Age-Dependent Utilization of Shelters and Habitat in Two Reptile Species with Contrasting Intraspecific Interactions


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
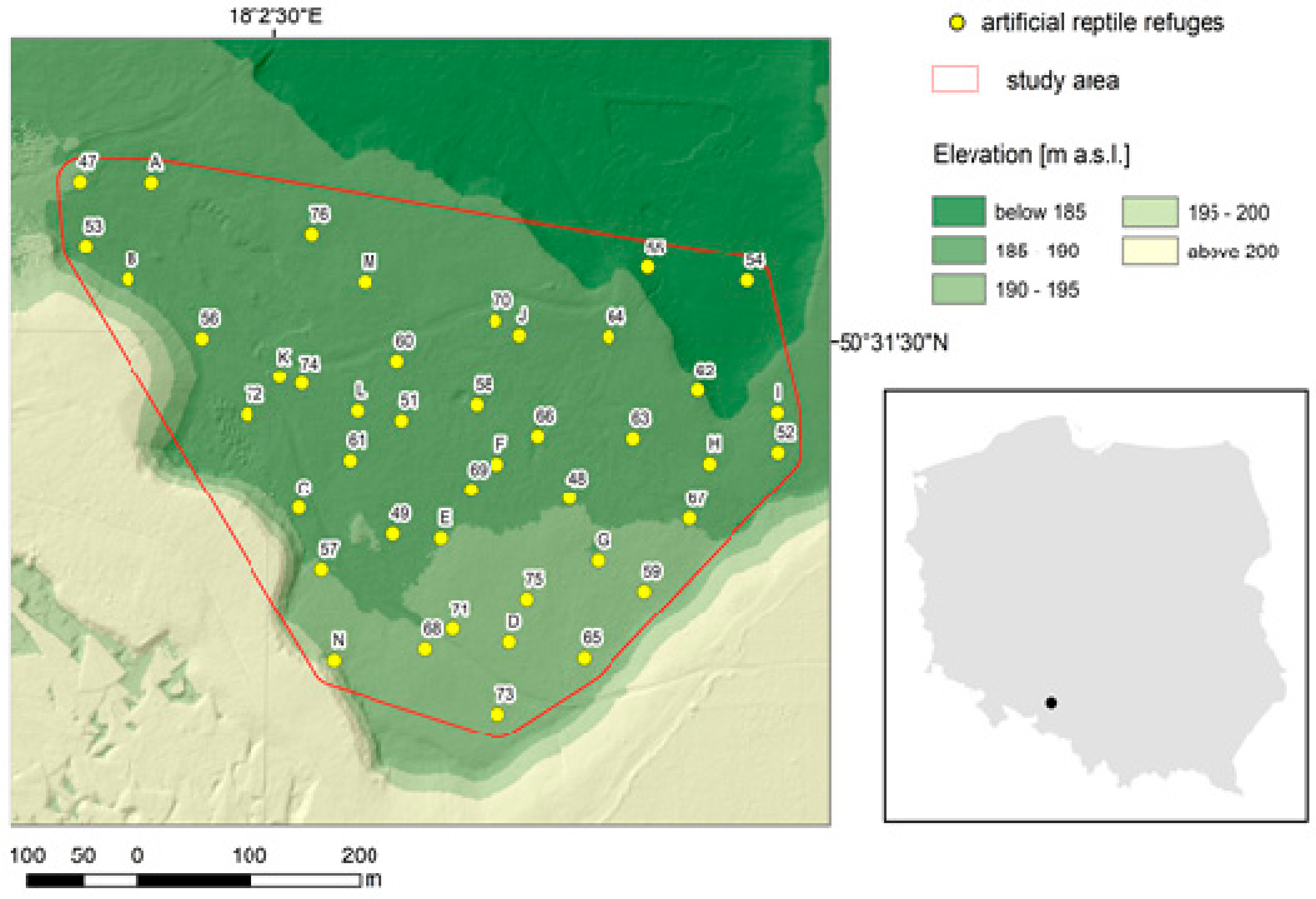
2.1. Study Area
2.2. Experimental Design and Sampling Data
2.3. Data Analysis
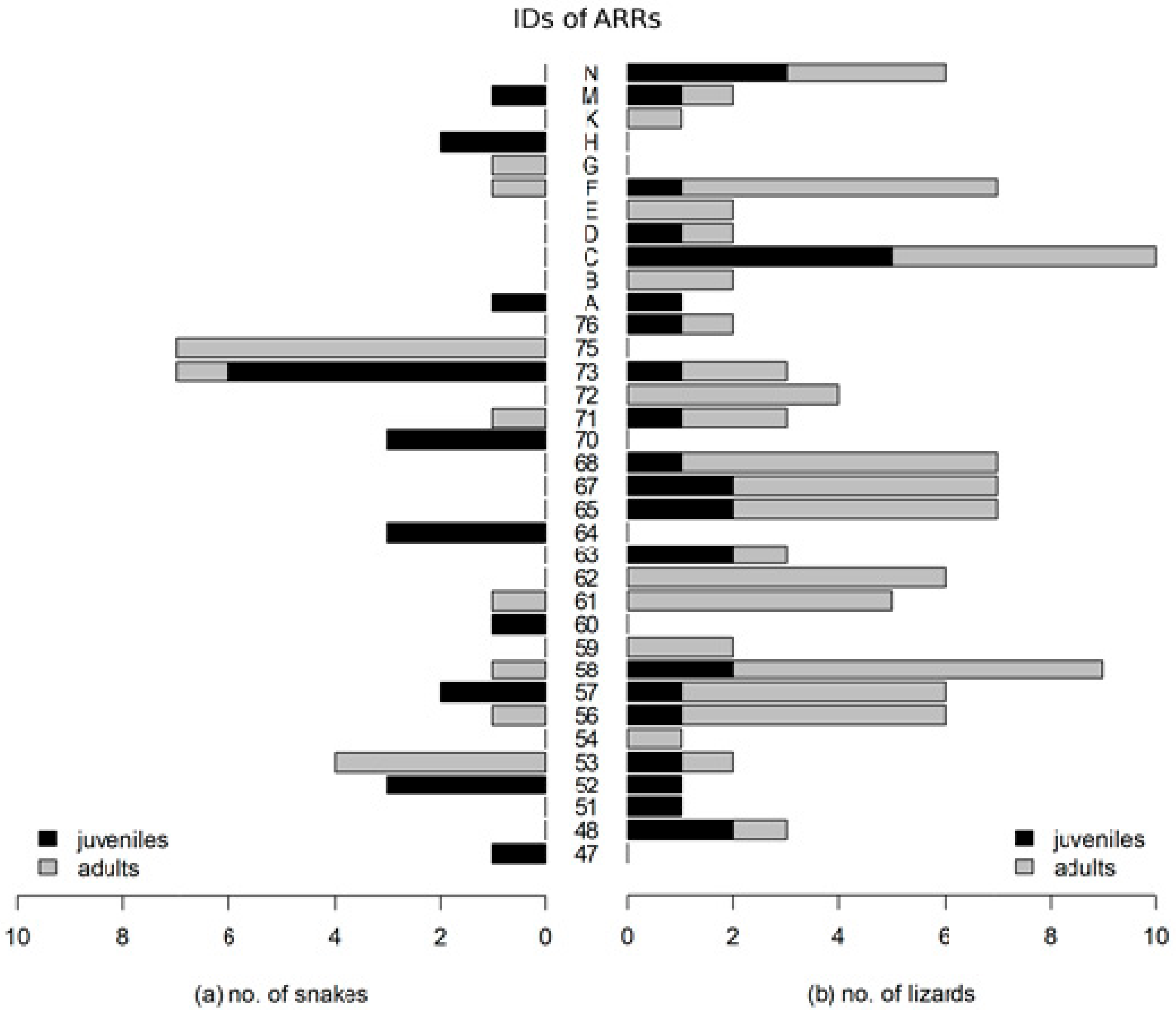
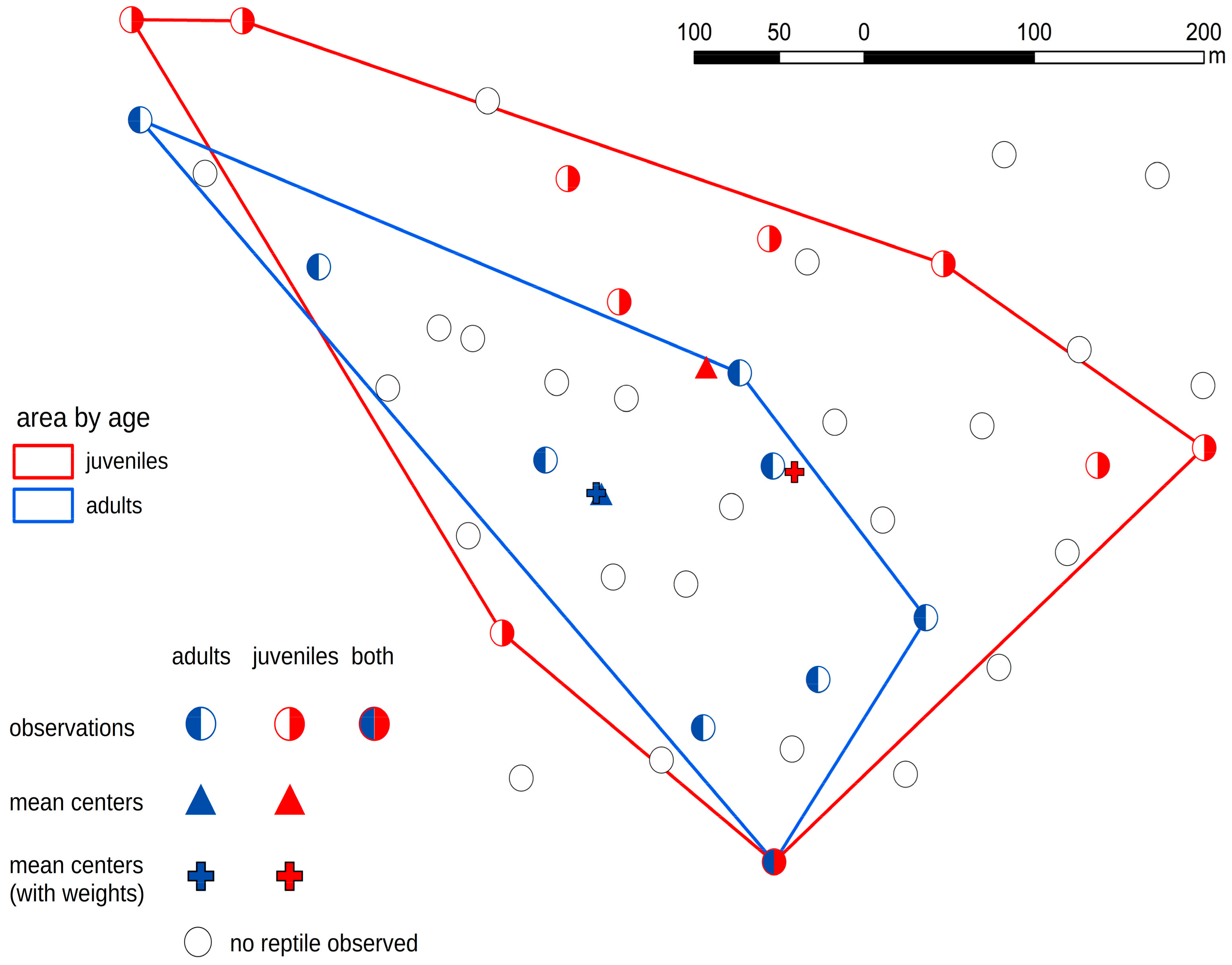
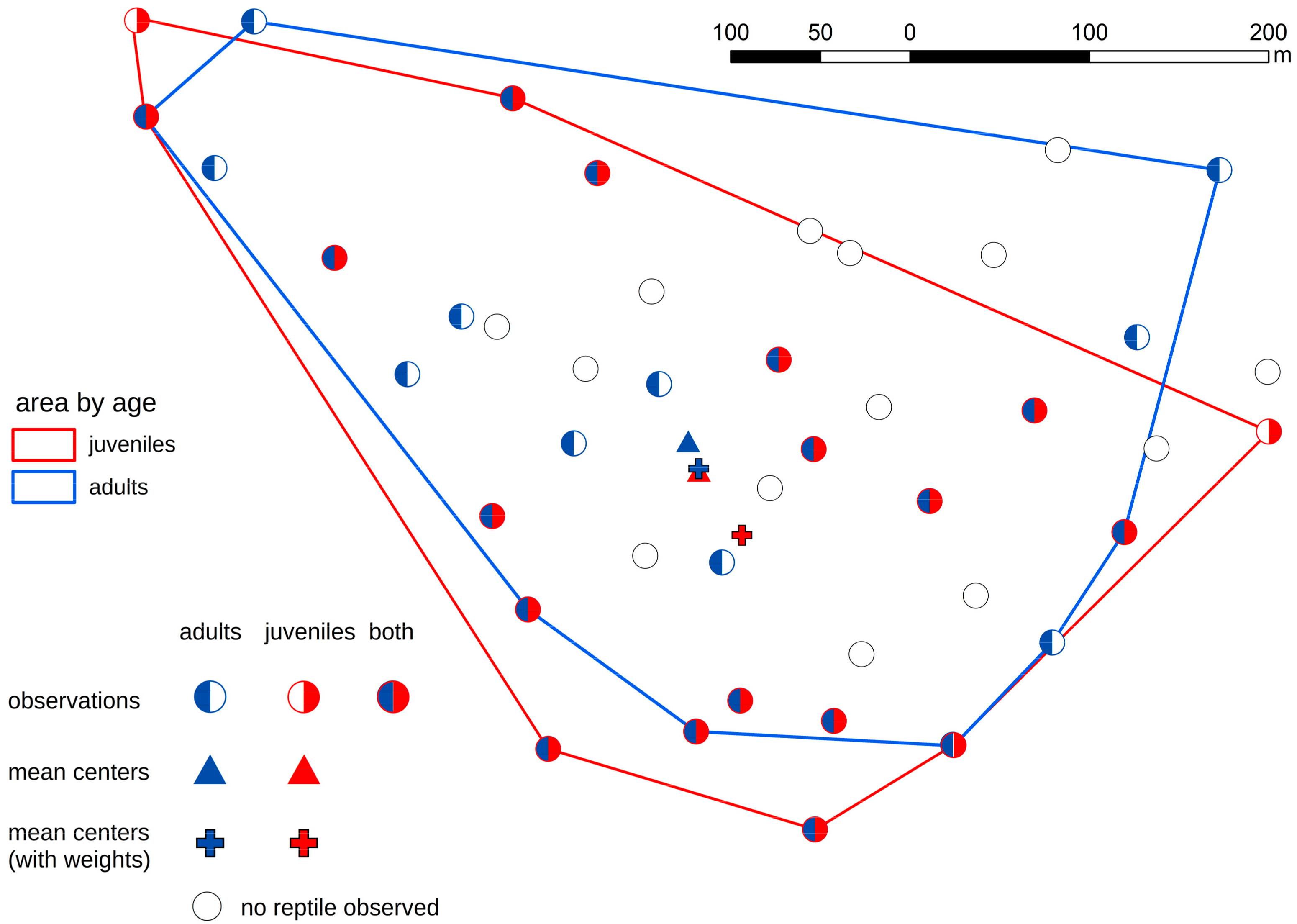
3. Results
4. Discussion
5. Conclusions
- Smooth snakes and slow worms showed different age-dependent spatial distribution.
- In the case of the smooth snake, we discovered that juvenile specimens occupied different artificial shelters from adults. In the case of lizards, there were no such dependencies.
- Juvenile snakes chose sites with suboptimal habitat conditions probably because sites with better habitat condition are occupied by adult snakes.
- Taken together, our results provide strong support for the role of age and behavioral cues in shaping the spatial ecology of terrestrial squamates.
- Our findings may also indicate that habitat-based reptile conservation needs to account for the social mode of a given species and associated spatial structuring of the population.
6. Licenses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brooks, T.M.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.D.; Rylands, A.B.; Konstant, W.R.; Flick, P.; Pilgrim, J.; Oldfield, S.; Magin, G.; et al. Habitat Loss and Extinction in the Hotspots of Biodiversity. Conserv. Biol. 2002, 16, 909–923. [Google Scholar] [CrossRef]
- Law, B.S.; Dickman, C.R. The use of habitat mosaics by terrestrial vertebrate fauna: Implications for conservation and management. Biodivers. Conserv. 1998, 7, 323–333. [Google Scholar] [CrossRef]
- Goulart, F.V.B.; Cáceres, N.C.; Graipel, M.E.; Tortato, M.A.; Ghizoni, I.R.; Oliveira-Santos, L.G.R. Habitat selection by large mammals in a southern Brazilian Atlantic Forest. Mamm. Biol. 2009, 74, 182–190. [Google Scholar] [CrossRef]
- Touihri, M.; Séguy, M.; Imbeau, L.; Mazerolle, M.J.; Bird, D.M. Effects of agricultural lands on habitat selection and breeding success of American kestrels in a boreal context. Agric. Ecosyst. Environ. 2019, 272, 146–154. [Google Scholar] [CrossRef]
- Regosin, J.V.; Windmiller, B.S.; Homan, R.N.; Reed, J.M. Variation in terrestrial habitat use by four pool-breeding amphibian species. J. Wildl. Manag. 2005, 69, 1481–1493. [Google Scholar] [CrossRef]
- Kurek, K.; Król, W.; Najberek, K.; Ćmiel, A.M.; Solarz, W.; Bury, S.; Baś, G.; Najbar, B.; Okarma, H. Habitat use of the Aesculapian snake at different spatial scales. J. Wildl. Manag. 2018, 82, 1746–1755. [Google Scholar] [CrossRef]
- Shine, R.; Wall, M. Why is intraspecific niche partitioning more common in snakes than in lizards? In Lizard Ecology; Reilly, S.M., McBrayer, L.B., Miles, D.B., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 173–208. [Google Scholar] [CrossRef]
- Peterson, C.R. Snake thermal ecology: The causes and consequences of body-temperature variation. In Snakes: Ecology and Behavior; McGraw-Hill: New York, NY, USA, 1993; pp. 241–314. [Google Scholar]
- Huang, W.-S.; Greene, H.W.; Chang, T.-J.; Shine, R. Territorial behavior in Taiwanese kukrisnakes (Oligodon formosanus). Proc. Natl. Acad. Sci. USA 2011, 108, 7455–7459. [Google Scholar] [CrossRef]
- Schuett, G.W.; Clark, R.W.; Repp, R.A.; Amarello, M.; Smith, C.F.; Greene, H.W. Social behavior of rattlesnakes: A shifting paradigm. Ratt. Ariz. Conserv. Behav. Venom Evol. 2016, 2, 161–244. [Google Scholar]
- Shine, R.; Harlow, P.S.; Keogh, J.S. The influence of sex and body size on food habits of a giant tropical snake, Python reticulatus. Funct. Ecol. 1998, 12, 248–258. [Google Scholar] [CrossRef]
- Camilleri, C.; Shine, R. Sexual Dimorphism and Dietary Divergence: Differences in Trophic Morphology between Male and Female Snakes. Copeia 1990, 1990, 649–658. [Google Scholar] [CrossRef]
- Luiselli, L.; Angelici, F.M. Sexual size dimorphism and natural history traits are correlated with intersexual dietary divergence in royal pythons (Python regius) from the rainforests of southeastern Nigeria. Ital. J. Zool. 1998, 65, 183–185. [Google Scholar] [CrossRef]
- Shine, R.; Shine, T.; Shine, B. Intraspecific habitat partitioning by the sea snake Emydocephalus annulatus (Serpentes, Hydrophiidae): The effects of sex, body size, and colour pattern. Biol. J. Linn. Soc. 2003, 80, 1–10. [Google Scholar] [CrossRef]
- Brown, W.S.; MacLean, F.M. Conspecific Scent-Trailing by Newborn Timber Rattlesnakes, Crotalus horridus. Herpetologica 1983, 39, 430–436. [Google Scholar]
- Keren-Rotem, T.; Bouskila, A.; Geffen, E. Ontogenetic habitat shift and risk of cannibalism in the common chameleon (Chamaeleo chamaeleon). Behav. Ecol. Sociobiol. 2006, 59, 723–731. [Google Scholar] [CrossRef]
- Drobenkov, S.M. Distribution, ecological traits and conservation of the smooth snake (Coronella austriaca) in Belarus. Acta Biol. Univ. Daugavp. 2014, 14, 21–27. [Google Scholar]
- Pernetta, A.P.; Reading, C.J.; Allen, J.A. Chemoreception and kin discrimination by neonate smooth snakes, Coronella austriaca. Anim. Behav. 2009, 77, 363–368. [Google Scholar] [CrossRef]
- Gonzalo, A.; Cabido, C.; Martín, J.; López, P. Detection and Discrimination of Conspecific Scents by the Anguid Slow-Worm Anguis fragilis. J. Chem. Ecol. 2004, 30, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.; Lee, J. Statistical Analysis of Geographic Information; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Santos, X.; Brito, J.C.; Sillero, N.; Pleguezuelos, J.M.; Llorente, G.A.; Fahd, S.; Parellada, X. Inferring habitat-suitability areas with ecological modelling techniques and GIS: A contribution to assess the conservation status of Vipera latastei. Biol. Conserv. 2006, 130, 416–425. [Google Scholar] [CrossRef]
- Franklin, J.; Wejnert, K.E.; Hathaway, S.A.; Rochester, C.J.; Fisher, R.N. Effect of species rarity on the accuracy of species distribution models for reptiles and amphibians in Southern California. Divers. Distrib. 2009, 15, 167–177. [Google Scholar] [CrossRef]
- Santos, X.; Brito, J.C.; Caro, J.; Abril, A.J.; Lorenzo, M.; Sillero, N.; Pleguezuelos, J.M. Habitat suitability, threats and conservation of isolated populations of the smooth snake (Coronella austriaca) in the southern Iberian Peninsula. Biol. Conserv. 2009, 142, 344–352. [Google Scholar] [CrossRef]
- Sillero, N.; Brito, J.C.; Skidmore, A.K.; Toxopeus, A.G. Biogeographical patterns derived from remote sensing variables: The amphibians and reptiles of the Iberian Peninsula. Amphib. Reptil. 2009, 30, 185–206. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Phelps, T.E. Biology, general ecology and behaviour of the snake, Coronella austriaca Laurenti. Biol. J. Linn. Soc. 1977, 9, 133–164. [Google Scholar] [CrossRef]
- Gent, A.H. Movement and Dispersion of the Smooth Snake Coronella austriaca Laurenti in Relation to Habitat. Ph.D. Thesis, University of Southampton, Southampton, UK, 1990. [Google Scholar]
- Meteorological Service with Climatic Data. Available online: http://pogodynka.pl/ (accessed on 2 December 2018).
- Juszczyk, W. Płazy i Gady Krajowe. Wyd. II; Polskie Wydawnictwo Naukowe: Warszawa, Poland, 1987. [Google Scholar]
- Brown, W.S.; Parker, W.S. A Ventral Scale Clipping System for Permanently Marking Snakes (Reptilia, Serpentes). J. Herpetol. 1976, 10, 247–249. [Google Scholar] [CrossRef]
- Ekner, A.; Sajkowska, Z.; Dudek, K.; Tryjanowski, P. Medical cautery units as a permanent and non-invasive method of marking lizards. Acta Herpetol. 2011, 6, 229–236. [Google Scholar] [CrossRef]
- Ciofi, C.; Puswati, J.; Winana, D.; Boer, M.E.D.; Chelazzi, G.; Sastrawan, P. Preliminary Analysis of Home Range Structure in the Komodo Monitor, Varanus komodoensis. Copeia 2007, 2007, 462–470. [Google Scholar] [CrossRef]
- Boyle, S.A.; Lourenço, W.C.; Silva, L.R.D.; Smith, A.T. Home Range Estimates Vary with Sample Size and Methods. Folia Primatol. 2009, 80, 33–42. [Google Scholar] [CrossRef]
- Van Beest, F.M.; Rivrud, I.M.; Loe, L.E.; Milner, J.M.; Mysterud, A. What determines variation in home range size across spatiotemporal scales in a large browsing herbivore? J. Anim. Ecol. 2001, 80, 771–785. [Google Scholar] [CrossRef]
- Row, J.R.; Blouin-Demers, G. Thermal quality influences habitat selection at multiple spatial scales in milksnakes. Écoscience 2006, 13, 443–450. [Google Scholar] [CrossRef]
- Burgman, M.A.; Fox, J.C. Bias in species range estimates from minimum convex polygons: Implications for conservation and options for improved planning. Anim. Conserv. 2003, 6, 19–28. [Google Scholar] [CrossRef]
- Nilsen, E.B.; Pedersen, S.; Linnell, J.D.C. Can minimum convex polygon home ranges be used to draw biologically meaningful conclusions? Ecol. Res. 2008, 23, 635–639. [Google Scholar] [CrossRef]
- Row, J.R.; Blouin-Demers, G. Kernels Are Not Accurate Estimators of Home-Range Size for Herpetofauna. Copeia 2006, 2006, 797–802. [Google Scholar] [CrossRef]
- Mitchell, A. The ESRI Guide to GIS Analysis. Volume 2: Spatial Measurements & Statistics; ESRI Press: Redlands, CA, USA, 2005. [Google Scholar]
- Mayhew, S. A Dictionary of Geography; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Documentation of ArcGIS Desktop. Available online: http://desktop.arcgis.com/en/documentation/ (accessed on 31 May 2019).
- Fu, P.; Rich, P.M. A geometric solar radiation model with applications in agriculture and forestry. Comput. Electron. Agric. 2002, 37, 25–35. [Google Scholar] [CrossRef]
- Gent, A.H.; Spellerberg, I.F. Movement rates of the smooth snake Coronella austriaca (Colubridae) A radio telemetric study. Herpetol. J. 1993, 3, 140–146. [Google Scholar]
- Hosmer, D.W., Jr.; Lemeshow, S.; Sturdivant, R.X. Applied Logistic Regression; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 227–242. ISBN 978-1-118-54838-7. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute. ArcGIS Desktop: Release 10.2; Environmental Systems Research Institute: Redlands, CA, USA, 2013. [Google Scholar]
- QGIS. Geographic Information System: Release 3.0. Open Source Geospatial Foundation Project. Technical documentation. Available online: https://www.qgis.org/en/docs/index.html (accessed on 1 August 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 31 May 2019).
- Technical documentation of Python Language, version 2.7. Available online: https://docs.python.org/2/reference/index.html (accessed on 01 May 2019).
- Shine, R.; Webb, J.K.; Fitzgerald, M.; Sumner, J. The impact of bush-rock removal on an endangered snake species, Hoplocephalus bungaroides (Serpentes: Elapidae). Wildl. Res. 1998, 25, 285–295. [Google Scholar] [CrossRef]
- Visagie, L.; Mouton, P.; Bauwens, D. Experimental analysis of grouping behaviour in cordylid lizards. Herpetol. J. 2005, 15, 91–96. [Google Scholar]
- Alfermann, D.; Bohme, W. Populationsstruktur und Raumnutzung der Schlingnatter auf Freileitungstrassen in Wäldern Freilandökologische Untersuchungen unter Zuhilfenahme künstlicher Verstecke (KV) und der Radiotelemetrie. Z. Feldherpetol. 2009, 15, 373–392. [Google Scholar]
- Madsen, T. Movements, Home Range Size and Habitat Use of Radio-Tracked Grass Snakes (Natrix natrix) in Southern Sweden. Copeia 1984, 1984, 707–713. [Google Scholar] [CrossRef]
- Shine, R. Intraspecific Variation in Thermoregulation, Movements and Habitat Use by Australian Blacksnakes, Pseudechis porphyriacus (Elapidae). J. Herpetol. 1987, 21, 165–177. [Google Scholar] [CrossRef]
- Goddard, P. Limited movements areas and spatial behaviour in the smooth snake in southern England. In Proceedings of the European Herpetological Symposium CWLP Oxford; Coborn, J., Ed.; Cotswold Wild Life Park: Burford, UK, 1981; pp. 25–40. [Google Scholar]
- Smith, N.D. The Ecology of the Slow-Worm (Anguis fragilis L.) in Southern England. Ph.D. Thesis, University of Southampton, Southampton, UK, 1990. [Google Scholar]
- Reinert, H.K. Habitat Selection in Snakes. In Snakes: Ecology and Behavior; Seigel, R.A., Collins, J.T., Eds.; McGraw-Hill: New York, NY, USA, 1993; pp. 201–240. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Character of the Distribution | ANN Ratio (p-Value) |
|---|---|---|
| All snakes | dispersed | 1.735 (<0.01) |
| Adult unique snakes | dispersed | 1.440 (<0.05) |
| Juvenile unique snakes | dispersed | 4.305 (<0.01) |
| All lizards | clustered | 0.338 (<0.01) |
| Adult unique lizards | clustered | 0.687 (<0.01) |
| Juvenile unique lizards | random | 0.824 (>0.1) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolanek, A.; Bury, S.; Turniak, E.; Szymanowski, M. Age-Dependent Utilization of Shelters and Habitat in Two Reptile Species with Contrasting Intraspecific Interactions. Animals 2019, 9, 995. https://doi.org/10.3390/ani9110995
Kolanek A, Bury S, Turniak E, Szymanowski M. Age-Dependent Utilization of Shelters and Habitat in Two Reptile Species with Contrasting Intraspecific Interactions. Animals. 2019; 9(11):995. https://doi.org/10.3390/ani9110995
Chicago/Turabian StyleKolanek, Aleksandra, Stanisław Bury, Edyta Turniak, and Mariusz Szymanowski. 2019. "Age-Dependent Utilization of Shelters and Habitat in Two Reptile Species with Contrasting Intraspecific Interactions" Animals 9, no. 11: 995. https://doi.org/10.3390/ani9110995
APA StyleKolanek, A., Bury, S., Turniak, E., & Szymanowski, M. (2019). Age-Dependent Utilization of Shelters and Habitat in Two Reptile Species with Contrasting Intraspecific Interactions. Animals, 9(11), 995. https://doi.org/10.3390/ani9110995