Influence of Different Housing Systems on Distribution, Function and Mitogen-Response of Leukocytes in Pregnant Sows

Abstract
:Simple Summary
Abstract
1. Introduction
2. Experimental Section
2.1. Experimental Animals and Housing Conditions
2.2. Experimental Procedure
2.3. Blood Sampling and Treatment of Samples
2.4. Body Mass and Medical Treatments
2.5. Flow Cytometry
2.6. Separation of Peripheral Blood Mononuclear Cells (PBMCs)
2.6.1. Lymphocyte Proliferation
2.6.2. Intracellular Cytokine Staining
2.7. Cortisol Determinations
2.8. Statistical Analysis

3. Results
3.1. Effects of Housing and Gestational Stage on Number and Ratio of Blood Immune Cells


3.2. Effects of Housing and Gestational Stage on Lymphocyte Functionality

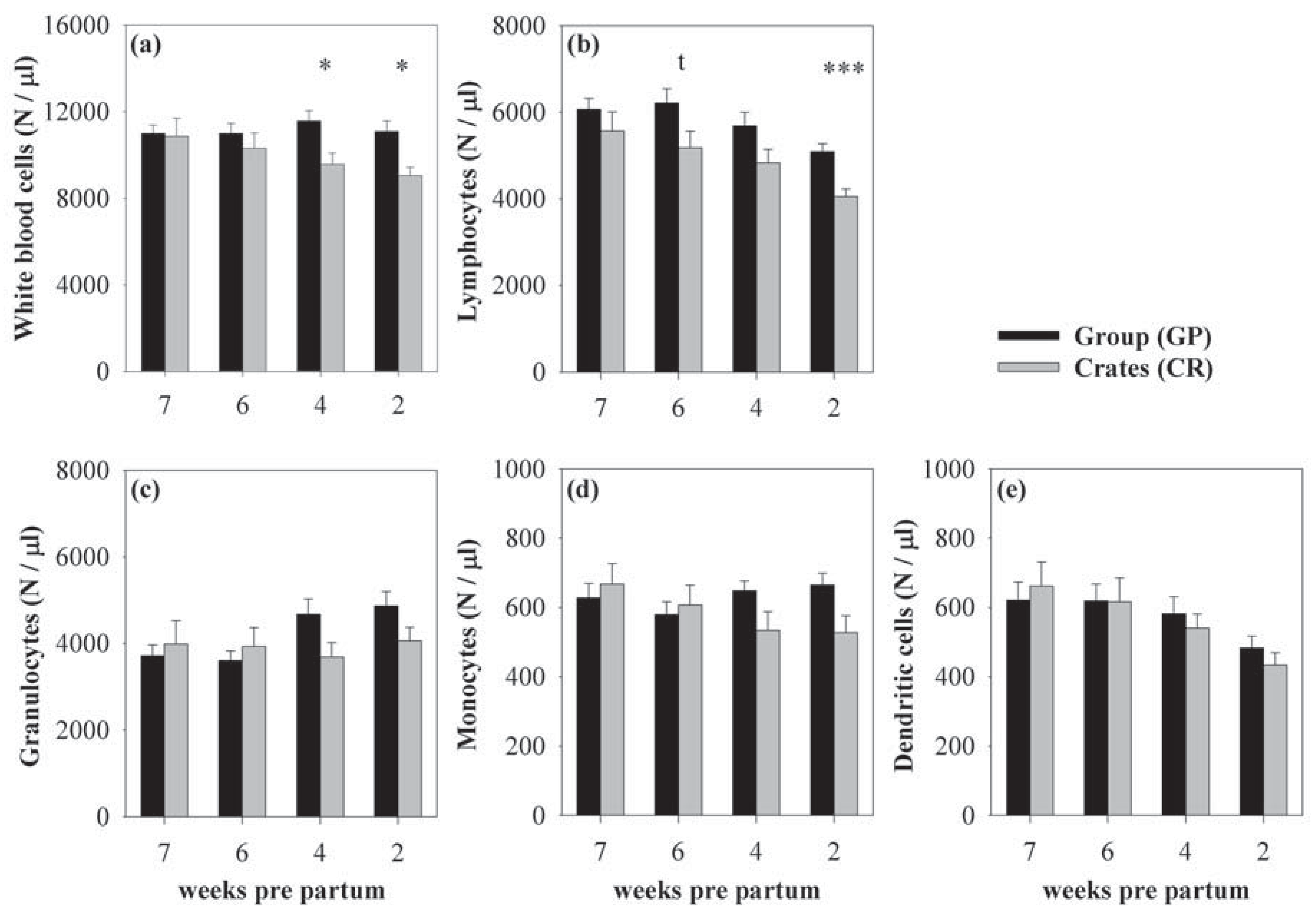{kind=link}
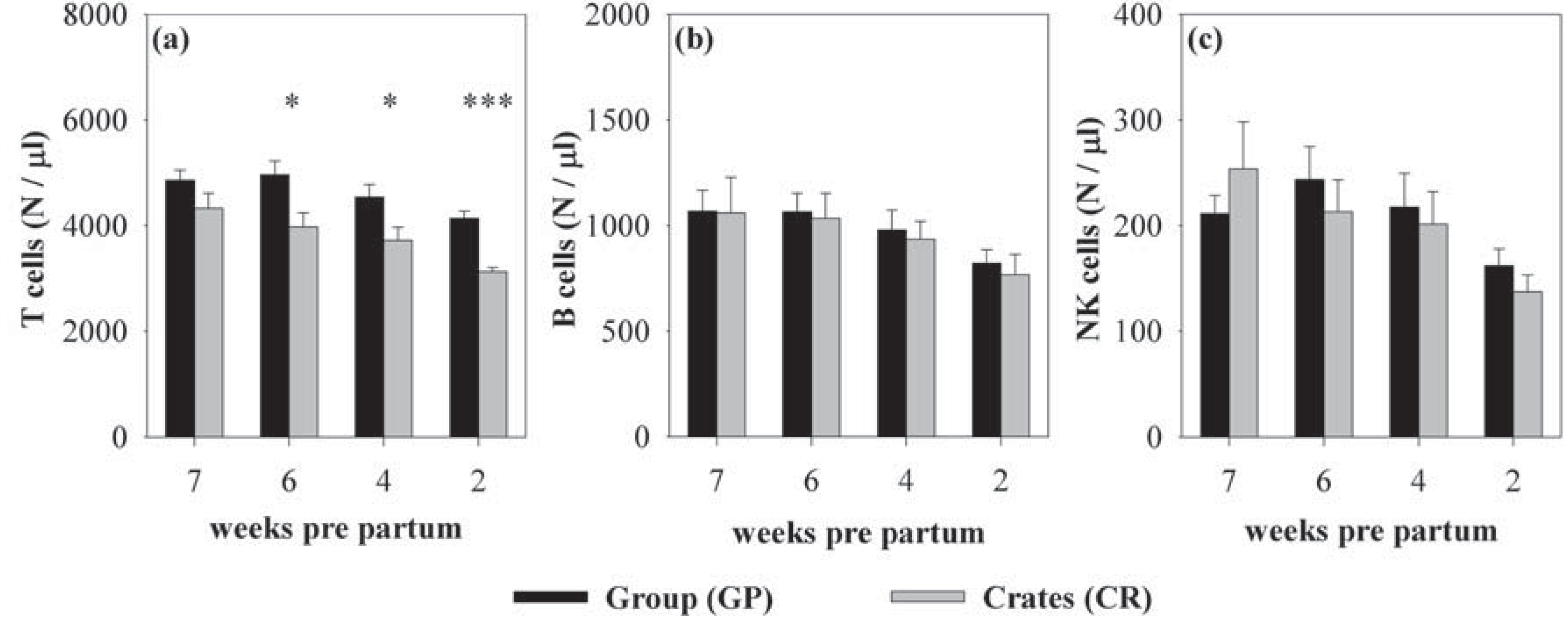{kind=link}
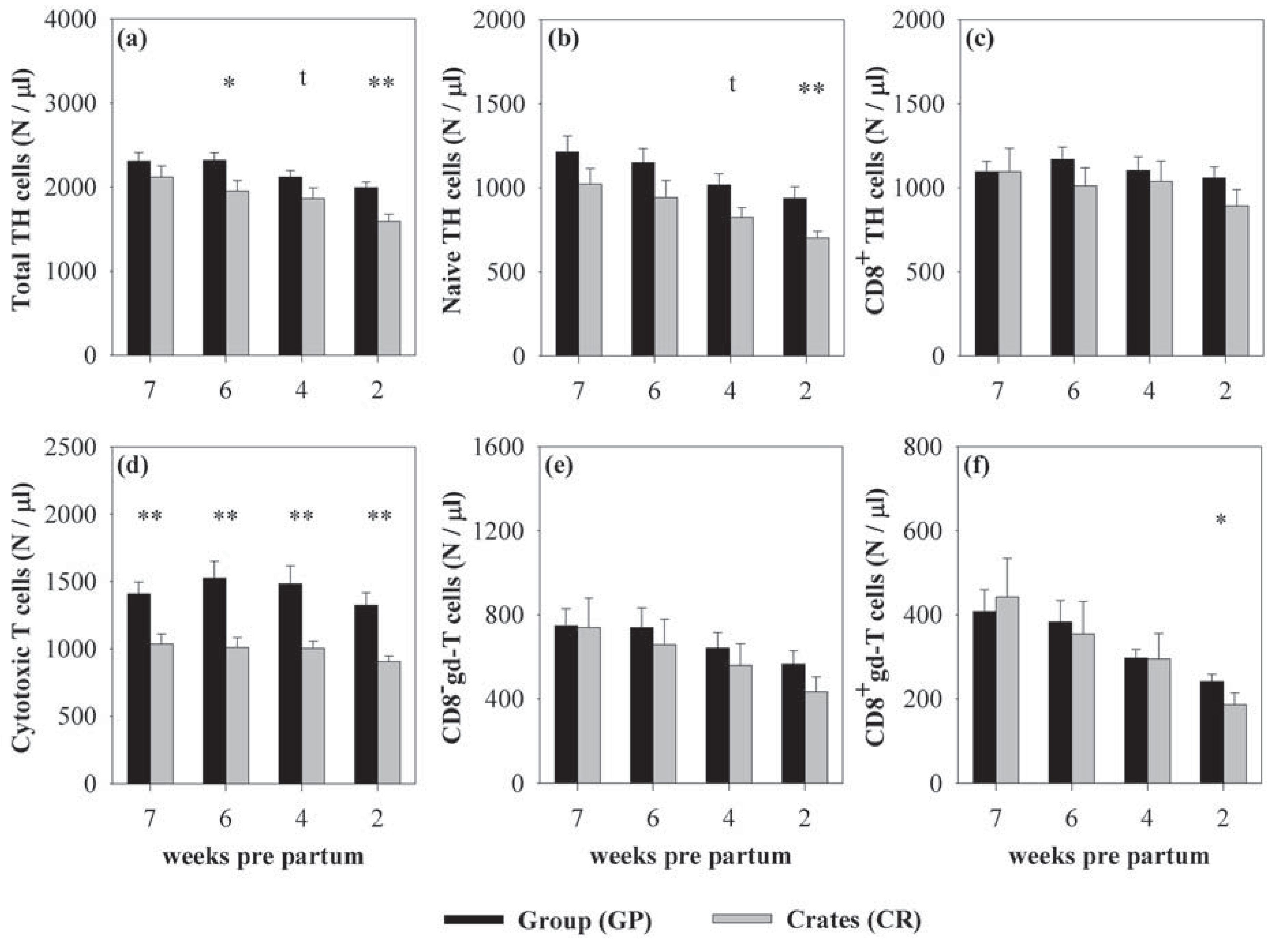{kind=link}
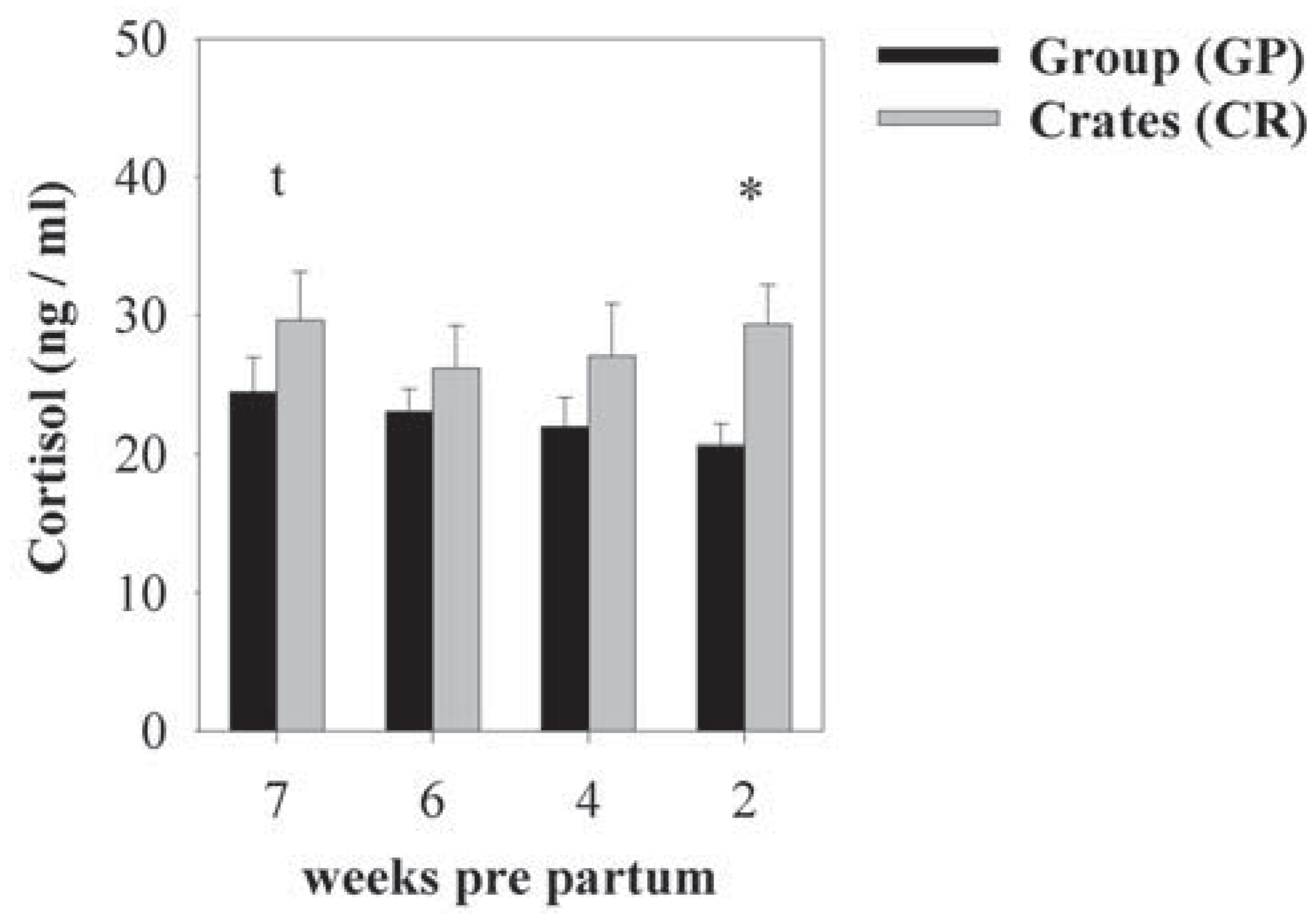{kind=link}
| Parameter | Weeks pre partum | p values | ||||||
|---|---|---|---|---|---|---|---|---|
| 7 | 6 | 4 | 2 | Housing | Gestational stage | Housing × gestational stage | ||
| Lymphocyte proliferation | ||||||||
| ∆ cpm ConA | GP | 50294 ± 2998 | 48318 ± 3584 | 38945 ± 2289 | 39431 ± 3486 | n.s. | n.s. | n.s. |
| CR | 48847 ± 3763 | 47777 ± 3926 | 41703 ± 3941 | 40191 ± 4459 | ||||
| ∆ cpm PWM | GP | 67368 ± 2530 | 69127 ± 2737 | 73542 ± 4213 | 70502 ± 2959 | n.s. | n.s. | n.s. |
| CR | 74993 ± 3982 | 73998 ± 4505 | 71465 ± 4086 | 71596 ± 3826 | ||||
| ∆% of cytokine-producing T cells among all T cells | ||||||||
| IFNγ+ | GP | 11.4 ± 2.6 | 8.6 ± 1.6 | 11.3 ± 1.7 | 12.2 ± 1.9 | n.s. | <0.1 | n.s. |
| CR | 12.1 ± 3.0 | 10.5 ± 2.5 | 14.1 ± 3.0 | 12.8 ± 3.5 | ||||
| TNFα+ | GP | 28.7 ± 5.8 | 27.0 ± 3.7 | 30.5 ± 2.7 | 32.8 ± 4.0 | n.s. | n.s. | n.s. |
| CR | 31.3 ± 7.6 | 29.2 ± 5.0 | 37.2 ± 7.4 | 28.1 ± 6.8 | ||||
| IFNγ+/TNFα+ | GP | 10.1 ± 2.4 | 7.9 ± 1.4 | 10.0 ± 1.5 | 10.8 ± 1.8 | n.s. | <0.05 | n.s. |
| CR | 10.3 ± 3.1 | 8.9 ± 2.3 | 12.2 ± 2.6 | 11.6 ± 3.4 | ||||
3.3. Effects of Housing and Gestational Stage on Plasma Cortisol Levels and Body Mass

4. Discussion
4.1. Housing-Associated Alterations of the Immune System
4.2. Housing-Associated Alterations of Plasma Cortisol
4.3. Factors Contributing to the Immunological Effects
4.4. Immunological Alterations During Pregnancy
5. Conclusions
Acknowledgments
Conflicts of Interest
References and Notes
- Turner, J. The Welfare of Europe’s Sows in Close Confinement Stalls: A Report Prepared for the European Coalition for Farm Animals; CIWF: Hampshire, UK, 2000. Available online: http://www.ciwf.org.uk/includes/documents/cm_docs/2008/w/welfare_of_europes_sows_in_close_confinement_stalls.pdf (accessed on 27 June 2011).
- Karlen, G.A.M.; Hemsworth, P.H.; Gonyou, H.W.; Fabrega, E.; Strom, D.A.; Smits, R.J. The welfare of gestating sows in conventional stalls and large groups on deep litter. Appl. Anim. Behav. Sci. 2007, 105, 87–101. [Google Scholar] [CrossRef]
- Chapinal, N.; Ruiz de la Torre, J.L.; Cerisuelo, A.; Gasa, J.; Baucells, M.D.; Coma, J.; Vidal, A.; Manteca, X. Evaluation of welfare and productivity in pregnant sows kept in stalls or in 2 different group housing systems. J. Vet. Behav. 2010, 5, 82–93. [Google Scholar] [CrossRef]
- Broom, D.M.; Mendl, M.T.; Zanella, A.J. A comparison of the welfare of sows in different housing conditions. Anim. Sc. 1995, 61, 369–385. [Google Scholar] [CrossRef]
- Barnett, J.L.; Hemsworth, P.H.; Winfield, C.G. The effects of design of individual stalls on the social behaviour and physiological responses related to the welfare of pregnant pigs. Appl. Anim. Behav. Sci. 1987, 18, 133–142. [Google Scholar] [CrossRef]
- Barnett, J.L.; Hemsworth, P.H.; Cronin, G.M.; Newman, E.A.; McCallum, T.H. Effects of design of individual cage-stalls on the behavioural and physiological responses related to the welfare of pregnant pigs. Appl. Anim. Behav. Sci. 1991, 32, 23–33. [Google Scholar] [CrossRef]
- The Welfare of Intensively Kept Pigs. Report of the Scientific Veterinary Committee. 1997. Available online: http://ec.europa.eu/food/fs/sc/oldcomm4/out17_en.pdf (accessed on 7 June 2013).
- Arey, D.S. Time course for the formation and disruption of social organisation in group-housed sows. Appl. Anim. Behav. Sci. 1999, 62, 199–207. [Google Scholar] [CrossRef]
- Couret, D.; Otten, W.; Puppe, B.; Prunier, A.; Merlot, E. Behavioural, endocrine and immune responses to repeated social stress in pregnant gilts. Animal 2009, 3, 118–127. [Google Scholar] [CrossRef]
- Krauss, V.; Hoy, S. Dry sows in dynamic groups: An investigation of social behaviour when introducing new sows. Appl. Anim. Behav. Sci. 2011, 130, 20–27. [Google Scholar] [CrossRef]
- Hayne, M.; Gonyou, H.W. Effects of regrouping on the individual behavioural characteristics of pigs. Appl. Anim. Behav. Sci. 2003, 8, 267–278. [Google Scholar] [CrossRef]
- Coutellier, L.; Arnould, C.; Boissy, A.; Orgeur, P.; Prunier, A.; Veissier, I.; Meunier-Salaün, M.C. Pig’s responses to repeated social regrouping and relocation during the growing-finishing period. Appl. Anim. Behav. Sci. 2007, 105, 102–114. [Google Scholar] [CrossRef]
- Ringgenberg, N.; Bergeron, R.; Meunier-Salaün, M.C.; Devillers, N. Impact of social stress during gestation and environmental enrichment during lactation on the maternal behavior of sows. Appl. Anim. Behav. Sci. 2012, 136, 126–135. [Google Scholar] [CrossRef]
- Deguchi, E.; Akuzawa, M. Effects of fighting after grouping on plasma cortisol concentration and lymphocyte blastogenesis of peripheral blood mononuclear cells induced by mitogens in piglets. J. Vet. Med. Sci. 1998, 60, 149–153. [Google Scholar] [CrossRef]
- Bauer, J. Untersuchungen zur Gruppenbildung von Sauen unter Verhaltens-, Gesundheits- und Leistungsaspekten. Inaugural Dissertation, Justus-Liebig University of Gießen, Gießen, Germany, 2005. [Google Scholar]
- Moore, A.S.; Gonyou, H.W.; Ghent, A.W. Integration of newly introduced and resident sows following grouping. Appl. Anim. Behav. Sci. 1993, 38, 257–267. [Google Scholar] [CrossRef]
- Stefanski, V.; Engler, H. Effects of acute and chronic social stress on blood cellular immunity in rats. Physiol. Behav. 1998, 64, 733–741. [Google Scholar]
- Stefanski, V.; Knopf, G.; Schulz, S. Long-term colony housing in long evans rats: Immunological, hormonal, and behavioral consequences. J. Neuroimmunol. 2001, 114, 122–130. [Google Scholar] [CrossRef]
- Bartolomucci, A. Social stress, immune functions and disease in rodents. Front. Neuroendocrinol. 2007, 28, 28–49. [Google Scholar] [CrossRef]
- Damgaard, B.M.; Studnitz, M.; Jensen, K.H. The effect of continuous grouping of pigs in large groups on stress response and haematological parameters. Livest. Sci. 2009, 121, 137–140. [Google Scholar] [CrossRef]
- de Groot, J.; Ruis, M.A.W.; Scholten, J.W.; Koolhaas, J.M.; Boersma, W.J.A. Long-term effects of social stress on antiviral immunity in pigs. Physiol. Behav. 2001, 73, 145–158. [Google Scholar] [CrossRef]
- Weng, R.C.; Edwards, S.A.; Hsia, L.C. Effect of individual, group or ESF housing in pregnancy and individual or group housing in lactation on sow behavior. Asian-Aust. J. Anim. Sc. 2009, 22, 1574–1580. [Google Scholar]
- Kongsted, A.G. Stress and fear as possible mediators of reproduction problems in group housed sows: A review. Acta Agric. Scand. A Anim. Sci. 2004, 54, 58–66. [Google Scholar]
- Rhodes, R.T.; Appleby, M.C.; Chinn, K.; Douglas, L.; Firkins, L.D.; Houpt, K.A.; Irwin, C.; McGlone, J.J.; Sundberg, P.; Tokach, L.; Wills, R.W. A comprehensive review of housing for pregnant sows. J. Am. Vet. Med. Assoc. 2005, 227, 1580–1590. [Google Scholar] [CrossRef]
- von Borell, E.; Morris, J.R.; Hurnik, J.F.; Mallard, B.A.; Buhr, M.M. The performance of gilts in a new group housing system: Endocrinological and immunological functions. J. Anim. Sci. 1992, 70, 2714–2721. [Google Scholar]
- Hulbert, L.E.; McGlone, J.J. Evaluation of drop versus trickle-feeding systems for crated or group-penned gestating sows. J. Anim. Sci. 2006, 84, 1004–1014. [Google Scholar]
- Sorrells, A.D.; Eicher, S.D.; Harris, M.J.; Pajor, E.A.; Richert, B.T. Periparturient cortisol, acute phase cytokine, and acute phase protein profiles of gilts housed in groups or stalls during gestation. J. Anim. Sci. 2007, 85, 1750–1757. [Google Scholar] [CrossRef]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef]
- Gerner, W.; Käser, T.; Saalmüller, A. Porcine T lymphocytes and NK cells—An update. Dev. Comp. Immunol. 2009, 33, 310–320. [Google Scholar] [CrossRef]
- Saalmüller, A.; Werner, T.; Fachinger, V. T-helper cells from naive to committed. Vet. Immunol. Immunopathol. 2002, 87, 137–145. [Google Scholar] [CrossRef]
- Charerntantanakul, W.; Roth, J.A. Biology of porcine T lymphocytes. Anim. Health Res. Rev. 2007, 7, 81–96. [Google Scholar] [CrossRef]
- Haas, W. Gamma/Delta Cells. Annu. Rev. Immunol. 1993, 11, 637–685. [Google Scholar] [CrossRef]
- Holderness, J.; Hedges, J.F.; Ramstead, A.; Jutila, M.A. Comparative biology of γδ T cell function in humans, mice, and domestic animals. Annu. Rev. Anim. Biosci. 2013, 1, 99–124. [Google Scholar] [CrossRef]
- Hayday, A.C. γδ Cells: A right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 2000, 18, 975–1026. [Google Scholar] [CrossRef]
- Stefanski, V.; Raabe, C.; Schulte, M. Pregnancy and social stress in female rats: Influences on blood leukocytes and corticosterone concentrations. J. Neuroimmunol. 2005, 162, 81–88. [Google Scholar] [CrossRef]
- Zelnickova, P.; Faldyna, M.; Stepanova, H.; Ondracek, J.; Kovaru, F. Intracellular cytokine detection by flow cytometry in pigs: Fixation, permeabilization and cell surface staining. J. Immunol. Methods 2007, 327, 18–29. [Google Scholar] [CrossRef]
- Mascher, B.; Schlenke, P.; Seyfarth, M. Expression and kinetics of cytokines determined by intracellular staining using flow cytometry. J. Immunol. Methods 1999, 223, 115–121. [Google Scholar] [CrossRef]
- Claus, R.; Weiler, U. Relationships between IGF-I, cortisol, and osteocalcin in peripheral plasma of growing pigs. Exp. Clin. Endocrinol. Diabetes 1996, 104, 344–349. [Google Scholar] [CrossRef]
- Grün, V.; Schmucker, S.; Schalk, C.; Flauger, B.; Weiler, U.; Stefanski, V.; University of Hohenheim, Stuttgart, Germany. Unpublished data. 2012.
- Tarcic, N.; Ovadia, H.; Weiss, D.W.; Weidenfeld, J. Restraint stress-induced thymic involution and cell apoptosis are dependent on endogenous glucocorticoids. J. Neuroimmunol. 1998, 82, 40–46. [Google Scholar] [CrossRef]
- Engler, H.; Stefanski, V. Social stress and T cell maturation in male rats: Transient and persistent alterations in thymic function. Psychoneuroendocrinology 2003, 28, 951–969. [Google Scholar] [CrossRef]
- Stefanski, V.; Peschel, A.; Reber, S. Social stress affects migration of blood T cells into lymphoid organs. J. Neuroimmunol. 2003, 138, 17–24. [Google Scholar] [CrossRef]
- Leandro, C.G.; Martins de Lima, T.; Folador, A.; Alba-Loreiro, T.; do Nascimento, E.; Manhães de Castro, R.; de Castro, C.M.; Pithon-Curi, T.; Curi, R. Physical training attenuates the stress-induced changes in rat T-lymphocyte function. Neuroimmunomodulation 2006, 13, 105–113. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Stress-induced augmentation of immune function—The role of stress hormones, leukocyte trafficking, and cytokines. Brain Behav. Immun. 2002, 16, 785–798. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Enhancing versus suppressive effects of stress on immune function: Implications for immunoprotection versus immunopathology. Allergy Asthma Clin. Immunol. 2008, 4, 2–11. [Google Scholar] [CrossRef]
- von Holst, D. The concept of stress and its relevance for animal behavior. Adv. Study Behav. 1998, 27, 1–131. [Google Scholar]
- Geverink, N.A.; Schouten, W.G.P.; Gort, G.; Wiegant, V.M. Individual differences in behaviour, physiology and pathology in breeding gilts housed in groups or stalls. Appl. Anim. Behav. Sci. 2003, 81, 29–41. [Google Scholar] [CrossRef]
- Dhabhar, F.S.; Miller, A.H.; McEwen, B.S.; Spencer, R.L. Effects of stress on immune cell distribution. Dynamics and hormonal mechanisms. J. Immunol. 1995, 154, 5511–5527. [Google Scholar]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating, permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef]
- Webster Marketon, J.I.; Glaser, R. Stress hormones and immune function. Cell. Immunol. 2008, 252, 16–26. [Google Scholar]
- Kanitz, E.; Tuchscherer, M.; Puppe, B.; Tuchscherer, A.; Stabenow, B. Consequences of repeated early isolation in domestic piglets (sus scrofa) on their behavioural, neuroendocrine, and immunological responses. Brain Behav. Immun. 2004, 18, 35–45. [Google Scholar] [CrossRef]
- Salak-Johnson, J.L.; McGlone, J.J. Making sense of apparently conflicting data: Stress and immunity in swine and cattle. J. Anim. Sci. 2007, 85, E81–E88. [Google Scholar] [CrossRef]
- Elenkov, I.J. Glucocorticoids and the Th1/Th2 balance. Ann. N. Y. Acad. Sci. 2004, 1024, 138–146. [Google Scholar] [CrossRef]
- Wegmann, T.G.; Lin, H.; Guilbert, L.; Mosmann, T.R. Bidirectional cytokine interactions in the maternal-fetal relationship: Is successful pregnancy a TH2 phenomenon? Immunol. Today 1993, 14, 353–356. [Google Scholar]
- Szerkeres-Bartho, J. Immunological relationship between the mother and the fetus. Int. Rev. Immunol. 2002, 21, 471–495. [Google Scholar] [CrossRef]
- Nakamura, H.; Seto, T.; Nagase, H.; Yoshida, M.; Dan, S.; Ogino, K. Inhibitory effect of pregnancy on stress-induced immunosuppression through corticotropin releasing hormone (CRH) and dopaminergic systems. J. Neuroimmunol. 1997, 75, 1–8. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar]
- Pedersen, B.K.; Toft, A.D. Effects of exercise on lymphocytes and cytokines. Br. J. Sports Med. 2000, 34, 246–251. [Google Scholar] [CrossRef]
- Papworth, T.A.; Clubb, S.K. Clinical pathology in the female rat during the pre- and postnatal period. Comp. Haematol. Int. 1995, 5, 13–24. [Google Scholar] [CrossRef]
- Watanabe, M.; Iwatani, Y.; Kaneda, T.; Hidaka, Y.; Mitsuda, N.; Morimoto, Y.; Amino, N. Changes in T, B, and NK lymphocyte subsets during and after normal pregnancy. Am. J. Reprod. Immunol. 1997, 37, 368–377. [Google Scholar] [CrossRef]
- Kühnert, M.; Strohmeier, R.; Stegmüller, M.; Halberstadt, E. Changes in lymphocyte subsets during normal pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 1998, 76, 147–151. [Google Scholar] [CrossRef]
- Luppi, P. How immune mechanisms are affected by pregnancy. Vaccine 2003, 21, 3352–3357. [Google Scholar] [CrossRef]
- Knight, J.W.; Kukoly, C.A. In vitro release of progesterone and estrone by the porcine placenta throughout gestation. Domest. Anim. Endocrinol. 1990, 7, 497–508. [Google Scholar] [CrossRef]
- Knight, J.W. Aspects of placental estrogen synthesis in the pig. Exp. Clin. Endocrinol. 1994, 102, 175–184. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Grün, V.; Schmucker, S.; Schalk, C.; Flauger, B.; Weiler, U.; Stefanski, V. Influence of Different Housing Systems on Distribution, Function and Mitogen-Response of Leukocytes in Pregnant Sows. Animals 2013, 3, 1123-1141. https://doi.org/10.3390/ani3041123
Grün V, Schmucker S, Schalk C, Flauger B, Weiler U, Stefanski V. Influence of Different Housing Systems on Distribution, Function and Mitogen-Response of Leukocytes in Pregnant Sows. Animals. 2013; 3(4):1123-1141. https://doi.org/10.3390/ani3041123
Chicago/Turabian StyleGrün, Verena, Sonja Schmucker, Christiane Schalk, Birgit Flauger, Ulrike Weiler, and Volker Stefanski. 2013. "Influence of Different Housing Systems on Distribution, Function and Mitogen-Response of Leukocytes in Pregnant Sows" Animals 3, no. 4: 1123-1141. https://doi.org/10.3390/ani3041123
APA StyleGrün, V., Schmucker, S., Schalk, C., Flauger, B., Weiler, U., & Stefanski, V. (2013). Influence of Different Housing Systems on Distribution, Function and Mitogen-Response of Leukocytes in Pregnant Sows. Animals, 3(4), 1123-1141. https://doi.org/10.3390/ani3041123