1. Introduction
Weaning is a stressful challenge for piglets because of the sudden changes of physiology and surrounding environment. These weaning stresses, usually presented as oxidative stress [
1], may reduce antioxidant status, immunity and intestinal functions, which results in a reduction in feed intake, weight gain and health of piglets, as well as an increase in diarrhea incidence, morbidity and mortality [
2]. To overcome these post-weaning disorders caused by oxidative stress, researchers report that the use of antibiotics, such as chlortetracycline (CTC), as in-feed supplements after weaning may help enhance weight gain by 16% and feed utilization by 7%, as well as reduce morbidity and mortality approximately 50% for weaned piglets [
3,
4]. However, the abuse or misuse of antibiotics in piglets’ feed can result in bacteria resistant to antibiotics and lead to potential residues in animal products (such as pork) and in the environment, which may enhance the possibility of antibiotic-resistant infections in humans [
5]. Therefore, the European Union, the United States and many other countries have reduced or prohibited the use of antibiotics in animal feed and found appropriate alternatives for antibiotics.
Studies in our labs have demonstrated that essential oils [
6], essential oils combined with mixed organic acids [
7], probiotics [
8], chito-oligosaccharide [
9] as well as natural plant herbs [
10] can potentially serve as antibiotic substitutes. Currently, the practice of using traditional herbal medicine is gaining more attention worldwide in animal health care systems [
11]. The addition of traditional Chinese herbs, especially
Coptis chinensis and
Forsythia suspense, have been used a potential non-antibiotic way to enhance anti-inflammatory and anti-microbial function, antioxidant status and performance in livestock [
10,
12,
13].
The fruit or leaf extract of
Forsythia suspensa Vahl (Oleaceae) could be widely used as Chinese medicine to help treat some infections, including pharyngitis, nephritis, febrile erysipelas, ulcers, tonsillitis, gonorrhea and acute respiratory syndrome [
14]. Previous studies in our lab have demonstrated that lignan, phenethyl alcohol glycoside, volatile oil as well as pentacyclic triterpenoids are the major active compounds of
Forsythia suspensa extract (FSE) [
15,
16], which has proven to have anti-oxidant [
17], anti-bacterial [
12,
18], anti-inflammatory [
19] and anti-allergy [
20] effects. Currently, our lab has used different stress models and different animal categories (e.g., piglets, broilers, laying hens and mice) to demonstrate the antioxidant properties and immune enhancement functions of FSE. Studies also show FSE can enhance performance by modulating intestinal permeability, antioxidant status and immune function in animals combined with chito-oligosaccharide [
13,
19] or berberine [
12]. However, there are few data to clarify the possibility of FSE as an antibiotic substitute and the possible mechanism functions of FSE. Therefore, the objective of this study is to evaluate the efficiency of FSE as an antibiotics substitute on performance, serum antioxidant status, fecal
Escherichia coli content and intestinal morphology in weaned piglets.
2. Materials and Methods
The study was carried out at the Animal Experimental Base (Fengning, Hebei, China) in the National Feed Engineering Technology Research Center of the Ministry of Agriculture Feed Industry Center. All the experimental procedures and operations used in the management and care of piglets were in agreement with the China Agricultural University Laboratory Animals Welfare and Animal Experimental Ethical Inspection (Beijing, China; No. AW09089102-1).
2.1. Experimental Products
Dried and ground forsythia fruits (about 100 g) were prepared and extracted by 80% ethanol (500 mL), sonicated for 1 h and then filtered. Ethanol was used to extract the residue twice and then rotary vaporization (Buchi, Rotavapor R-124, Flawil, Switzerland) was utilized to dry and combine the filtrates. The main functional ingredients in FSE are forsythoside A (1.65%), phillyrin (8.17%), forythialan A (4.13%) and phillygenin (1.67%) [
10]. An additional antibiotic mixture (chlortetracycline, CTC) was produced by Beijing Tongli Xing Department of Agricultural Science and Technology Company Limited (Beijing, China).
2.2. Experimental Animals, Management and Design
A total of 108 Duroc × (Landrace × Yorkshire) weaned piglets (28 days (d) weaned, average body weight of 8.68 ± 1.36 kg) were randomly assigned into 3 dietary treatments, 6 pens per treatment, 3 barrows and 3 gilts per pen. The treatments contained a corn-soybean meal basal diet (CTR), an antibiotic diet (basal diet + 75 mg/kg chlortetracycline; CTC), and an FSE diet (basal diet + 200 mg/kg FSE; FSE). The experiment included phase 1 (d 1 to 14), phase 2 (d 15 to 28) and phase 3 (d 29 to 35). As shown in
Table 1, nutrients in the diet met the recommended requirements (National Research Council, NRC, 2012) [
21].
All the piglets were raised in experimental pens (1.2 m × 2 m) fitted with a duckbill drinker, an adjustable stainless steel feeder and plastic slatted floors. The piglets were given access to water and fed ad libitum in powder form. Inside the pen, these piglets also had free access to feed (in mash form) and water ad libitum. The environment in the pig house, including the contents of carbon dioxide and ammonium in the air, ventilation intensity, humidity and temperature, was controlled automatically. The average temperature in house was controlled at 24–26 °C, while the relative humidity was maintained at 60%–70%. In order to prevent disease, the experimental house was cleaned every day and the immunization procedure was conducted every week. After 12 h of starvation, the individual weight of piglets and the feed weight of each pen were weighed on day 0, 14, 28 and 35 to calculate the average daily gain (ADG), average daily feed intake (ADFI) and feed efficiency (FE, ADFI/ADG). From day 0 to 28, the diarrhea score was monitored according to the previously described system [
22] from 1 to 5: 1, hard firm feces (rarely seen); 2, slightly soft feces; 3, soft, partially formed feces; 4, loose, semi-liquid feces (diarrhea); 5, watery, mucus-like feces (severe diarrhea). The determination of diarrhea rate was mainly dependent on the average diarrhea score following the formula: diarrhea rate (%) = diarrhea days × the number of diarrhea pigs/(experiment days × the total number of pigs) [
23].
2.3. Experimental Sample Collection and Analysis
During this experiment, a total of 2 kg representative feed samples were collected weekly. From day 12 to 14 and day 26 to 28, the rectal palpation was used to make sure approximately 100 g of fresh feces were collected using the grab sample technique. All the fecal samples were frozen at −20 °C immediately after collection until analysis. The feces collected after 3 days were pooled by pen and dried at 65 °C for 72 h. Before analysis, all these dried feces and feed samples were ground to pass through a 1-mm sieve.
The dry matter (DM), ether extract (EE) and crude protein (CP) of the feed and fecal samples were measured using the methods of Association of Official Agricultural Chemists (AOAC) [
24]. The gross energy (GE) in feed and fecal samples was determined by an automatic isoperibolic oxygen bomb calorimeter (Parr 1281, Automatic Energy Analyzer; Moline, IL, USA). Moreover, an atomic absorption spectrophotometer (Z-5000; Hitachi, Tokyo, Japan) was used to determine the content of chromium in feed and fecal samples. Organic matter (OM) was calculated as 1 − ash content (DM base). The calculation formula for total carbohydrates was as follows: the calculation = dry matter − crude protein − ether extract − ash [
25]. Nutrient digestibility was determined by the equation as follows: Apparent total tract digestibility
nutrient (ATTD) = 1 − (Cr
diet × nutrient
feces)/(Cr
feces × nutrient
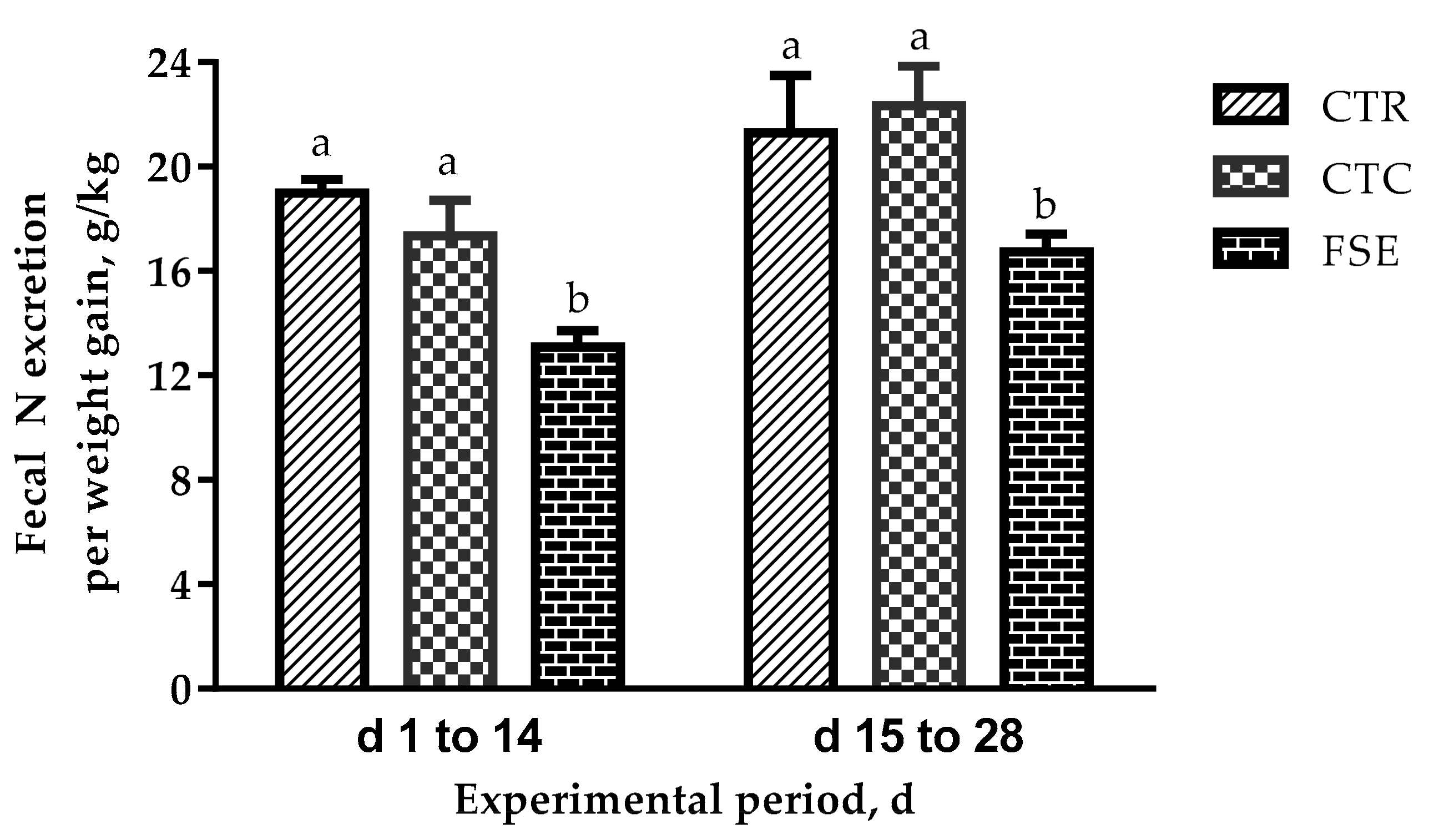
diet). The manure nitrogen output from piglets fed FSE during a 28-d period of the experiment was calculated by the equation as follows: fecal nitrogen (N) excretion per weight gain (g/kg) = (N intake (g/d) × (100 − ATTD of N))/([100 × ADG (kg/d)).
For the determination of fecal microbiota, the fresh fecal samples (about 100 g) of piglets in the CTR and FSE groups collected on day 14 and day 28 were first thawed at room temperature. A total of 1 g of fresh fecal sample was taken and transferred into a 9-mL diluent tube and then diluted 6 times serially in order to make sure every sample was fully dissolved. The test of fecal E. coli concentrations in fresh fecal samples was carried out within a day after collection. The total content of E. coli was determined using Maconkey agar to plate 0.1 mL diluent. An electro-heating standing-temperature cultivator (37 °C) was used to incubate all the petri dishes for 24 h. Before statistical analysis, the fecal E. coli concentrations were transformed (Log).
After starvation for 12 h, approximately 8 mL of blood was collected from a piglet near the average group body weight in each pen via jugular vein puncture into a vacutainer at 7:00 a.m. on day 28 (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ, USA). After stewing for 3 h, all the collected blood samples were centrifuged at 3000× g for 15 min at 4 °C to get the serum samples, which were also stored at −20 °C until analysis. Estimation of triglyceride (TG) and blood urea nitrogen (BUN) contents in serum were conducted by Hitachi 7600 Automatic Biochemical Instrument. An ELISA kit (IgG, IgM and IgA quantitation kit; Bethyl Laboratories, Inc., Montgomery, TX, USA) was used to determine the concentrations of serum immunoglobulins (including immunoglobulin G, immunoglobulin M and immunoglobulin A). Determination of total antioxidant capacity (T-AOC), catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px) and malondialdehyde (MDA) levels in serum were conducted by spectrophotometric methods using a spectrophotometer (Leng Guang SFZ1606017568, Shanghai, China) following the instructions of the kit’s manufacturer (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
On day 28, the aseptic duodenal, jejunal and ileal samples (about 5 cm fragment in the middle of each intestine, duodenum was selected as the proximal 1/3 of the small intestine, jejunum as the 1/3 mid and ileum as 1/3 distal part) were collected from slaughtered barrows (near the average group body weight) selected in each pen for the determination of intestinal morphology. Then, 10% neutral buffered formalin was used to fix rapidly these histological samples for slicing. After 48 h of fixation, the sections of intestinal tissues were washed, excised, dehydrated, as well as embedded in the paraffin wax, and then 5 transverse sections were sliced, installed on glass slides and dyed with eosin and hematoxylin. At least 20 orientated villi and their adjoining crypts were selected randomly on each slice and measured to calculate the average villus height and crypt depth via a light microscope in small caps using a calibrated 10-fold eyepiece graticule. The ratio of villus height to crypt depth was calculated and used for further analysis.
2.4. Statistical Analysis
The mixed model of SAS (version 9.2, 2008) [
26] was used for variance analysis of all the data. The dietary treatments were fixed effects, while sex and body weight of pigs were the random effects. For the analysis of growth performance and diarrhea rate, the pen was treated as the statistical unit, whereas for the analysis of other data, individual piglets were taken as a statistical unit. The Student–Newman–Keul multiple range test was used for determining the statistical differences among all the treatments. Significant difference between the mean value was defined at
p ≤ 0.05, while a trend for the significance between the mean value was designated at 0.05 <
p ≤ 0.10.
4. Discussion
Weaning stress may cause a reduction of growth rate and feed intake in piglets, while current results show dietary FSE supplementation can enhance ADG and ADFI in phase 2, but there is no difference of performance among dietary treatments in phase 1. Similar results are also shown in the study by Han et al. [
18], which demonstrated that during the first period, performance has no difference in the FSE group, while it experiences a significant enhancement during the finisher and overall phase in broilers. This indicates that the beneficial effect of FSE on piglets may relate to its cumulative effect in animals. However, according to the study of Zhao et al. [
19], dietary supplementation with 100 mg/kg FSE can improve ADG and FE in the first 2 weeks. These different findings may relate to the amount and composition of FSE in different studies. The present study showed that there is no significant difference of FE between treatments, while other studies show dietary plant polyphenols supplementation can improve the FE in pigs via the improvement of nutrients digestibility [
27] and the enhancement of health status via inhibiting inflammation [
28]. These contradictory functions of FSE on FE in different studies may also relate to the type, additive amount and chemical composition of Chinese plant polyphenols. The current study also shows that the effects of FSE on enhancing growth performance is better than CTC, which is partly in line with the study of Han et al. [
18], who also indicated that FSE has the potential to replace antibiotics in improving the performance and intestinal health of animals. The reason for the improved performance may also be due to the decreased diarrhea rate and improved nutrient utilization of piglets fed an FSE diet.
After weaning, piglets are susceptible to disease due to environmental changes, which usually cause severe post-weaning diarrhea. The present study shows that piglets fed a CTC and FSE diet experienced a significant reduction in diarrhea rates in phase 1 and 2; this could possibly be due to the lower content of
E. coli in feces of piglets fed a CTC and FSE diet since diarrhea caused by
E. coli is a major challenge for post-weaning pigs [
8]. The present result is in agreement with the study of Han et al. [
18], who reported that FSE can regulate intestinal flora via reducing the cecal
E. coli counts in vivo and inhibiting the reproduction of
E. coli K88,
Staphylococcus aureus and
salmonella in vitro. The result may be because polyphenol in FSE can increase the fecal pH value and lower the concentrations of volatile fatty acids [
27]. Another reason may be that the forsythiaside and phillyrin in FSE have strong broad-spectrum antimicrobial activity [
29], which can effectively inhibit
E. coli,
Pseudomonas aeruginosa and
S. aureus [
30,
31]. Moreover, the essential oils from FSE have also demonstrated their effectiveness in inhibiting the growth of
S. aureus,
Bacillus subtilis,
E. coli,
P. aeruginosa,
Candida albicans and
Aspergillus Niger [
32]. In addition, according to a study by Zhang et al. [
12], FSE may also increase the level of
Lactobacillus while reducing the level of
E. coli in the cecum of broilers on day 21 and 42, thus improving the structure of intestinal flora in broilers. In our present study, we consider the number of CFUs of
E. coli only; the differences may not represent a real reduction in
E. coli, therefore, this finding still needs to be further estimated in our following studies.
During the first two weeks after weaning, piglets may face severe challenges when they utilize nutrients, which is mainly due to disorders in the digestive and absorption systems. In the present study, the ATTD of nutrients were enhanced in piglets fed an FSE diet in phase 1, which is partly in agreement with the previous study of Han et al. [
18] and Zhang et al. [
12]. The digestibility of nutrients strongly increased in piglets fed FSE compared to CTR in phase 1 and tended to be enhanced in phase 2, which can normally lead to an improvement of ADG and FE [
19]. However, our study only found the enhancement of ADG in phase 2, from day 15 to 35 and day 1 to 28, which may be due to that the enhancement of ADFI in phase 2 as well as the difference of the additive amount and composition of FSE. After weaning within 2 weeks, the digestive system of piglets is not well developed and Chinese medicine usually has an accumulative effect for animals, which may lead to the positive effect of FSE on ADG to be easier to find in phase 2, from day 15 to 35 and day 1 to 28 [
18]. Yet this finding still remains to be investigated in further study. Moreover, the improved nutrient digestibility in the current study is in line with the increased villus height to crypt depth ratio and decreased crypt depth in ileum of piglets fed an FSE diet. This may be because FSE can help improve small intestinal villus morphology, enhancing the growth of villus as well as the villus height to crypt depth ratio, which in turn may increase the absorption of DM and CP [
12]. In addition, FSE can also improve the proliferation of peripheral blood lymphocytes and intestinal permeability, so as to increase the intestinal capacity to utilize nutrients [
20]. The higher numerical changes of the villus height, crypt depth and their ratio in the small intestine probably indicates FSE may benefit and help in the development of the small intestine in weaned piglets [
18]. A possible reason for the obviously improved nutrient digestibility in the early phase for piglets may be that after weaning, the development of the intestine and activity of digestible enzymes are poor, while the intestine is better developed after two weeks in the later phase. The reduction of manure N output from piglets fed diets based on a corn-soybean meal diet supplemented with FSE is mainly due to the improved ATTD of CP.
When oxidants and antioxidants are in an unbalanced state (oxidative stress reaction), a large number of reactive oxygen species (ROS) will be produced. These excessive ROS and free radicals can lead to lipid peroxidation, DNA damage, cell apoptosis or cell cycle arrest, eventually leading to cell death [
33]. In the present study, the contents of T-AOC, SOD and CAT are enhanced, whereas the concentration of MDA is decreased in piglets fed an FSE diet. This may because FSE could be an effective antioxidant in vitro and in vivo. In vitro studies have shown that the scavenging rates of 100 g/mL and 250 g/mL FSE for 1,1-diphenyl-2-trinitrophenylhydrazine (DPPH) free radical can reach 77.2% and 81.3% respectively, which are comparable to vitamin C [
17]. Moreover, the phenolic hydroxyl ortho-position on the phenyl ring of forsythiaside in FSE has a strong antioxidant and free radical scavenging ability, which can effectively protect cells from hydrogen peroxide-induced cell damage (reducing ROS and MDA levels) and mitochondrial-dependent cell apoptosis [
10]. In vivo studies have shown that FSE can effectively alleviate the oxidative damage of diquat to Sprague-Dawley (SD) rats, reduce MDA content and inflammatory factors in the liver and serum [
10], which is mainly because forsythiaside in FSE can activate non-enzymatic system, improve total antioxidant capacity and increase the expression of antioxidant enzymes in cells by promoting the level of nuclear factor erythroid-2-related factor 2 in the nucleus [
29], so as to activate antioxidant enzymes system (such as SOD, CAT and GSH-Px). Moreover, the polyphenols in FSE can also play an important role in enhancing the activities of antioxidant enzymes in the blood, liver, spleen and kidneys, which largely contributes to improving the health status and growth performance of weaned piglets [
34].




{kind=link}