Effects of the Methionine Hydroxyl Analogue Chelate Zinc on Antioxidant Capacity and Liver Metabolism Using 1H-NMR-Based Metabolomics in Aged Laying Hens

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management
2.2. Diet Treatment
2.3. Antioxidant and Serum Biochemical Indices
2.4. Sample Preparation for 1H-NMR
2.5. Statistical Analysis
3. Results
3.1. Antioxidant Indices
3.2. Serum Biochemical Indices
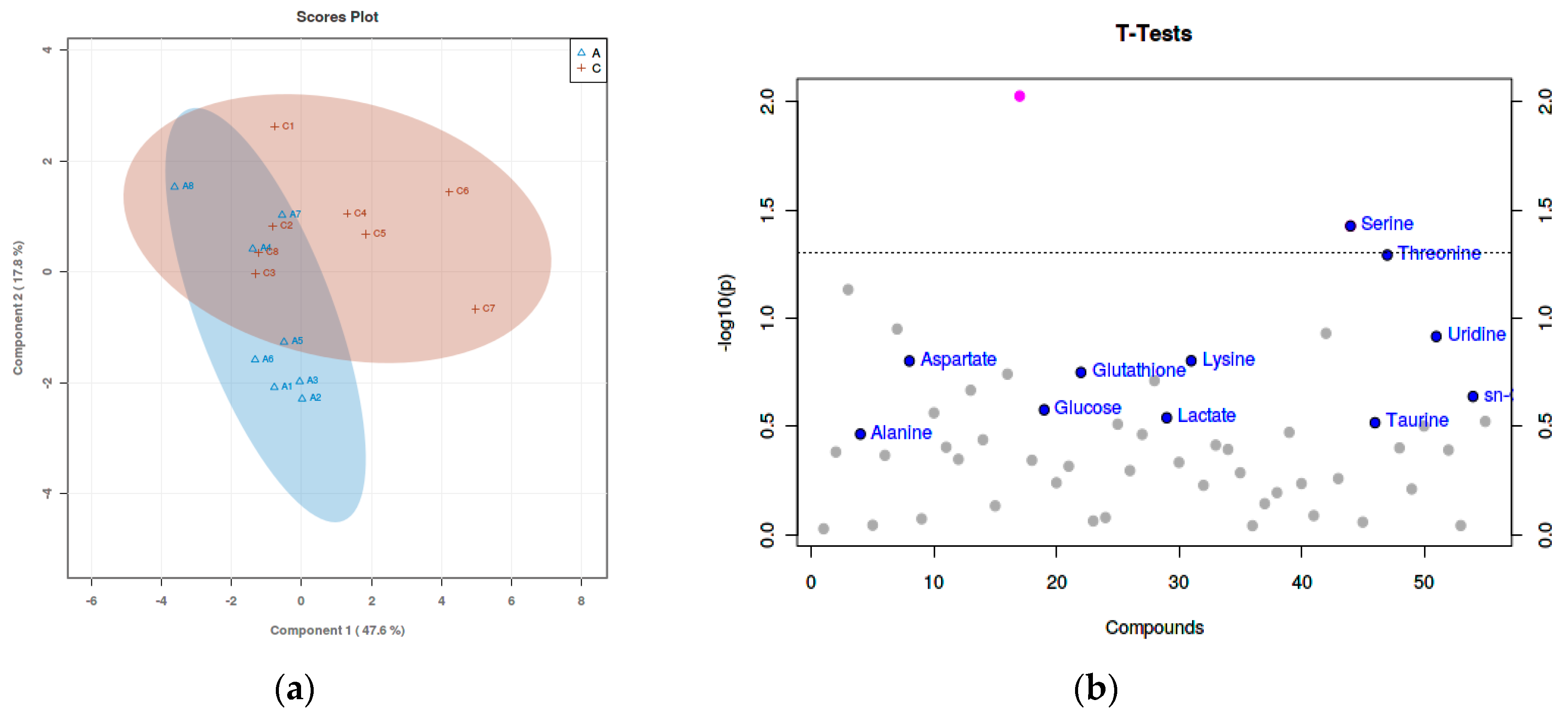
3.3. Metabolite Identification
3.4. Result of Pathway Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Molnár, A.; Maertens, L.; Ampe, B.; Buyse, J.; Kempen, I.; Zoons, J.; Delezie, E. Changes in egg quality traits during the last phase of production: Is there potential for an extended laying cycle? Br. Poult. Sci. 2016, 57, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Mocchegiani, E.; Rink, L. Correlation between zinc status and immune function in the elderly. Biogerontology 2006, 7, 421. [Google Scholar] [CrossRef] [PubMed]
- Zhan, H.Q.; Li, L.L.; Gong, Y.J.; Zheng, Y.X.; Zou, X.T. Effects of dietary Zn-methionine supplementation on the laying performance, egg quality, antioxidant capacity, and serum parameters of laying hens. Poult. Sci. 2018, 98, 923–931. [Google Scholar]
- Cao, J.; Henry, P.R.; Guo, R.; Holwerda, R.A.; Toth, J.P.; Littell, R.C.; Miles, R.D.; Ammerman, C.B. Chemical characteristics and relative bioavailability of supplemental organic zinc sources for poultry and ruminants. J. Anim. Sci. 2000, 78, 2039–2054. [Google Scholar] [CrossRef] [PubMed]
- Nitrayova, S.; Windisch, W.; von Heimendahl, E.; Müller, A.; Bartelt, J. Bioavailability of zinc from different sources in pigs. J. Anim. Sci 2012, 90, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, S.; Timmons, J.; Ao, T.; Paul, M.; Macalintal, L.; Pescatore, A.; Cantor, A.; Ford, M.; Dawson, K.A. Effect of zinc imprinting and replacing inorganic zinc with organic zinc on early performance of broiler chicks. Poult. Sci. 2017, 96, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Bun, S.D.; Guo, Y.M.; Guo, F.C.; Ji, F.J.; Cao, H. Influence of organic zinc supplementation on the antioxidant status and immune responses of broilers challenged with Eimeria tenella. Poult. Sci. 2011, 90, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Akbari Moghaddam Kakhki, R.; Bakhshalinejad, R.; Hassanabadi, A.; Ferket, P. Effects of dietary organic zinc and α-tocopheryl acetate supplements on growth performance, meat quality, tissues minerals, and α-tocopherol deposition in broiler chickens. Poult. Sci. 2017, 96, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.N.; Liu, F.X.; Qi, X.; Ji, S.; Ma, S.X.; Liu, X.; Wang, Z.P.; Gao, Y.P. Effects of methionine hydroxyl analog chelated zinc on laying performance, eggshell quality, eggshell mineral deposition, and activities of Zn-containing enzymes in aged laying hens. Poult. Sci. 2018, 97, 3587–3593. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.N.; Liu, F.X.; Qi, X.; Ji, S.; Cui, L.; Wang, Z.P.; Gao, Y.P. Effects of organic zinc on tibia quality, mineral deposit, and metallothionein expression level of aged hens. Poult. Sci. 2019, 98, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Bao, S.Y.; Bolin, E.R.; Burris, C.L.; Xu, X.H.; Sun, Q.H.; Killilea, D.W.; Shen, O.; Ziouzenkova, O.; Belury, M.A.; et al. Zinc deficiency augments leptin production and exacerbates macrophage infiltration into adipose tissue in mice fed a high-fat diet. J. Nutr. 2013, 143, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, P.; Pigera, S.; Galappatthy, P.; Katulanda, P.; Constantine, G.R. Zinc and diabetes mellitus: Understanding molecular mechanisms and clinical implications. J. Facul. Pharm. 2015, 23, 44. [Google Scholar] [CrossRef] [PubMed]
- Markley, J.L.; Brüschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr. Opin. Biotech. 2017, 43, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Stacklies, W.; Redestig, H.; Scholz, M.; Walther, D.; Selbig, J. PCA Methods--a bioconductor package providing PCA methods for incomplete data. Bioinformatics 2007, 23, 1164–1167. [Google Scholar] [CrossRef] [PubMed]
- Mevik, B.H.; Wehrens, R. The pls Package: Principal Component and Partial Least Squares Regression in R. J. Stat. Soft 2007, 18, 23. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc coordination environments in proteins as redox sensors and signal transducers. Antioxid Redox Signal. 2006, 8, 1419–1441. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: Role in immunity, oxidative stress and chronic inflammation. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Mccord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [PubMed]
- Noor, R.; Mittal, S.; Iqbal, J. Superoxide dismutase--applications and relevance to human diseases. Med. Sci. Monitor 2002, 8, 210–215. [Google Scholar]
- Oteiza, P.I.; Clegg, M.S.; Zago, M.P.; Keen, C.L. Zinc deficiency induces oxidative stress and AP-1 activation in 3T3 cells. Free Radical Bio. Med. 2000, 28, 1091–1099. [Google Scholar] [CrossRef]
- Orr, W.C.; Sohal, R.S. Extension of life-span by overexpression of superoxide dismutase and catalase in Drosophila melanogaster. Science 1994, 263, 1128–1130. [Google Scholar] [CrossRef] [PubMed]
- Dimitrios, T. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar]
- Chen, W.; Wang, S.; Zhang, H.X.; Ruan, D.; Xia, W.G.; Cui, Y.Y.; Zheng, C.T.; Lin, Y.C. Optimization of dietary zinc for egg production and antioxidant capacity in Chinese egg-laying ducks fed a diet based on corn-wheat bran and soybean meal. Poult. Sci. 2017, 96, 2336–2343. [Google Scholar] [CrossRef] [PubMed]
- Abedini, M.; Shariatmadari, F.; Karimi Torshizi, M.A.; Ahmadi, H. Effects of zinc oxide nanoparticles on the egg quality, immune response, zinc retention, and blood parameters of laying hens in the late phase of production. J. Anim. Physiol. Anim. Nutr. 2018, 102, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Alagawany, M.; Salah, A.S.; Abdel-Latif, M.A.; Farghly, M.F.A. Effects of Dietary Supplementation of Zinc Oxide and Zinc Methionine on Layer Performance, Egg Quality, and Blood Serum Indices. Biol. Trace Elem. 2018, 184, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Alagawany, M.; Amer, S.A.; Arif, M.; Kmm, W.; El-Kholy, M.S. Effect of dietary supplementation of organic zinc on laying performance, egg quality and some biochemical parameters of laying hens. J. Anim. Physiol. Anim. Nutr. 2017, 102, e542–e549. [Google Scholar] [CrossRef] [PubMed]
- Azzam, M.M. Influence of L-threonine supplementation on goblet cell numbers, histological structure and antioxidant enzyme activities of laying hens reared in a hot and humid climate. Br. Poult. Sci. 2012, 53, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Cheng, Y.F.; Li, X.H.; Yang, W.L.; Wen, C.; Zhuang, S.; Zhou, Y.M. Effects of threonine supplementation on the growth performance, immunity, oxidative status, intestinal integrity, and barrier function of broilers at the early age. Poult. Sci. 2017, 96, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Habte-Tsion, H.M.; Ren, M.; Liu, B.; Ge, X.; Xie, J.; Chen, R. Threonine modulates immune response, antioxidant status and gene expressions of antioxidant enzymes and antioxidant-immune-cytokine-related signaling molecules in juvenile blunt snout bream (Megalobrama amblycephala). Fish. Shellfish Immun. 2016, 51, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.N.; Liu, S.G.; Qu, Z.X.; Meng, G.H.; Gao, Y.P. Effects of dietary threonine levels on growth performance, serum biochemical indexes, antioxidant capacities, and gut morphology in broiler chickens. Poult. Sci. 2017, 96, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Akrabawi, S.S.; Kratzer, F.H. Effects of arginine or serine on the requirement for glycine by the chick. J. Nutr. 1968, 95, 41. [Google Scholar] [CrossRef] [PubMed]
- Stekol, J.A.; Weiss, S.; Weiss, K.W. Vitamin B 12 and folic acid in the synthesis of choline in the rat. Arch. Biochem. Bioph. 1952, 36, 5–10. [Google Scholar] [CrossRef]
- Baker, D.H.; Sugahara, M.; Scott, H.M. The glycine-serine interrelationship in chick nutrition. Poult. Sci. 1968, 47, 1376–1377. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H. Sulfur Amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr. 2004, 24, 539–577. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, O.D.; Labuschagne, C.F.; Adams, P.D.; Vousden, K.H. Serine Metabolism Supports the Methionine Cycle and DNA/RNA Methylation through De Novo ATP synthesis in cancer cells. Mol. Cell 2016, 61, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Vance, D.E. Thematic Review Series: Glycerolipids. Phosphatidylcholine and choline homeostasis. J. Lipid Res. 2008, 49, 1187–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.W.; Wu, Z.L.; Dai, Z.L.; Yang, Y.; Wang, J.J.; Wu, G.Y. Glycine metabolism in animals and humans: Implications for nutrition and health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef] [PubMed]
- El Hendy, H.A.; Yousef, M.I.; Abo El-Naga, N.I. Effect of dietary zinc deficiency on hematological and biochemical parameters and concentrations of zinc, copper, and iron in growing rats. Toxicology 2001, 167, 163–170. [Google Scholar] [CrossRef]
- Quest, A.F.; Bloomenthal, J.; Bardes, E.S.; Bell, R.M. The regulatory domain of protein kinase C coordinates four atoms of zinc. J. Biol. Chem. 1992, 267, 10193–10197. [Google Scholar] [PubMed]
- Ma, Z.; Zhang, J.; Ma, H.; Dai, B.; Zheng, L.; Miao, J.; Zhang, Y. The influence of dietary taurine and reduced housing density on hepatic functions in laying hens. Poult. Sci. 2014, 93, 1724–1736. [Google Scholar] [CrossRef] [PubMed]
- Kolho, K.L.; Pessia, A.; Jaakkola, T.; de Vos, W.M.; Velagapudi, V. Faecal and serum metabolomics in paediatric inflammatory bowel disease. J. Crohns Colitis 2017, 11, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, L.; Zhang, P.F.; Liu, X.Q.; Zhang, W.D.; Ding, Z.P.; Wang, S.W.; Shen, W.; Min, L.J.; Hao, Z.H. Regulation of egg quality and lipids metabolism by Zinc Oxide Nanoparticles. Poult. Sci. 2016, 95, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Lu, L.; Wang, R.L.; Lei, H.L.; Li, S.F.; Zhang, L.Y.; Luo, X.G. Effects of supplemental zinc source and level on antioxidant ability and fat metabolism-related enzymes of broilers1. Poult. Sci. 2015, 94, 2686–2694. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | % | Nutrients | % |
|---|---|---|---|
| Corn | 58.50 | Metabolic energy (MJ/kg) | 12.29 |
| Soybean meal | 23.00 | Crude protein 2 | 15.47 |
| Ground limestone | 8.74 | Calcium 2 | 3.60 |
| CaHPO4 | 1.10 | Total phosphorus | 0.62 |
| Wheat bran | 7.50 | Available phosphorus | 0.32 |
| Soybean oil | 0.50 | Lysine | 0.94 |
| NaCl | 0.24 | Methionine | 0.44 |
| Methionine | 0.16 | Threonine | 0.70 |
| Threonine | 0.03 | Zinc 2 (mg/kg) | 35.08 |
| Lysine | 0.05 | ||
| Premix 1 | 0.18 | ||
| Total | 100.00 |
| Dietary Zinc Supplementation | Calculated (mg/kg) | Analyzed (mg/kg) |
|---|---|---|
| 80 mg/kg ZnSO4 | 115.08 | 115.53 |
| 20 mg/kg MHA-Zn | 55.08 | 55.38 |
| 40 mg/kg MHA-Zn | 75.08 | 75.97 |
| 80 mg/kg MHA-Zn | 115.08 | 114.18 |
| Items | 80 mg/kg ZnSO4 | 20 mg/kg MHA-Zn | 40 mg/kg MHA-Zn | 80 mg/kg MHA-Zn | p-Value | |
|---|---|---|---|---|---|---|
| Cu/Zn-SOD | Liver (U/mgprot) | 66.62 ± 2.80 b | 61.25 ± 7.92 b | 67.90 ± 5.82 b | 75.23 ± 3.97 a | 0.003 |
| Serum (U/mL) | 60.09 ± 1.43 b | 54.45 ± 3.25 c | 56.04 ± 1.22 c | 64.89 ± 2.35 a | <0.001 | |
| T-AOC | Liver (U/gprot) | 1.76 ± 0.18 b | 1.49 ± 0.24 b | 1.81 ± 0.17 ab | 2.12 ± 0.40 a | 0.006 |
| Serum (U/mL) | 9.31 ± 0.28 ab | 7.73 ± 0.58 c | 8.98 ± 0.57 b | 9.81 ± 0.23 a | <0.001 | |
| MDA | Liver (nmol/gprot) | 3.41 ± 0.34 ab | 3.72 ± 0.37 a | 3.26 ± 0.35 ab | 3.04 ± 0.61 b | 0.088 |
| Serum (nmol/mL) | 3.57 ± 0.16 bc | 4.36 ± 0.42 a | 4.03 ± 0.46 ab | 3.48 ± 0.49 c | 0.008 | |
| Item | 80 mg/kg ZnSO4 | 20 mg/kg MHA-Zn | 40 mg/kg MHA-Zn | 80 mg/kg MHA-Zn | p-Value |
|---|---|---|---|---|---|
| GPT/mmol/L | 1.53 ± 0.67 | 2.26 ± 0.15 | 1.76 ± 0.66 | 1.43 ± 0.15 | 0.229 |
| SGOT/mmol/L | 187.2 ± 1.35 ab | 214.2 ± 13.15 a | 189.7 ± 13.05 ab | 157.7 ± 23.72 b | 0.018 |
| TP (g/L) | 42 ± 2.86 ab | 35.83 ± 5.38 b | 47.67 ± 3.02 a | 43.86 ± 4.11 a | 0.037 |
| ALB (g/L) | 17.43 ± 1.05 | 14.9 ± 2.69 | 16.2 ± 1.68 | 17.5 ± 1.18 | 0.302 |
| GLB (g/L) | 24.57 ± 3.85 b | 20.93 ± 2.69 b | 31.47 ± 1.79 a | 26.37 ± 3.25 ab | 0.016 |
| ALB/GLB (%) | 0.67 ± 0.11 | 0.71 ± 0.04 | 0.59 ± 0.09 | 0.67 ± 0.06 | 0.38 |
| UN/mmol/L | 0.29 ± 0.07 b | 0.24 ± 0.09 b | 0.49 ± 0.28 b | 1.06 ± 0.26 a | 0.025 |
| TC/mmol/L | 2.25 ± 0.18 | 1.89 ± 0.27 | 2.24 ± 0.5 | 2.31 ± 0.58 | 0.625 |
| TG/mmol/L | 13.93 ± 1.22 | 13.97 ± 3.5 | 13.23 ± 0.37 | 10.06 ± 2.18 | 0.163 |
| UA/mmol/L | 0.067 ± 0.011 | 0.101 ± 0.016 | 0.089 ± 0.037 | 0.091 ± 0.009 | 0.343 |
| Groups | Pathway Name | Match Status | p-Value | -log(p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|---|
| A/C | Glycine, serine, and threonine metabolism | 8/33 | 0.05481 | 2.9039 | 1 | 0.64662 | 0.5867 |
| Beta-alanine metabolism | 4/20 | 0.17964 | 1.7168 | 1 | 0.64662 | 0.4510 | |
| Alanine, aspartate, and glutamate metabolism | 9/23 | 0.36335 | 1.0124 | 1 | 0.64662 | 0.6733 | |
| Glutathione metabolism | 5/26 | 0.46206 | 0.77205 | 1 | 0.64662 | 0.4183 | |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | 2/4 | 0.48015 | 0.73366 | 1 | 0.64662 | 1 | |
| A/D | Glutathione metabolism | 5/26 | 0.16582 | 1.7969 | 1 | 0.56967 | 0.4183 |
| Beta-alanine metabolism | 4/20 | 0.20855 | 1.5676 | 1 | 0.56967 | 0.4510 | |
| D-glutamine and D-glutamate metabolism | 2/5 | 0.39423 | 0.93081 | 1 | 0.64063 | 1 | |
| Glycine, serine, and threonine metabolism | 8/33 | 0.4149 | 0.87971 | 1 | 0.64684 | 0.5867 | |
| Alanine, aspartate, and glutamate metabolism | 9/23 | 0.47665 | 0.74098 | 1 | 0.64684 | 0.6733 |
| Item | 80 mg/kg ZnSO4 | 20 mg/kg MHA-Zn | 40 mg/kg MHA-Zn | 80 mg/kg MHA-Zn | p-Value |
|---|---|---|---|---|---|
| Pyruvic acid (mmol/L) | 0.066 ± 0.010 | 0.046 ± 0.007 | 0.057 ± 0.006 | 0.060 ± 0.014 | 0.237 |
| Glycine (mmol/L) | 0.601 ± 0.040 | 0.663 ± 0.076 | 0.592 ± 0.038 | 0.516 ± 0.065 | 0.114 |
| Serine (mmol/L) | 0.518 ± 0.050 b | 0.652 ± 0.094 a | 0.679 ± 0.047 a | 0.605 ± 0.078 b | 0.040 |
| Cysteine (mmol/L) | 0.096 ± 0.011 a | 0.086 ± 0.010 a | 0.077 ± 0.008 b | 0.056 ± 0.007 b | 0.042 |
| Choline (mmol/L) | 0.079 ± 0.011 | 0.085 ± 0.009 | 0.065 ± 0.009 | 0.057 ± 0.009 | 0.199 |
| Threonine (mmol/L) | 0.359 ± 0.035 b | 0.446 ± 0.073 ab | 0.483 ± 0.045 a | 0.386 ± 0.053 b | 0.045 |
| Creatine (mmol/L) | 0.057 ± 0.008 | 0.049 ± 0.005 | 0.076 ± 0.009 | 0.054 ± 0.008 | 0.148 |
| Betaine (mmol/L) | 0.229 ± 0.036 | 0.293 ± 0.052 | 0.240 ± 0.040 | 0.260 ± 0.039 | 0.342 |
| Methionine (mmol/L) | 0.143 ± 0.012 | 0.152 ± 0.021 | 0.168 ± 0.015 | 0.172 ± 0.012 | 0.368 |
| Glutamic acid (mmol/L) | 0.774 ± 0.066 | 0.806 ± 0.013 | 0.726 ± 0.055 | 0.655 ± 0.080 | 0.485 |
| Glutamine (mmol/L) | 0.376 ± 0.045 | 0.431 ± 0.055 | 0.527 ± 0.050 | 0.382 ± 0.047 | 0.163 |
| Glutathione (mmol/L) | 0.067 ± 0.010 | 0.057 ± 0.012 | 0.048 ± 0.008 | 0.042 ± 0.009 | 0.134 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, X.; Ma, S.; Liu, X.; Wang, Y.; Liu, Y.; Gao, Y.; Min, Y. Effects of the Methionine Hydroxyl Analogue Chelate Zinc on Antioxidant Capacity and Liver Metabolism Using 1H-NMR-Based Metabolomics in Aged Laying Hens. Animals 2019, 9, 898. https://doi.org/10.3390/ani9110898
Qi X, Ma S, Liu X, Wang Y, Liu Y, Gao Y, Min Y. Effects of the Methionine Hydroxyl Analogue Chelate Zinc on Antioxidant Capacity and Liver Metabolism Using 1H-NMR-Based Metabolomics in Aged Laying Hens. Animals. 2019; 9(11):898. https://doi.org/10.3390/ani9110898
Chicago/Turabian StyleQi, Xi, Shuxue Ma, Xing Liu, Yamin Wang, Yinglu Liu, Yupeng Gao, and Yuna Min. 2019. "Effects of the Methionine Hydroxyl Analogue Chelate Zinc on Antioxidant Capacity and Liver Metabolism Using 1H-NMR-Based Metabolomics in Aged Laying Hens" Animals 9, no. 11: 898. https://doi.org/10.3390/ani9110898
APA StyleQi, X., Ma, S., Liu, X., Wang, Y., Liu, Y., Gao, Y., & Min, Y. (2019). Effects of the Methionine Hydroxyl Analogue Chelate Zinc on Antioxidant Capacity and Liver Metabolism Using 1H-NMR-Based Metabolomics in Aged Laying Hens. Animals, 9(11), 898. https://doi.org/10.3390/ani9110898