Genetic Diversity of IGHM and IGHE in the Leporids Revealed Different Patterns of Diversity in the Two European Rabbit Subspecies (O. cuniculus algirus and O. c. cuniculus)



Abstract
:Simple Summary
Abstract
1. Introduction
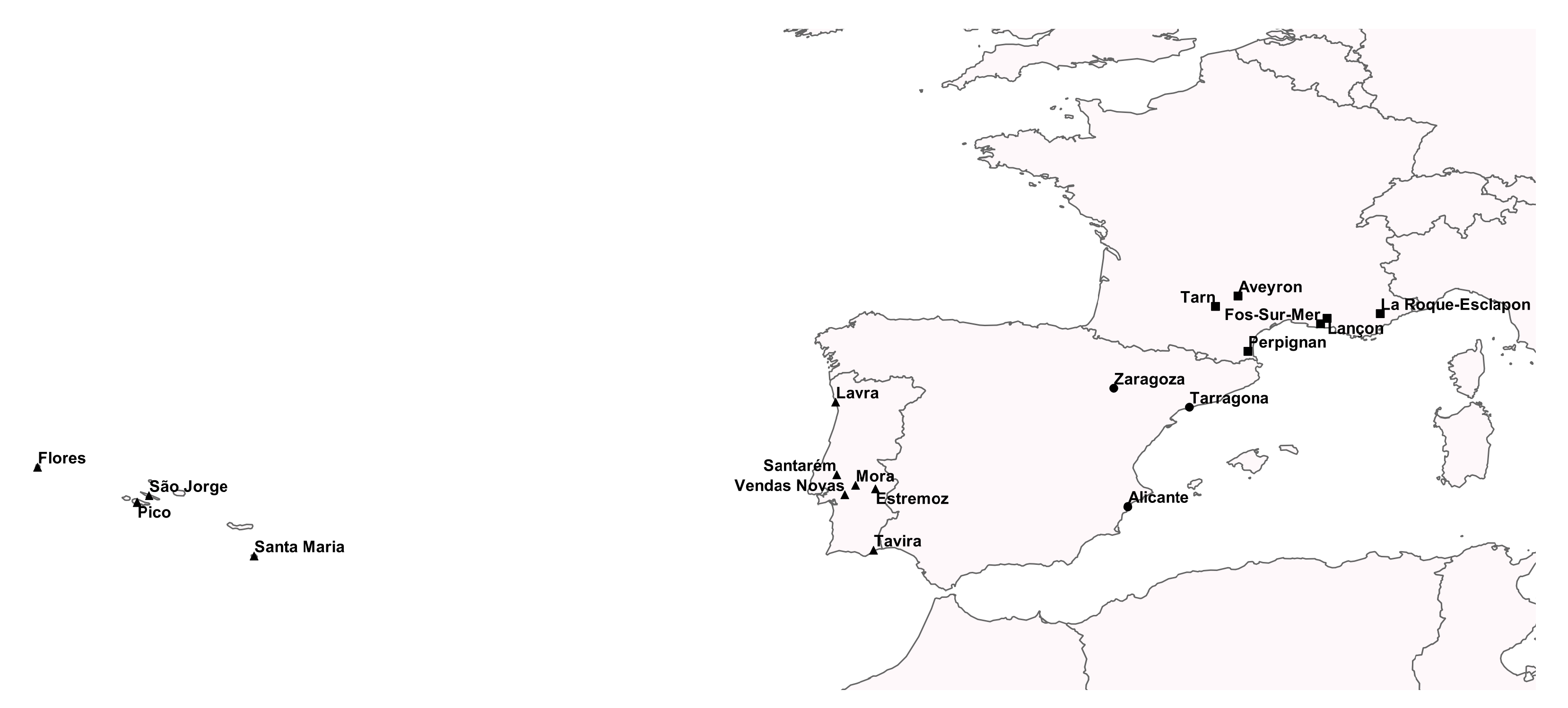
2. Material and Methods
3. Results
3.1. Leporids IGHE and IGHM
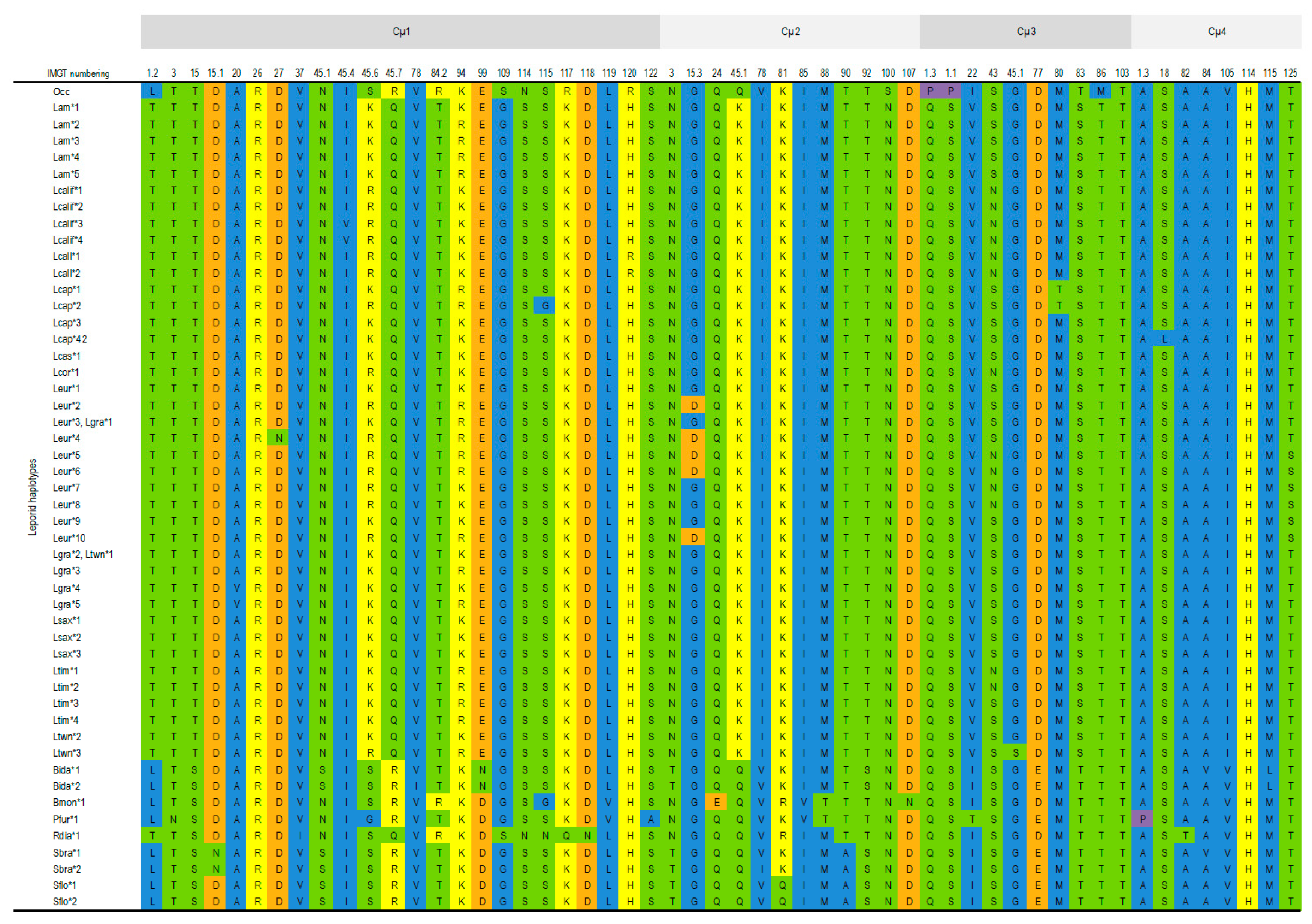
3.1.1. IGHM
Cµ1 Domain
Cµ2 Domain
Cµ3 Domain
Cµ4 domain
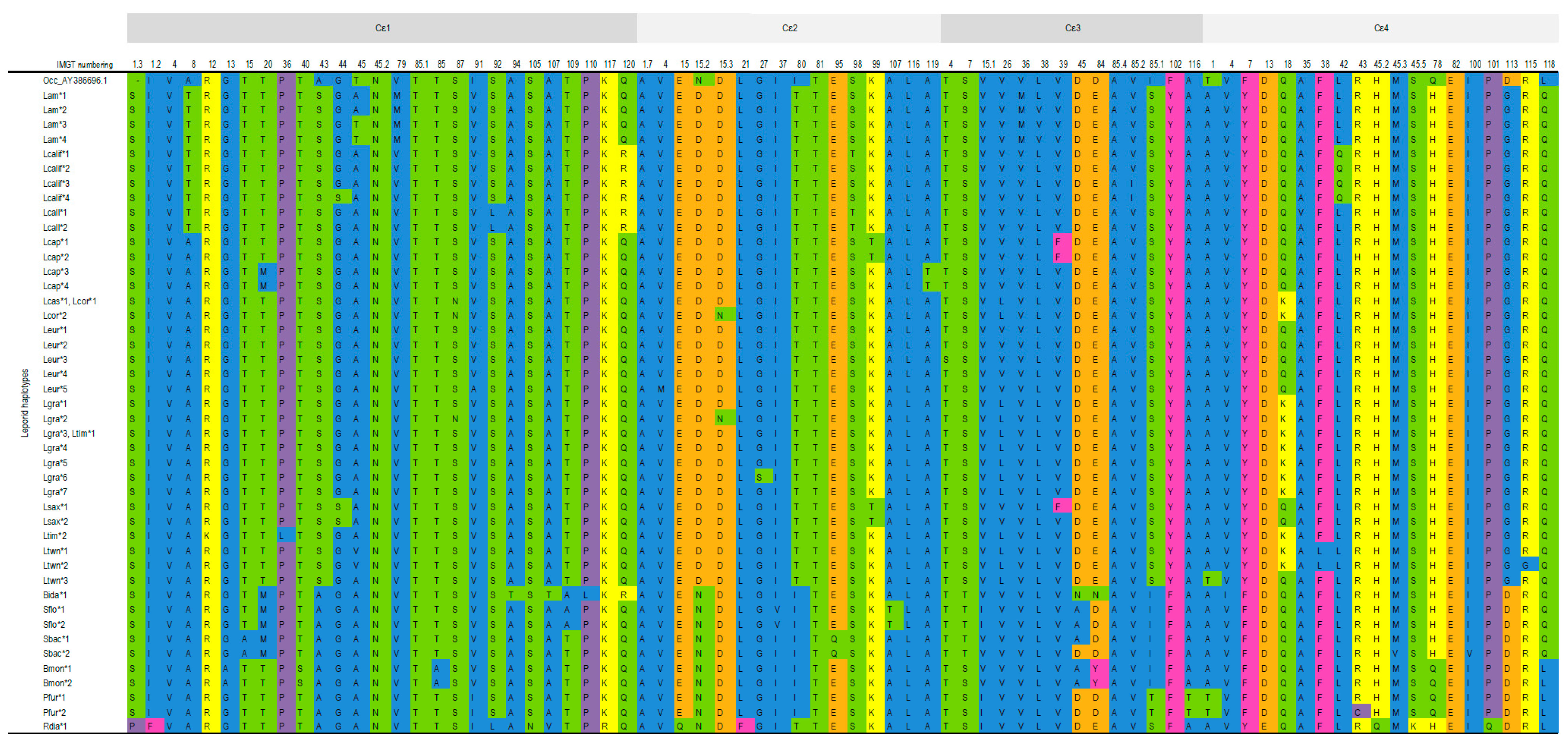
3.1.2. IGHE
Cε1 Domain
Cε2 Domain
Cε3 Domain
Cε4 Domain
3.2. Wild and Domestic European Rabbits IgM and IgE Diversity
3.2.1. IGHM
3.2.2. IGHE
4. Discussion
4.1. IGHM and IGHE Evolution
4.2. Wild and Domestic European Rabbits IgM and IgE Diversity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schroeder, H.W., Jr.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [PubMed]
- Esteves, P.J.; Abrantes, J.; Baldauf, H.M.; BenMohamed, L.; Chen, Y.; Christensen, N.; Gonzalez-Gallego, J.; Giacani, L.; Hu, J.; Kaplan, G.; et al. The wide utility of rabbits as models of human diseases. Exp. Mol. Med. 2018, 50, 66. [Google Scholar] [CrossRef] [PubMed]
- Lanning, D.K.; Zhai, S.K.; Knight, K.L. Analysis of the 3′ Cmu region of the rabbit Ig heavy chain locus. Gene 2003, 309, 135–144. [Google Scholar] [CrossRef]
- Ros, F.; Puels, J.; Reichenberger, N.; van Schooten, W.; Buelow, R.; Platzer, J. Sequence analysis of 0.5 Mb of the rabbit germline immunoglobulin heavy chain locus. Gene 2004, 330, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Gertz, E.M.; Schaffer, A.A.; Agarwala, R.; Bonnet-Garnier, A.; Rogel-Gaillard, C.; Hayes, H.; Mage, R.G. Accuracy and coverage assessment of Oryctolagus cuniculus (rabbit) genes encoding immunoglobulins in the whole genome sequence assembly (OryCun2.0) and localization of the IGH locus to chromosome 20. Immunogenetics 2013, 65, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Burnett, R.C.; Hanly, W.C.; Zhai, S.K.; Knight, K.L. The IgA heavy-chain gene family in rabbit: Cloning and sequence analysis of 13 C alpha genes. EMBO J. 1989, 8, 4041–4047. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.; de Sousa-Pereira, P.; Strive, T.; Knight, K.L.; Woof, J.M.; Esteves, P.J.; Abrantes, J. Identification of a new European rabbit IgA with a serine-rich hinge region. PLoS ONE 2018, 13, e0201567. [Google Scholar] [CrossRef] [PubMed]
- Esteves, P.J.; Alves, P.C.; Ferrand, N.; van der Loo, W. Hotspot variation at the CH2-CH3 interface of leporid IgG antibodies (Oryctolagus, Sylvilagus and Lepus). Eur. J. Immunogenet. 2002, 29, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Girardi, E.; Holdom, M.D.; Davies, A.M.; Sutton, B.J.; Beavil, A.J. The crystal structure of rabbit IgG-Fc. Biochem. J. 2009, 417, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Hamers, R.; Hamers-Casterman, C. Molecular localization of a chain allotypic specificities in rabbit IgG (7S gamma-globulin). J. Mol. Biol. 1965, 14, 288–289. [Google Scholar] [CrossRef]
- Pinheiro, A.; Woof, J.M.; Almeida, T.; Abrantes, J.; Alves, P.C.; Gortazar, C.; Esteves, P.J. Leporid immunoglobulin G shows evidence of strong selective pressure on the hinge and CH3 domains. Open Biol. 2014, 4, 140088. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.A.; Punt, J.; Stranford, S.A.; Jones, P.P.; Kuby, J. Kuby Immunology, 7th ed.; W.H. Freeman: New York, NY, USA, 2013; p. 692. [Google Scholar]
- Lopez-Martinez, N. Revisión Sistemática y Biostratigráfica de los Lagomorpha (Mammalia) del Terciario y Cuatemario de España; Diputación General de Aragón: Zaragoza, Spain, 1989; Volume 3. [Google Scholar]
- Queney, G.; Ferrand, N.; Weiss, S.; Mougel, F.; Monnerot, M. Stationary Distributions of Microsatellite Loci between Divergent Population Groups of the European Rabbit (Oryctolagus cuniculus). Mol. Biol. Evol. 2001, 18, 2169–2178. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.; Afonso, S.; Geraldes, A.; Garreau, H.; Bolet, G.; Boucher, S.; Tircazes, A.; Queney, G.; Nachman, M.W.; Ferrand, N. The genetic structure of domestic rabbits. Mol. Biol. Evol. 2011, 28, 1801–1816. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.; Albert, F.W.; Afonso, S.; Pereira, R.J.; Burbano, H.; Campos, R.; Melo-Ferreira, J.; Blanco-Aguiar, J.A.; Villafuerte, R.; Nachman, M.W.; et al. The genomic architecture of population divergence between subspecies of the European rabbit. PLoS Genet 2014, 10, e1003519. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.C.; Hackländer, K. Lagomorph species: Geographical distribution and conservation status. In Lagomorph Biology; Alves, P.C., Ferrand, N., Hackländer, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 395–405. [Google Scholar]
- Chapman, J.; Flux, J. Introduction and overview of the lagomorphs. In Rabbits, Hares and Pikas, Status, Survey and Conservation Action Plan; Chapman, J., Flux, J., Eds.; UICN/SSC Lagomorph Specialist Group: Gland, Switzerland, 1990; pp. 1–6. [Google Scholar]
- Corbet, G.B.; Hill, J.E. A World List of Mammalian Species; Oxford University Press: Oxford, UK, 1980. [Google Scholar]
- Can, D.N.; Abramov, A.V.; Tikhonov, A.N.; Averianov, A.O. Annamite striped rabbit Nesolagus timminsi in Vietnam. Acta Theriol. 2001, 46, 437–440. [Google Scholar] [CrossRef]
- Chapman, J.A.; Cramer, K.L.; Dippenaar, N.J.; Robinson, T.J. Systematics and Biogeography of the New-England Cottontail, Sylvilagus transitionalis (Bangs, 1895), with the Description of a New Species from the Appalachian Mountains. Proc. Biol. Soc. Wash. 1992, 105, 841–866. [Google Scholar]
- Flux, J.E.C.; Angermann, R. The hares and jackrabbits. In Rabbits, Hares and Pikas: Status Conservation Action Plan; Chapman, J.A., Flux, J.E.C., Eds.; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 1990; pp. 61–94. [Google Scholar]
- Frey, J.K.; Fisher, R.D.; Ruedas, L.A. Identification and restriction of the type locality of the Manzano Mountains cottontail, Sylvilagus cognatus Nelson, 1907 (Mammalia: Lagomorpha: Leporidae). Proc. Biol. Soc. Wash. 1997, 110, 329–331. [Google Scholar]
- Matthee, C.A.; Robinson, T.J. Mitochondrial DNA differentiation among geographical populations of Pronolagus rupestris, Smith’s red rock rabbit (Mammalia: Lagomorpha). Heredity 1996, 76 Pt 5, 514–523. [Google Scholar] [CrossRef]
- Surridge, A.K.; Timmins, R.J.; Hewitt, G.M.; Bell, D.J. Striped rabbits in Southeast Asia. Nature 1999, 400, 726. [Google Scholar] [CrossRef]
- Matthee, C.A.; van Vuuren, B.J.; Bell, D.; Robinson, T.J. A molecular supermatrix of the rabbits and hares (Leporidae) allows for the identification of five intercontinental exchanges during the Miocene. Syst. Biol. 2004, 53, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Sparwel, M.; Doronina, L.; Churakov, G.; Stegemann, A.; Brosius, J.; Robinson, T.J.; Schmitz, J. The Volcano Rabbit in the Phylogenetic Network of Lagomorphs. Genome Biol. Evol. 2019, 11, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.J.; Matthee, C.A. Phylogeny and evolutionary origins of the Leporidae: A review of cytogenetics, molecular analyses and a supermatrix analysis. Mammal Rev. 2005, 35, 231–247. [Google Scholar] [CrossRef]
- Pinheiro, A.; Almeida, T.; Esteves, P.J. Survey of genetic diversity of IgG in wild and domestic rabbits. Int. J. Immunogenet. 2015, 42, 364–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrantes, J.; Areal, H.; Esteves, P.J. Insights into the European rabbit (Oryctolagus cuniculus) innate immune system: Genetic diversity of the toll-like receptor 3 (TLR3) in wild populations and domestic breeds. BMC Genet. 2013, 14, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 3. [Google Scholar]
- Lefranc, M.P.; Pommie, C.; Kaas, Q.; Duprat, E.; Bosc, N.; Guiraudou, D.; Jean, C.; Ruiz, M.; Da Piedade, I.; Rouard, M.; et al. IMGT unique numbering for immunoglobulin and T cell receptor constant domains and Ig superfamily C-like domains. Dev. Comp. Immunol. 2005, 29, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, M.; Donnelly, P. A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, A.; Neves, F.; Lemos de Matos, A.; Abrantes, J.; van der Loo, W.; Mage, R.; Esteves, P.J. An overview of the lagomorph immune system and its genetic diversity. Immunogenetics 2016, 68, 83–107. [Google Scholar] [CrossRef] [PubMed]
- Ghumra, A.; Shi, J.; McIntosh, R.S.; Rasmussen, I.B.; Braathen, R.; Johansen, F.E.; Sandlie, I.; Mongini, P.K.; Areschoug, T.; Lindahl, G.; et al. Structural requirements for the interaction of human IgM and IgA with the human Fcalpha/mu receptor. Eur. J. Immunol. 2009, 39, 1147–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braathen, R.; Sorensen, V.; Brandtzaeg, P.; Sandlie, I.; Johansen, F.E. The carboxyl-terminal domains of IgA and IgM direct isotype-specific polymerization and interaction with the polymeric immunoglobulin receptor. J. Biol. Chem. 2002, 277, 42755–42762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, K.A.; Wang, J.; Urban, B.C.; Czajkowsky, D.M.; Pleass, R.J. Glycan-independent binding and internalization of human IgM to FCMR, its cognate cellular receptor. Sci. Rep. 2017, 7, 42989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, T.H.; Boyle, A.L.; Diebolder, C.A.; Kros, A.; Koster, A.J.; Gros, P. Insights into IgM-mediated complement activation based on in situ structures of IgM-C1-C4b. Proc. Natl. Acad. Sci. USA 2019, 116, 11900–11905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arya, S.; Chen, F.; Spycher, S.; Isenman, D.E.; Shulman, M.J.; Painter, R.H. Mapping of amino acid residues in the C mu 3 domain of mouse IgM important in macromolecular assembly and complement-dependent cytolysis. J. Immunol. 1994, 152, 1206–1212. [Google Scholar] [PubMed]
- Taylor, B.; Wright, J.F.; Arya, S.; Isenman, D.E.; Shulman, M.J.; Painter, R.H. C1q binding properties of monomer and polymer forms of mouse IgM mu-chain variants. Pro544Gly and Pro434Ala. J. Immunol. 1994, 153, 5303–5313. [Google Scholar] [PubMed]
- Holdom, M.D.; Davies, A.M.; Nettleship, J.E.; Bagby, S.C.; Dhaliwal, B.; Girardi, E.; Hunt, J.; Gould, H.J.; Beavil, A.J.; McDonnell, J.M.; et al. Conformational changes in IgE contribute to its uniquely slow dissociation rate from receptor FcepsilonRI. Nat. Struct. Mol. Biol. 2011, 18, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garman, S.C.; Wurzburg, B.A.; Tarchevskaya, S.S.; Kinet, J.P.; Jardetzky, T.S. Structure of the Fc fragment of human IgE bound to its high-affinity receptor FcepsilonRI alpha. Nature 2000, 406, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.; Woof, J.M.; Abi-Rached, L.; Parham, P.; Esteves, P.J. Computational analyses of an evolutionary arms race between mammalian immunity mediated by immunoglobulin A and its subversion by bacterial pathogens. PLoS ONE 2013, 8, e73934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayner, L.E.; Kadkhodayi-Kholghi, N.; Heenan, R.K.; Gor, J.; Dalby, P.A.; Perkins, S.J. The solution structure of rabbit IgG accounts for its interactions with the Fc receptor and complement C1q and its conformational stability. J. Mol. Biol. 2013, 425, 506–523. [Google Scholar] [CrossRef] [PubMed]
- Sondermann, P.; Huber, R.; Oosthuizen, V.; Jacob, U. The 3.2-A crystal structure of the human IgG1 Fc fragment-Fc gammaRIII complex. Nature 2000, 406, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Zacharias, M. Atomic resolution model of the antibody Fc interaction with the complement C1q component. Mol. Immunol. 2012, 51, 66–72. [Google Scholar] [CrossRef] [PubMed]
- van der Loo, W.; Mougel, F.; Sanchez, M.S.; Bouton, C.; Castien, E.; Fonseca, A.; Ferrand, N.; Soriguer, R.; Monnerot, M. Cytonuclear disequilibria in wild populations of rabbit (Oryctolagus cuniculus L.) suggest unequal allele turnover rates at the b locus (IGKC1). Immunogenetics 1999, 49, 629–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteves, P.J.; Lanning, D.; Ferrand, N.; Knight, K.L.; Zhai, S.K.; van der Loo, W. Allelic variation at the VHa locus in natural populations of rabbit (Oryctolagus cuniculus, L.). J. Immunol. 2004, 172, 1044–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surridge, A.K.; van der Loo, W.; Abrantes, J.; Carneiro, M.; Hewitt, G.M.; Esteves, P.J. Diversity and evolutionary history of the MHC DQA gene in leporids. Immunogenetics 2008, 60, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.; Lanning, D.; Alves, P.C.; Mage, R.G.; Knight, K.L.; van der Loo, W.; Esteves, P.J. Molecular bases of genetic diversity and evolution of the immunoglobulin heavy chain variable region (IGHV) gene locus in leporids. Immunogenetics 2011, 63, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilic, T.; Stepanovic, P.; Nenadovic, K.; Dimitrijevic, S. Improving agricultural production of domestic rabbits in Serbia by follow-up study of their parasitic infections. Iran. J. Vet. Res. 2018, 19, 290–297. [Google Scholar] [PubMed]
- Szkucik, K.; Pyz-Lukasik, R.; Szczepaniak, K.O.; Paszkiewicz, W. Occurrence of gastrointestinal parasites in slaughter rabbits. Parasitol. Res. 2014, 113, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Loo, W. Variance analysis of immunoglobulin alleles in natural populations of rabbit (Oryctolagus cuniculus): The extensive interallelic divergence at the b locus could be the outcome of overdominance-type selection. Genetics 1993, 135, 171–187. [Google Scholar] [PubMed]
- Flajnik, M.F.; Kasahara, M. Origin and evolution of the adaptive immune system: Genetic events and selective pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Subspecies | Population | N | S | π | D | H | Hd | Hd Variance | Hd Std Deviation |
|---|---|---|---|---|---|---|---|---|---|---|
| IGHM | All | 45 | 31 | 0.00317 | −0.98878 | 40 | 0.949 | 0.00017 | 0.013 | |
| O. c. algirus | Portugal | 10 | 13 | 0.00283 | 0.10630 | 11 | 0.911 | 0.00176 | 0.042 | |
| O. c. cuniculus | Spain | 10 | 20 | 0.00340 | −0.84715 | 13 | 0.911 | 0.00289 | 0.054 | |
| France | 10 | 15 | 0.00345 | 0.39814 | 13 | 0.958 | 0.00065 | 0.025 | ||
| Domestic | 15 | 11 | 0.00243 | 0.59465 | 8 | 0.747 | 0.00363 | 0.060 | ||
| IGHE | All | 45 | 26 | 0.00226 | −1.38819 | 24 | 0.817 | 0.00078 | 0.028 | |
| O. c. algirus | Portugal | 10 | 16 | 0.00348 | −0.02986 | 11 | 0.868 | 0.0041 | 0.064 | |
| O. c. cuniculus | Spain | 10 | 16 | 0.00222 | −1.37449 | 8 | 0.774 | 0.0051 | 0.071 | |
| France | 10 | 3 | 0.00068 | 0.08868 | 3 | 0.542 | 0.01094 | 0.105 | ||
| Domestic | 15 | 9 | 0.00136 | −0.71769 | 8 | 0.743 | 0.003 | 0.055 |
| Subspecies | Population | Domain, Amino Acid Position and Most Common Triplet and Amino Acid | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cµ1 | Cµ3 | Cµ4 | |||||||||||||
| 26 | 37 | 78 | 109 | 103 | 82 | 114 | |||||||||
| cgg | R | gtc | V | gtc | V | agc | S | acg | T | gcc | A | cac | H | ||
| O. c. algirus | Portugal | … | R | … | V | … | V | ga. | D | …/..a | T | … | A | … | H |
| .a. | Q | a.. | I | … | S | ||||||||||
| g.. | G | ||||||||||||||
| O. c. cuniculus | Spain | … | R | … | V | … | V | ga. | D | …/..a | T | … | A | … | H |
| .a. | Q | a.. | I | … | S | .ta | I | a.. | T | .t. | L | ||||
| France | … | R | … | V | … | V | ga. | D | …/..a | T | … | A | … | H | |
| a.. | I | a.. | I | … | S | ||||||||||
| Domestic | … | R | … | V | … | V | ga. | D | …/..a | T | … | A | |||
| a.. | I | a.. | I | … | S | ||||||||||
| Subspecies | Population | Domain, Amino Acid Position and Most Common Triplet and Amino Acid | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cε1 | Cε2 | Cε3 | Cε4 | ||||||||||||||||||||||
| 4 | 12 | 44 | 45.2 | 84.4 | 1.7 | 81 | 116 | 84 | 85.4 | 1 | 82 | ||||||||||||||
| gtc | V | aga | R | ggc | G | aac | N | acg | T | gcg | A | acg | T | ctc | L | tat | Y | gcc | A | acc | T | gag | E | ||
| O. c. algirus | Portugal | … | V | … | R | … | G | … | N | … | T | … | A | … | T | L | … | Y | … | A | … | T | … | E | |
| .t. | M | t.. | F | g.. | D | g.. | A | ||||||||||||||||||
| O. c. cuniculus | Spain | … | V | … | R | … | G | … | N | … | T | … | A | … | T | … | L | g.. | D | … | A | …. | T | … | E |
| a.. | I | .t. | M | … | Y | g.. | A | a.. | K | ||||||||||||||||
| France | … | V | … | R | … | G | … | N | … | T | … | A | … | T | … | L | g.. | D | … | A | … | T | … | E | |
| g.. | A | ||||||||||||||||||||||||
| Domestic | … | V | … | R | … | G | … | N | … | T | … | A | … | T | … | L | g.. | D | A | … | T | … | E | ||
| .a. | K | .c. | A | .c. | T | .g. | G | .t. | V | g.. | A | ||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, A.; Almeida, T.; Esteves, P.J. Genetic Diversity of IGHM and IGHE in the Leporids Revealed Different Patterns of Diversity in the Two European Rabbit Subspecies (O. cuniculus algirus and O. c. cuniculus). Animals 2019, 9, 955. https://doi.org/10.3390/ani9110955
Pinheiro A, Almeida T, Esteves PJ. Genetic Diversity of IGHM and IGHE in the Leporids Revealed Different Patterns of Diversity in the Two European Rabbit Subspecies (O. cuniculus algirus and O. c. cuniculus). Animals. 2019; 9(11):955. https://doi.org/10.3390/ani9110955
Chicago/Turabian StylePinheiro, Ana, Tereza Almeida, and Pedro J. Esteves. 2019. "Genetic Diversity of IGHM and IGHE in the Leporids Revealed Different Patterns of Diversity in the Two European Rabbit Subspecies (O. cuniculus algirus and O. c. cuniculus)" Animals 9, no. 11: 955. https://doi.org/10.3390/ani9110955
APA StylePinheiro, A., Almeida, T., & Esteves, P. J. (2019). Genetic Diversity of IGHM and IGHE in the Leporids Revealed Different Patterns of Diversity in the Two European Rabbit Subspecies (O. cuniculus algirus and O. c. cuniculus). Animals, 9(11), 955. https://doi.org/10.3390/ani9110955