2.3 | 0.0020 |
| C16:1 n-9 | 0.24 | −4.3 | 1.3 | 0.0004 | −5.3 | 1.3 | <.0001 |
| C18:1 n-9 | 22.81 | 0.4 | 0.1 | 0.0001 | 0.5 | 0.1 | <.0001 |
| C18:2 n-6 | 36.43 | 0.3 | 0.1 | 0.047 | 0.4 | 0.1 | <.0001 |
| C18:3 n-3 | 13.15 | 0.7 | 0.1 | <.0001 | 1.0 | 0.096 | <.0001 |
| C20:0 | 0.25 | 4.4 | 1.4 | 0.042 | 14.0 | 1.5 | <.0001 |
| C20:1 n-9 | 0.80 | 0.8 | 0.2 | <.0001 | 0.3 | 0.2 | 0.21 |
| C20:2 n-6 | 0.46 | 1.7 | 0.2 | <.0001 | 2.1 | 0.2 | <.0001 |
| r | r2 | |||||
|---|---|---|---|---|---|---|
| Diets | Muscle | Perirenal | Diets | Muscle | Perirenal | |
| Diets | 1 | 0.870 | 0.926 | 1 | 0.756 | 0.856 |
| Muscle | 0.870 | 1 | 0.975 | 0.756 | 1 | 0.951 |
| Perirenal | 0.926 | 0.975 | 1 | 0.857 | 0.951 | 1 |
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Dalle Zotte, A. Perception of rabbit meat quality and major factors influencing the rabbit carcass and meat quality. Livest. Prod. Sci. 2002, 75, 11–32. [Google Scholar] [CrossRef]
- Hernández, P. Enhancement of nutritional quality and safety in rabbit meat. In Proceeding of 9th World Rabbit Congress, Verona, Italy, 10–13 June 2008.
- Oliver, M.A.; Guerrero, L.; Diaz, I.; Gispert, M.; Pla, M.; Blasco, A. The effect of fat-enriched diets on the perirenal fat quality and sensory characteristics of meat from rabbits. Meat Sci. 1997, 47, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.A.; Díaz, I.; Pla, M.; Gil, M.; Blasco, A.; Oliver, M.À. Fatty acid composition of leg meat and perirenal fat of rabbits selected by growth rate. Food Chem. 2005, 90, 251–256. [Google Scholar] [CrossRef]
- Polak, T.; Gašperlin, L.; Rajar, A.; Žlender, B. Influence of genotype lines, age at slaughter and sexes on the composition of rabbit meat. Food Technol. Biotechnol. 2006, 44, 65–73. [Google Scholar]
- Hernández, P.; Pla, M.; Oliver, M.A.; Blasco, A. Relationships between meat quality measurements in rabbits fed with three diets with different fat type and content. Meat Sci. 2000, 55, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Szabó, A.; Husvéth, F.; Szendrõ, Z.; Repa, I.; Romvári, R. Effects of transcutaneous electrical nerve stimulation on the fatty acid profile of rabbit longissimus dorsi muscle (preliminary report). J. Anim. Physiol. Anim. Nutr. 2003, 87, 309–314. [Google Scholar] [CrossRef]
- Cavani, C.; Petracci, M.; Trocino, A.; Xiccato, G. Advances in research on poultry and rabbit meat quality. Ital. J. Anim. Sci. 2009, 8, 741–750. [Google Scholar]
- Szabó, A.; Fébel, H.; Dalle Zotte, A.; Mézes, M.; Szendrő, Zs.; Romvári, R. Reversibility of the changes of rabbit muscle fatty acid profile. Ital. J. Food Sci. 2004, 16, 69–77. [Google Scholar]
- Xiccato, G. Feeding and meat quality in rabbits: A review. World Rabbit Sci. 1999, 7, 75–86. [Google Scholar]
- Dalle Zotte, A.; Szendrő, Z. The role of rabbit meat as functional food. Meat Sci. 2011, 88, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Meineri, G.; Ingravalle, F.; Radice, E.; Aragno, M.; Peiretti, P.G. Effects of high fat diets and Spirulina platensis supplementation in New Zealand White Rabbits. J. Anim. Vet. Adv. 2009, 8, 2735–2744. [Google Scholar]
- Peiretti, P.G.; Mussa, P.P.; Prola, L.; Meineri, G. Use of different levels of false flax (Camelina sativa L.) seed in diets for fattening rabbits. Livest. Sci. 2007, 107, 192–198. [Google Scholar] [CrossRef]
- Peiretti, P.G.; Meineri, G. Effects on growth performance, carcass characteristics, and the fat and meat fatty acid profile of rabbits fed diets with chia (Salvia hispanica L.) seed supplements. Meat Sci. 2008, 80, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Peiretti, P.G.; Meineri, G. Effects of diets with increasing levels of golden flaxseed on carcass characteristics, meat quality and lipid traits of growing rabbits. Ital. J. Anim. Sci. 2010, 9, 372–377. [Google Scholar]
- Peiretti, P.G.; Meineri, G. Effects of Spirulina platensis supplementation on carcass characteristics, meat quality and lipid traits of growing rabbits. Livest. Sci. 2011, 140, 218–224. [Google Scholar] [CrossRef]
- Peiretti, P.G.; Gasco, L.; Brugiapaglia, A.; Gai, F. Effects of perilla (Perilla frutescens L.) seeds supplementation on performance, carcass characteristics, meat quality and fatty acid composition of rabbits. Livest. Sci. 2011, 138, 118–124. [Google Scholar] [CrossRef]
- Peiretti, P.G.; Masoero, G.; Meineri, G. Effects of replacing palm oil with maize oil and Curcuma longa supplementation on the performance, carcass characteristics, meat quality, and fatty acid profile of the perirenal fat and muscle of growing rabbits. Animal 2011, 5, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Peiretti, P.G.; Gai, F.; Rotolo, L.; Gasco, L. Effects of diets with increasing levels of dried tomato pomace on the performances and apparent digestibility of growing rabbits. Asian J. Anim. Vet. Adv. 2012, 7, 521–527. [Google Scholar] [CrossRef]
- Fernández-Carmona, J.; Blas, E.; Pascual, J.J.; Maertens, L.; Gidenne, T.; Xiccato, G.; García, J. Recommendations and guidelines for applied nutrition experiments in rabbits. World Rabbit Science 2005, 13, 209–228. [Google Scholar]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [PubMed]
- Chouinard, P.Y.; Corneau, L.; Sæbø, A.; Bauman, D.E. Milk yield and composition during abomasal infusion of conjugated linoleic acids in dairy cows. J. Dairy Sci. 1999, 82, 2737–2745. [Google Scholar] [CrossRef] [PubMed]
- Nie, N.H.; Hull, C.H.; Jenkins, J.G.; Steinbrenner, K.; Bent, D.H. SPSS Statistical Package for the Social Sciences; McGraw-Hill: New York, NY, USA, 1981. [Google Scholar]
- Bernardini, M.; Dal Bosco, A.; Castellini, C. Effect of dietary n-3/n-6 ratio on fatty acid composition of liver, meat and perirenal fat in rabbits. Anim. Sci. 1999, 68, 647–654. [Google Scholar]
- Combes, S. Valeur nutritionnelle de la viande de lapin. INRA Prod. Anim. 2004, 17, 373–383. [Google Scholar]
- Petracci, M.; Bianchi, M.; Cavani, C. Development of rabbit meat products fortified with n-3 polyunsaturated fatty acids. Nutrients 2009, 1, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ouhayoun, J.; Demarne, Y.; Delmas, D.; Lebas, F. Utilization de pellicules de colza dans l’alimentation du lapin en croissance. II. Effect sur la qualite des carcasses. Ann. Zootech. 1981, 30, 325–333. [Google Scholar] [CrossRef]
- Cobos, A.; Cambero, M.I.; Ordotiez, J.A.; de la Hoz, L. Effect of fat enriched diets on rabbit meat fatty acid composition. J. Sci. Food Agric. 1993, 62, 83–88. [Google Scholar] [CrossRef]
- Xiccato, G.; Parigi-Bini, R.; Dalle Zotte, A.; Carazzolo, A. Effect of age, sex and transportation on the composition and sensory properties of rabbit meat. In Proceedings 40th International Congress of Meat Science and Technology, The Hague, The Netherland, 28 August–3 September 1994; p. 52.
- Szabó, A.; Romvári, R.; Fébel, H.; Nagy, I.; Szendrõ, Zs. Fatty acid composition of two different muscles in rabbits: Alterations in response to saturated or unsaturated dietary fatty acid complementation. World Rabbit Sci. 2001, 9, 155–158. [Google Scholar]
- Ouhayoun, J.; Kopp, J.; Bonnet, M.; Demarne, Y.; Delmas, D. Influence de la composition des graisses alimentaires sur les proprietes des lipides perirenaux et la qualiteé de la viande de lapin. Sci. Alim. 1987, 7, 521–534. [Google Scholar]
- Hernández, P.; Cesari, V.; Pla, M. Effect of the dietary fat on fatty acid composition and oxidative stability of rabbit meat. In Proceeding of the 53rd International Congress of Meat Science and Technology, Beijing, China, 5–10 August 2007; pp. 367–370.
- Gigaud, V.; Combes, S. The effect of decreasing the omega-6/omega-3 ratio in feed on fatty acid content of rabbit meat to meet human dietary recommendations. In Proceeding of the 9th World Rabbit Congress, Verona, Italy, 10–13 June 2008; pp. 1353–1357.
- Colin, M.; Raguanes, N.; Le Berre, G.; Charrier, S.; Prigent, A.Y.; Perrin, G. Influence d’un enrichissement de l’aliment en acides gras oméga 3 provenant de graines de lin extrudées (Tradi-Lin®) sur le lipids et les caractéristiques hédoniques de la viande de lapin. In Proceeding of the 11èmes Journées de la Recherche Cunicole, Paris, France, 29–30 November 2005; pp. 163–166.
- Garcia, P.T. Metabolism of α-linolenic acid (ALA) in meat animals. Soybean and Nutrition; El-Shemy, H., Ed.; InTech: Rijeka, Croatia, 2011. Available online: http://www.intechopen.com/articles/show/title/metabolism-of-linolenic-acid-ala-in-meat-animals (accessed on 20 February 2012).
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peiretti, P.G. Effects of Dietary Fatty Acids on Lipid Traits in the Muscle and Perirenal Fat of Growing Rabbits Fed Mixed Diets. Animals 2012, 2, 55-67. https://doi.org/10.3390/ani2010055
Peiretti PG. Effects of Dietary Fatty Acids on Lipid Traits in the Muscle and Perirenal Fat of Growing Rabbits Fed Mixed Diets. Animals. 2012; 2(1):55-67. https://doi.org/10.3390/ani2010055
Chicago/Turabian StylePeiretti, Pier Giorgio. 2012. "Effects of Dietary Fatty Acids on Lipid Traits in the Muscle and Perirenal Fat of Growing Rabbits Fed Mixed Diets" Animals 2, no. 1: 55-67. https://doi.org/10.3390/ani2010055
APA StylePeiretti, P. G. (2012). Effects of Dietary Fatty Acids on Lipid Traits in the Muscle and Perirenal Fat of Growing Rabbits Fed Mixed Diets. Animals, 2(1), 55-67. https://doi.org/10.3390/ani2010055