
Behavioral Response of Invertebrates to Experimental Simulation of Pre-Seismic Chemical Changes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction

2. Experimental Section
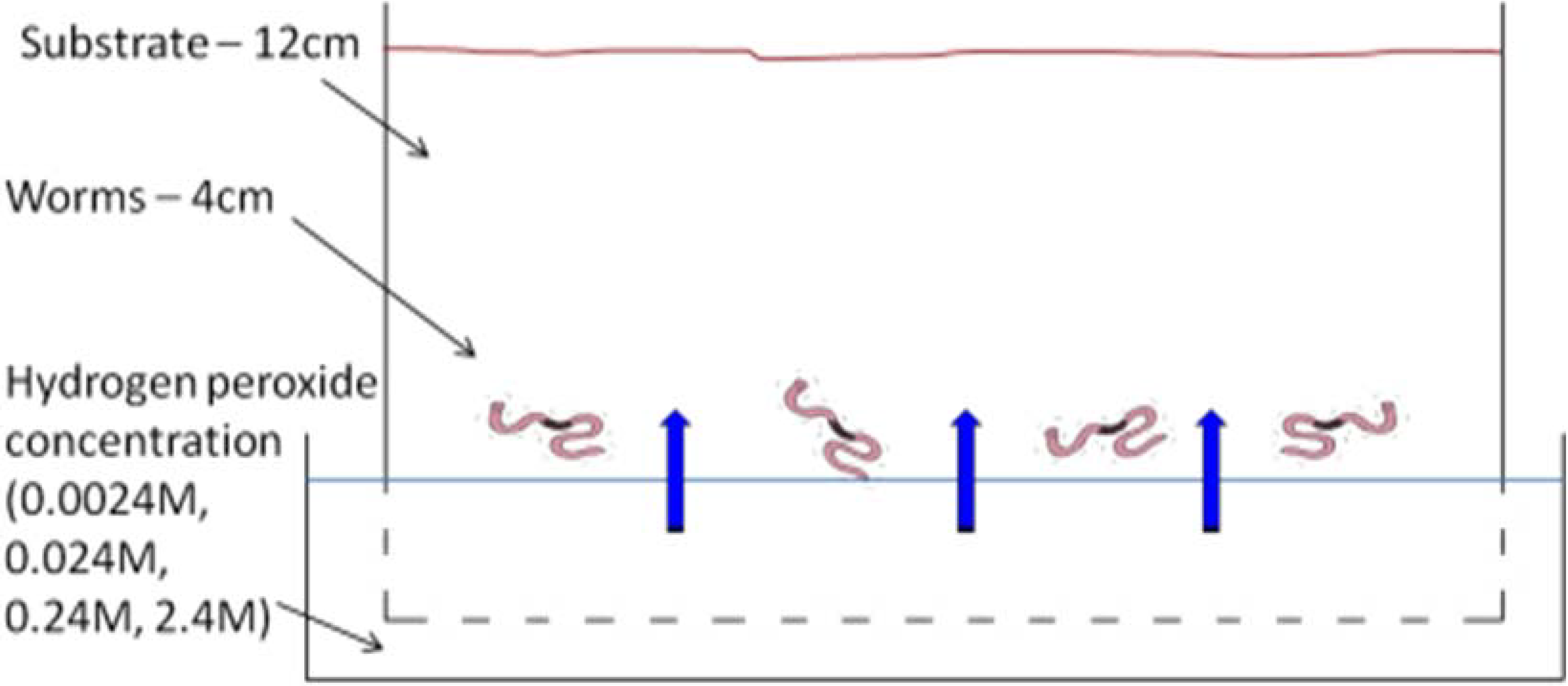
2.1. Earthworm Perfusion Test

2.2. Earthworm Pre-Mixed Soil Test
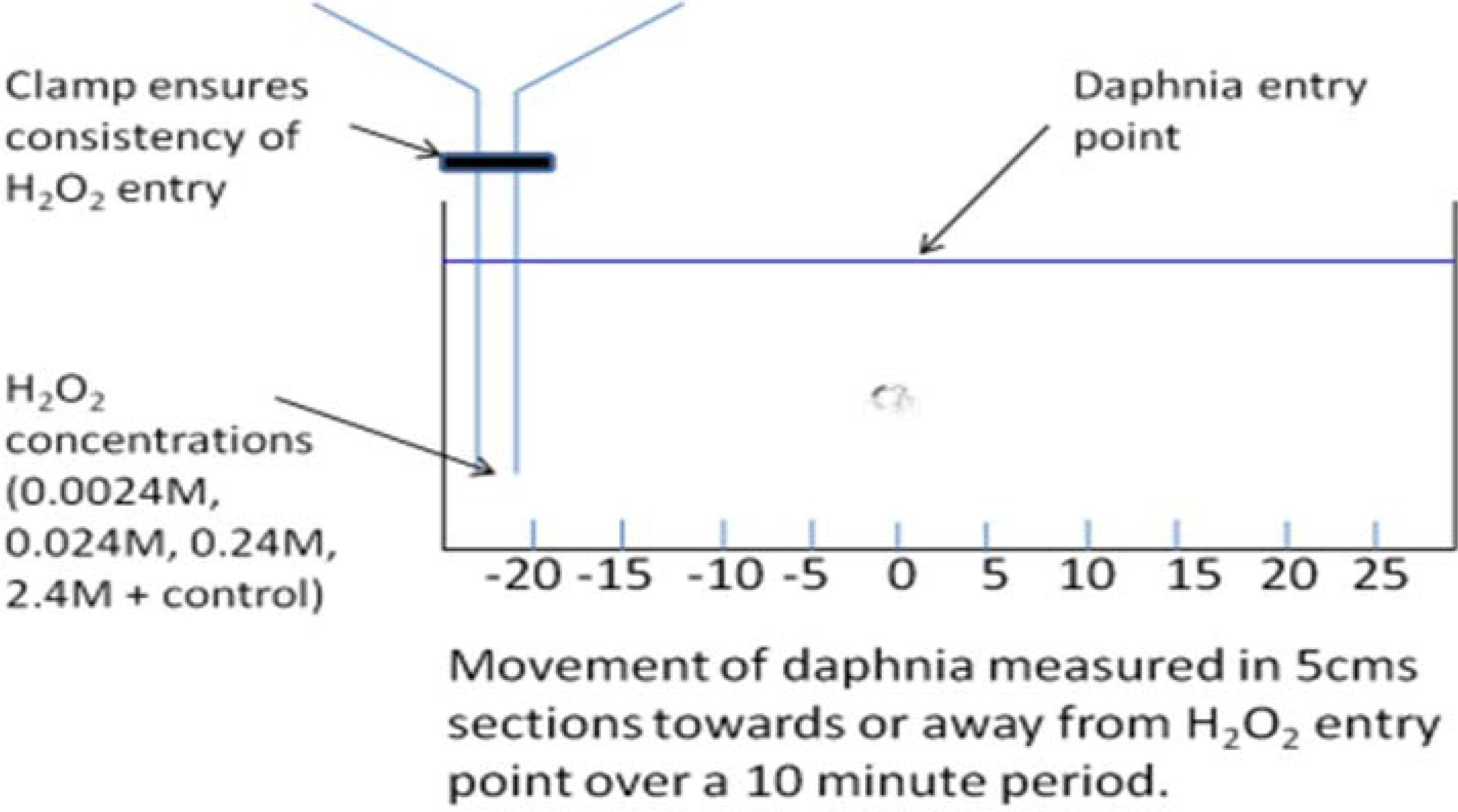
2.3. Daphnia Test


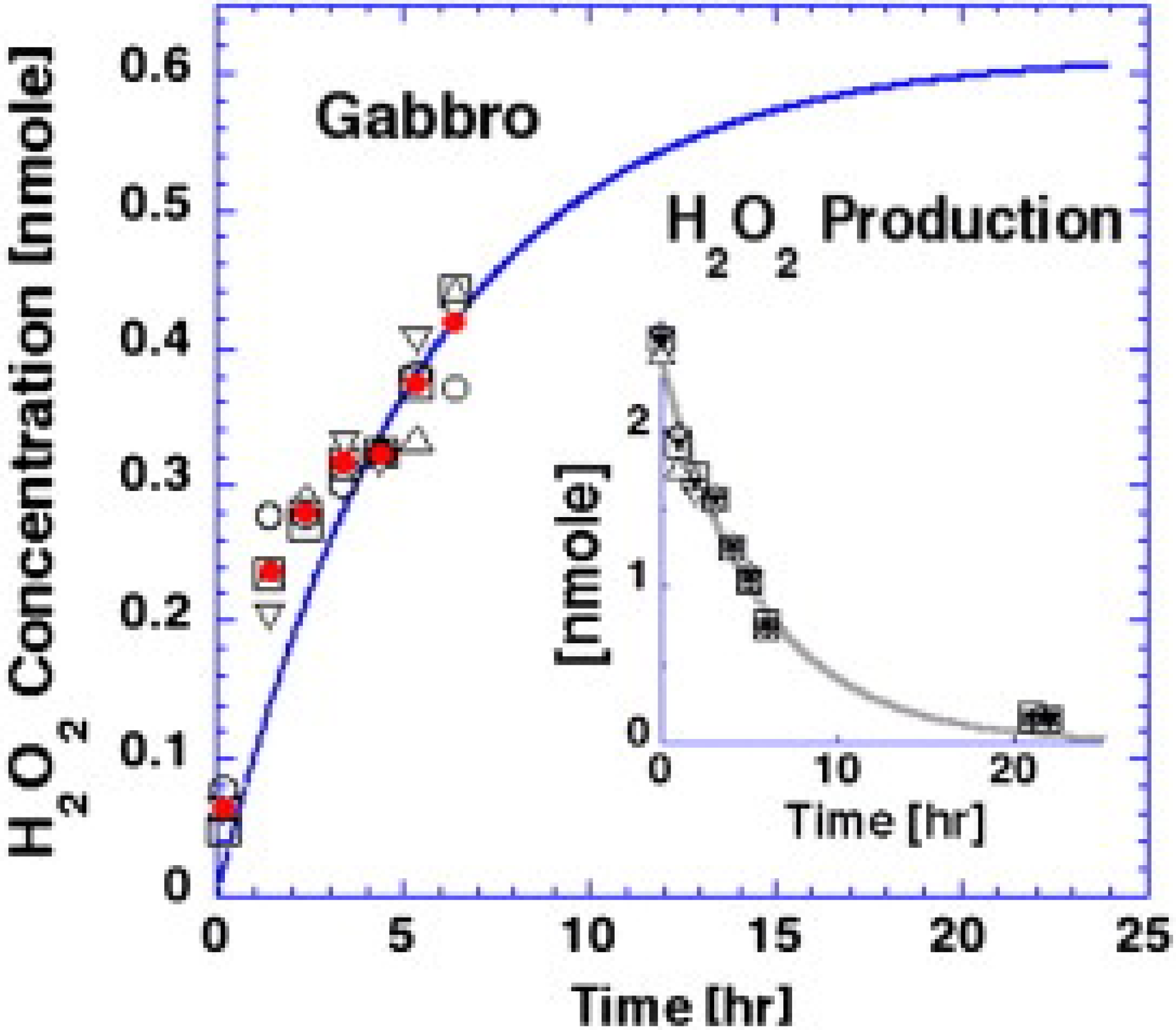
3. Results and Discussion
3.1. Earthworm Perfusion Test
3.2. Earthworm Pre-Mixed Soil Test
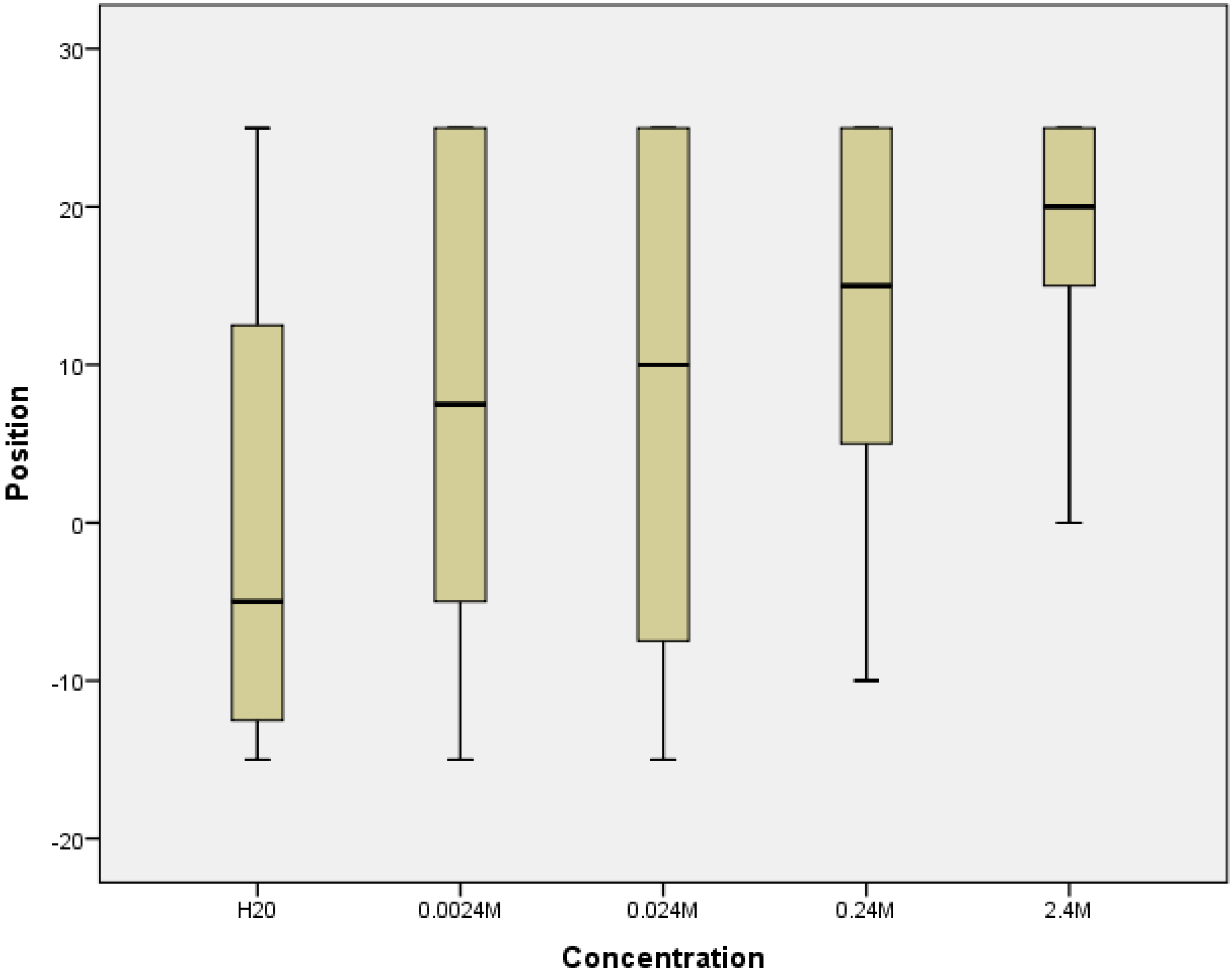
3.3. Daphnia Test

4. Conclusions
- Worms are not affected by H2O2 at the concentration that we used,
- The H2O2 could be decomposed more rapidly in soils than in water due to surface area effects,
- The higher presence of organic matter in soils may provide numerous oxidation pathways for the H2O2,
- There could be behavioral confounding factors due to handling the worms.
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Tributsch, H. When Snakes Awake: Animal and Earthquake Predictions; MIT Press: Cambridge, MA, USA, 1984. [Google Scholar]
- Whitehead, N.E.; Ulusoy, U.; Asahara, H.; Ikeya, M. Are any public-reported earthquake precursors valid? Nat. Hazards Earth Syst. Sci. 2004, 4, 463–468. [Google Scholar] [CrossRef]
- Fidani, C. Biological Anomalies around the 2009 L’Aquila Earthquake. Animals 2013, 3, 693–721. [Google Scholar] [CrossRef]
- Grant, R.A.; Halliday, T. Predicting the unpredictable; evidence of pre-seismic anticipatory behaviour in the common toad. J. Zool. 2010, 281, 263–271. [Google Scholar]
- Grant, R.A.; Halliday, T.; Balderer, W.P.; Leuenberger, F.; Newcomer, M.; Cyr, G.; Freund, F.T. Ground Water Chemistry Changes before Major Earthquakes and Possible Effects on Animals. Int. J. Environ. Res. Public Health 2011, 8, 1936–1956. [Google Scholar] [CrossRef] [PubMed]
- Tsunogai, U.; Wakita, H. Precursory chemical changes in ground water: Kobe earthquake, Japan. Science 1995, 269, 61–63. [Google Scholar] [CrossRef] [PubMed]
- King, C.Y.; Azuma, S.; Ohno, M.; Asai, Y.; He, P.; Kitagawa, Y.; Wakita, H. In search of earthquake precursors in the water-level data of 16 closely clustered wells at Tono, Japan. Geophys. J. Int. 2000, 143, 469–477. [Google Scholar] [CrossRef]
- Brancelj, A.; Žibrat, U.; Mezek, T.; Brancelj, I.R.; Dumont, H.J. Consecutive earthquakes temporarily restructured the zooplankton community in an Alpine Lake. Ann. Limnol. Int. J. Limnol. 2012, 48, 113–123. [Google Scholar] [CrossRef]
- Balk, M.; Bose, M.; Ertem, G.; Rogoff, D.A.; Rothschild, L.J.; Freund, F.T. Oxidation of water to hydrogen peroxide at the rock-water interface due to stress-activated electric currents in rocks. Earth Planet. Sci. Lett. 2009, 283, 87–92. [Google Scholar] [CrossRef]
- Freund, F.T.; Kulahci, I.G.; Cyr, G.; Ling, J.; Winnick, M.; Tregloan-Reed, J.; Freund, M.M. Air ionization at rock surface and pre-earthquake signals. J. Atmos. Sol.-Terr. Phys. 2009, 71, 1824–1834. [Google Scholar]
- Ikeya, M.; Takaki, S.; Takashimizu, D. Electric shocks resulting in seismic animal anomalous behaviors (SAABs). J. Phys. Soc. Jpn. 1996, 65, 710–712. [Google Scholar] [CrossRef]
- Čoja, T.; Zehetner, K.; Bruckner, A.; Watzinger, A.; Meyer, E. Efficacy and side effects of five sampling methods for soil earthworms (Annelida, Lumbricidae). Ecotoxicol. Environ. Saf. 2008, 71, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Minadakis, G.; Potirakis, S.M.; Nomicos, C.; Eftaxias, K. Linking electromagnetic precursors with earthquake dynamics: An approach based on nonextensive fragment and self-affine asperity models. Phys. A Stat. Mech. Appl. 2012, 391, 2232–2244. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grant, R.A.; Conlan, H. Behavioral Response of Invertebrates to Experimental Simulation of Pre-Seismic Chemical Changes. Animals 2015, 5, 206-213. https://doi.org/10.3390/ani5020206
Grant RA, Conlan H. Behavioral Response of Invertebrates to Experimental Simulation of Pre-Seismic Chemical Changes. Animals. 2015; 5(2):206-213. https://doi.org/10.3390/ani5020206
Chicago/Turabian StyleGrant, Rachel A., and Hilary Conlan. 2015. "Behavioral Response of Invertebrates to Experimental Simulation of Pre-Seismic Chemical Changes" Animals 5, no. 2: 206-213. https://doi.org/10.3390/ani5020206
APA StyleGrant, R. A., & Conlan, H. (2015). Behavioral Response of Invertebrates to Experimental Simulation of Pre-Seismic Chemical Changes. Animals, 5(2), 206-213. https://doi.org/10.3390/ani5020206