Effect of Dietary Supplementation of Lactobacillus Casei YYL3 and L. Plantarum YYL5 on Growth, Immune Response and Intestinal Microbiota in Channel Catfish

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Probiotic Strains and Culture Conditions
2.3. Experimental Diets
2.4. Fish and Rearing Conditions
2.5. Non-Specific Immune Parameters of Serum
2.6. Challenge Test
2.7. Sample Collection and DNA Extraction
2.8. Bioinformatic Analysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
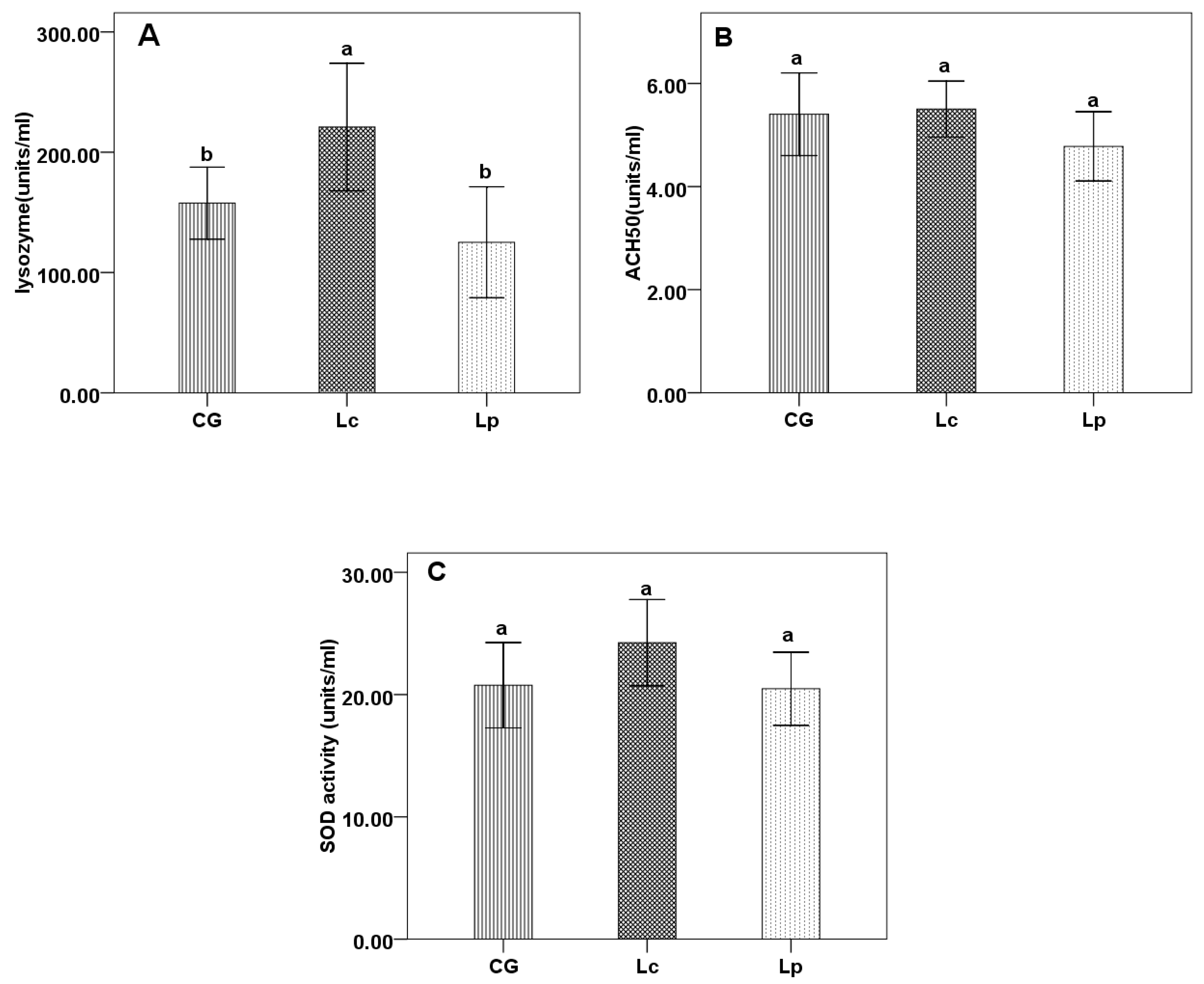
3.2. Immune Parameters
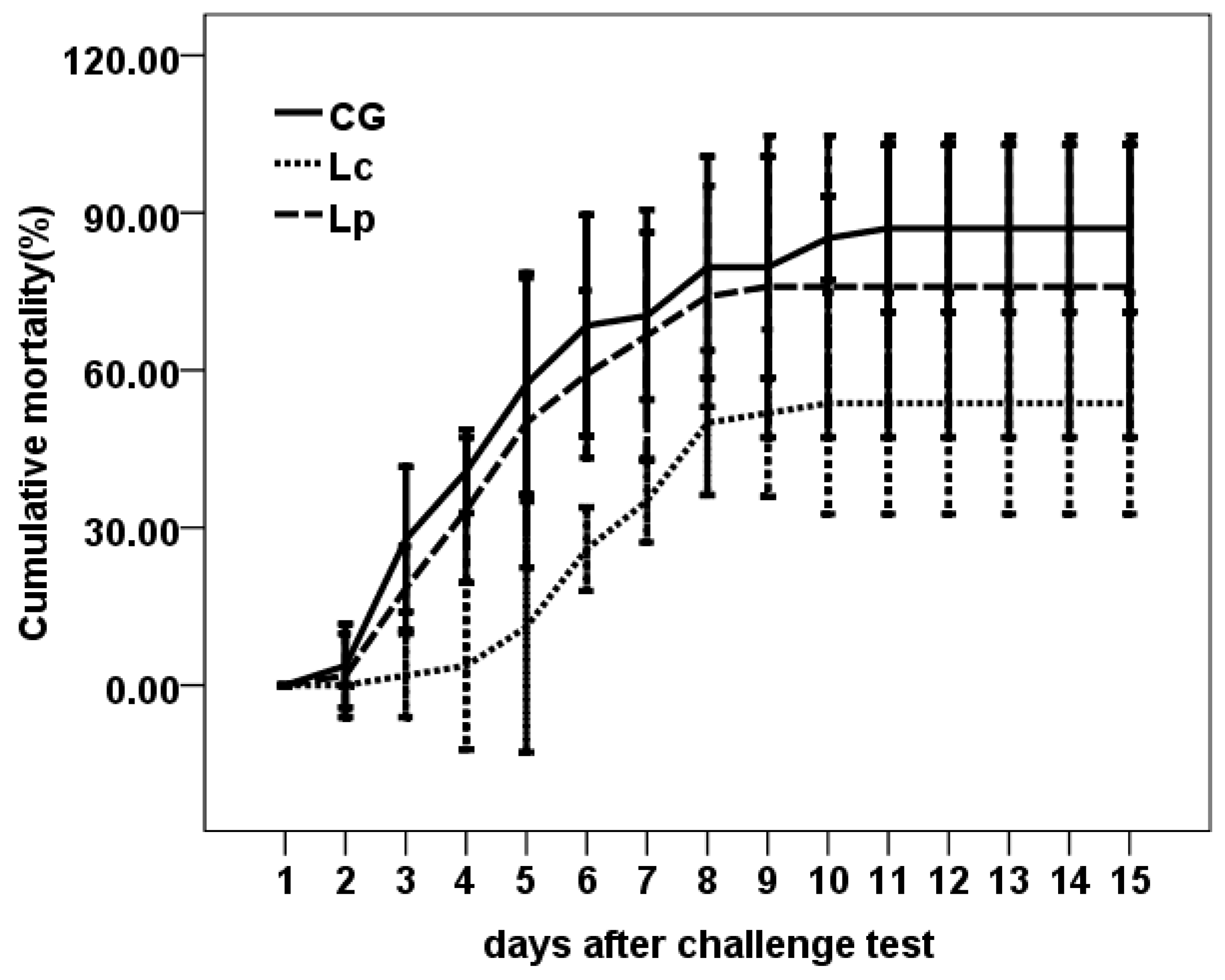
3.3. Challenge Test
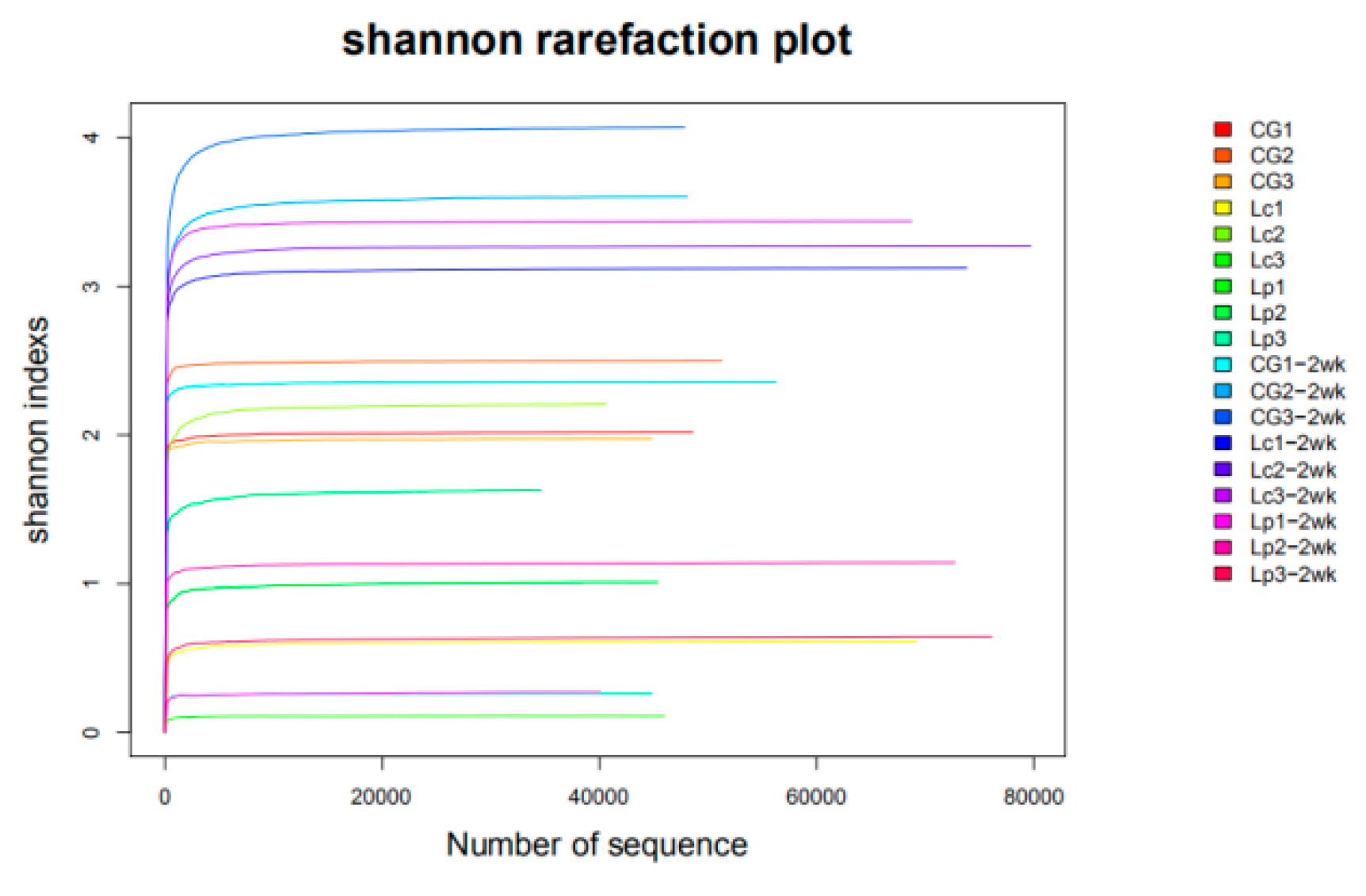
3.4. Characteristics of the High-Throughput Sequence Data
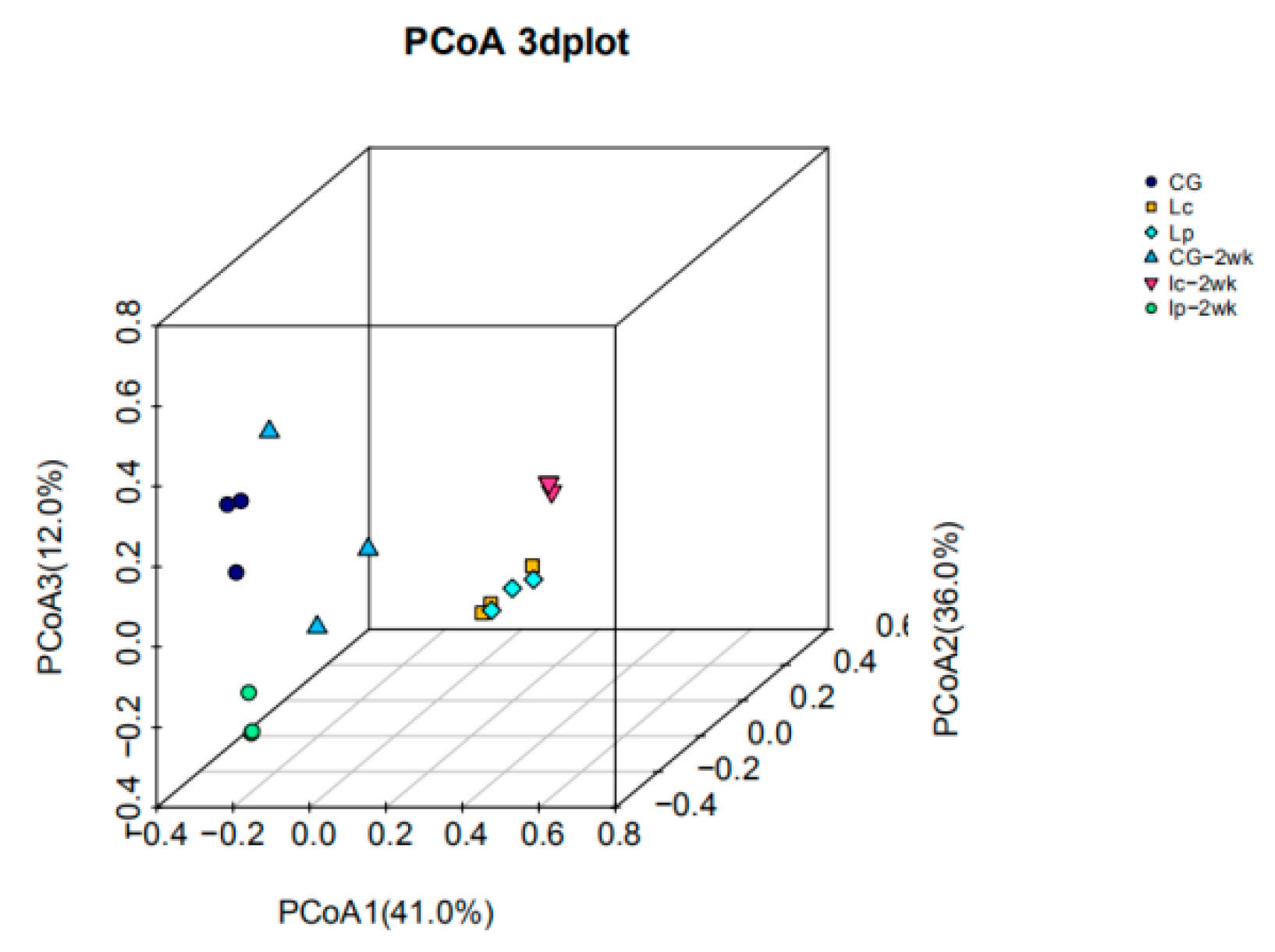
3.5. Diversity Analysis
3.6. Changes in Community Structure and Intestinal Microbiota Abundance in the Catfish
3.7. Functional Analysis of the Intestinal Microflora
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Burgos, F.A.; Ray, C.L.; Arias, C.R. Bacterial diversity and community structure of the intestinal microbiome of Channel Catfish (Ictalurus punctatus) during ontogenesis. Syst. Appl. Microbiol. 2018, 41, 494–505. [Google Scholar] [CrossRef] [PubMed]
- China, B.O.F.I. Chinese Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2018. [Google Scholar]
- Wang, R.H.; Xiao, T.Y.; Zeng, L.B.; Liu, X.Y.; Zhou, Y.; Ma, J. Generation and use of Edwardsiella ictaluri ghosts as a vaccine against enteric septicemia of catfish (ESC). Aquaculture 2016, 456, 9–15. [Google Scholar] [CrossRef]
- Peatman, E.; Mohammed, H.; Kirby, A.; Shoemaker, C.A.; Yildirim-Aksoy, M.; Beck, B.H. Mechanisms of pathogen virulence and host susceptibility in virulent Aeromonas hydrophila infections of channel catfish (Ictalurus punctatus). Aquaculture 2018, 482, 1–8. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S. Recent advances in the role of probiotics and prebiotics in carp aquaculture: A review. Aquaculture 2016, 454, 243–251. [Google Scholar] [CrossRef]
- Zorriehzahra, M.J.; Delshad, S.T.; Adel, M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: An update on their multiple modes of action: A review. Vet. Q. 2016, 36, 228–241. [Google Scholar] [CrossRef]
- Bernardeau, M.; Guguen, M.; Vernoux, J.P. Beneficial lactobacilli in food and feed: Long-term use, biodiversity and proposals for specific and realistic safety assessments. FEMS Microbiol. Rev. 2006, 30, 487–513. [Google Scholar] [CrossRef]
- Ringø, E.; Gatesoupe, F. Lactic acid bacteria in fish: A review. Aquaculture 1998, 160, 177–203. [Google Scholar] [CrossRef]
- Ren, P.F.; Xu, L.; Yang, Y.L.; He, S.X.; Liu, W.S.; Ringø, E.; Zhou, Z.G. Lactobacillus planarum subsp. plantarum JCM 1149 vs. Aeromonas hydrophila NJ-1 in the anterior intestine and posterior intestine of hybrid tilapia Oreochromis niloticus♀ × Oreochromis aureus♂: An ex vivo study. Fish Shellfish Immun. 2013, 35, 146–153. [Google Scholar]
- Gao, Q.X.; Gao, Q.; Min, M.H.; Zhang, C.J.; Peng, S.M.; Shi, Z.H. Ability of Lactobacillus plantarum lipoteichoic acid to inhibit Vibrio anguillarum-induced inflammation and apoptosis in silvery pomfret (Pampus argenteus) intestinal epithelial cells. Fish Shellfish Immun. 2016, 54, 573–579. [Google Scholar] [CrossRef]
- Yu, L.L.; Zhai, Q.X.; Zhu, J.M.; Zhang, C.C.; Li, T.Q.; Liu, X.M.; Zhao, J.X.; Zhang, H.; Tian, F.W.; Chen, W. Dietary Lactobacillus plantarum supplementation enhances growth performance and alleviates aluminum toxicity in tilapia. Ecotoxicol. Environ.Saf. 2017, 143, 307–314. [Google Scholar] [CrossRef]
- Zheng, X.T.; Duan, Y.F.; Dong, H.B.; Zhang, J.S. Effects of Dietary Lactobacillus plantarum on Growth Performance, Digestive Enzymes and Gut Morphology of Litopenaeus vannamei. Probiotics Antimicro 2018, 10, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Effects of heat killed Lactobacillus plantarum (LP20) supplemental diets on growth performance, stress resistance and immune response of red sea bream, Pagrus major. Aquaculture 2015, 442, 29–36. [Google Scholar] [CrossRef]
- Son, V.M.; Chang, C.C.; Wu, M.C.; Guu, Y.K.; Chiu, C.H.; Cheng, W.T. Dietary administration of the probiotic, Lactobacillus plantarum, enhanced the growth, innate immune responses, and disease resistance of the grouper Epinephelus coioides. Fish Shellfish Immun. 2009, 26, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.R.; Song, J.H.; Park, B.S.; Kim, D.; Kwak, J.H.; Do, H.K.; Kim, A.R.; Kim, W.J.; Song, S.K. Distinct immune tones are established by Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 in the gut of olive flounder (Paralichthys olivaceus). Fish Shellfish Immun. 2016, 55, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.S.; Sukumaran, V.; Oviya, M. Potential probiotic Lactobacillus plantarum VSG3 improves the growth, immunity, and disease resistance of tropical freshwater fish, Labeorohita. Fish Shellfish Immun. 2013, 34, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Aktas, B.; Wolfe, T.J.D.; Safdar, N.; Darien, B.J.J.; Steele, J.L. The Impact of Lactobacillus casei on the Composition of the Cecal Microbiota and Innate Immune System Is Strain Specific. PLoS ONE 2016, 11, e156374. [Google Scholar] [CrossRef]
- Lamari, F.; Castex, M.; Larcher, T.; Ledevin, M.; Mazurais, D.; Bakhrouf, A.; Gatesoupe, F.J. Comparison of the effects of the dietary addition of two lactic acid bacteria on the development and conformation of sea bass larvae, Dicentrarchus labrax, and the influence on associated microbiota. Aquaculture 2013, 376–379, 137–145. [Google Scholar] [CrossRef]
- Abasali, H.; Mohamad, S. Effect of Dietary Supplementation with Probiotic on Reproductive Performance of Female Livebearing Ornamental Fish. Res. J. Anim. Sci. 2010, 4, 103–107. [Google Scholar]
- Zhang, D.D.; Beck, B.H.; Lange, M.D.; Zhao, H.G.; Thongda, W.; Ye, Z.; Li, C.; Peatman, E. Impact of oral and waterborne administration of rhamnolipids on the susceptibility of channel catfish (Ictalurus punctatus) to Flavobacterium columnare infection. Fish Shellfish Immun. 2017, 60, 44–49. [Google Scholar] [CrossRef]
- Li, E.C.; Lim, C.; Klesius, P. Enhancement Effects of Dietary Wheat Distiller’s Dried Grains with Solubles on Growth, Immunity, and Resistance to Edwardsiella ictaluri Challenge of Channel Catfish, Ictalurus punctatus. J. WorldAquac. Soc. 2012, 43, 814–827. [Google Scholar]
- Liu, C.H.; Wu, K.C.; Chu, T.W.; Wu, T.M. Dietary supplementation of probiotic, Bacillus subtilis E20, enhances the growth performance and disease resistance against Vibrio alginolyticus in parrot fish (Oplegnathus fasciatus). Aquacult. Int. 2018, 26, 63–74. [Google Scholar] [CrossRef]
- Xia, Y.; Lu, M.X.; Chen, G.; Cao, J.M.; Gao, F.Y.; Wang, M.; Liu, Z.G.; Zhang, D.F.; Zhu, H.P.; Yi, M.M. Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia, Oreochromis niloticus. Fish Shellfish Immun. 2018, 76, 368–379. [Google Scholar]
- Gupta, A.; Sehgal, H.S.; Sehgal, G.K. Growth and carcass composition of giant freshwater prawn, Macrobrachium rosenbergii (De Man), fed different isonitrogenous and isocaloric diets. Aquac. Res. 2007, 38, 1355–1363. [Google Scholar] [CrossRef]
- Doan, H.V.; Hoseinifar, S.H.; Tapingkae, W.; Tongsiri, S.; Khamtavee, P. Combined administration of low molecular weight sodium alginate boosted immunomodulatory, disease resistance and growth enhancing effects of Lactobacillus plantarum in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immun. 2016, 58, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Doan, H.V.; Doolgindachbaporn, S.; Suksri, A. Effects of low molecular weight agar and Lactobacillus plantarum on growth performance, immunity, and disease resistance of basa fish (Pangasius bocourti, Sauvage 1880). Fish Shellfish Immun. 2014, 41, 340–345. [Google Scholar] [CrossRef]
- Liu, H.T.; Wang, S.F.; Cai, Y.; Guo, X.H.; Cao, Z.J.; Zhang, Y.Z.; Liu, S.B.; Yuan, W.; Zhu, W.W.; Zheng, Y.; et al. Dietary administration of Bacillus subtilis HAINUP40 enhances growth, digestive enzyme activities, innate immune responses and disease resistance of tilapia, Oreochromis niloticus. Fish Shellfish Immun. 2017, 60, 326–333. [Google Scholar] [CrossRef]
- Huang, W.; Cheng, Z.Q.; Lei, S.N.; Liu, L.Y.; Lv, X.; Chen, L.H.; Wu, M.H.; Wang, C.; Tian, B.Y.; Song, Y.K. Community composition, diversity, and metabolism of intestinal microbiota in cultivated European eel (Anguilla anguilla). Appl. Microbiol.Biot. 2018, 102, 4143–4157. [Google Scholar] [CrossRef]
- Zhang, J.J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAdmergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Doan, H.V.; Hoseinifar, S.H.; Dawood, M.A.O.; Chitmanat, C.; Tayyamath, K. Effects of Cordyceps militaris spent mushroom substrate and Lactobacillus plantarum on mucosal, serum immunology and growth performance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immun. 2017, 70, 87–94. [Google Scholar] [CrossRef]
- Soltani, M.; Abdy, E.; Alishahi, M.; Mirghaed, A.T.; Hosseini-Shekarabi, P. Growth performance, immune-physiological variables and disease resistance of common carp (Cyprinus carpio) orally subjected to different concentrations of Lactobacillus plantarum. Aquac. Int. 2017, 25, 1913–1933. [Google Scholar] [CrossRef]
- Vendrell, D.; Balcázar, J.L.; Blas, I.D.; Ruiz-Zarzuela, I.; Gironés, O.; Múzquiz, J.L. Protection of rainbow trout (Oncorhynchus mykiss) from lactococcosis by probiotic bacteria. Comp. Immun. Microbiol. Infect. Dis. 2008, 31, 337–345. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Basuini, M.F.E.; Hossain, M.S.; Nhu, T.H.; Dossou, S.; Moss, A.S. Effects of dietary supplementation of Lactobacillus rhamnosus or/and Lactococcus lactis on the growth, gut microbiota and immune responses of red sea bream, Pagrus major. Fish Shellfish Immun. 2016, 49, 275–285. [Google Scholar] [CrossRef]
- Kim, D.; Beck, B.R.; Heo, S.; Kim, J.J.; Kim, H.D.; Lee, S.; Kim, Y.; Oh, S.Y.; Lee, K.; Do, H.K.; et al. Lactococcus lactis BFE920 activates the innate immune system of olive flounder (Paralichthys olivaceus), resulting in protection against Streptococcus iniae infection and enhancing feed efficiency and weight gain in large-scale field studies. Fish Shellfish Immun. 2013, 35, 1585–1590. [Google Scholar] [CrossRef]
- Andani, H.R.R.; Tukmechi, A.; Meshkini, S.; Sheikhzadeh, N. Antagonistic activity of two potential probiotic bacteria from fish intestines and investigation of their effects on growth performance and immune response in rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2012, 28, 728–734. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Aly, S.M.; Ahmed, Y.A.; Ghareeb, A.A.; Mohamed, M.F. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immun. 2008, 25, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Park, K.H.; Yoon, T.J.; Kim, J.B.; Jang, Y.S.; Choe, C.H. Dietary Korean mistletoe enhances cellular non-specific immune responses and survival of Japanese eel (Anguilla japonica). Fish Shellfish Immun. 2008, 24, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.B.; Zhang, Z.; Wang, Y.B.; Li, S.N.; Ran, C.; Hu, J.; Xie, Y.D.; Li, W.F.; Zhou, Z.G. EPSP of L. casei BL23 Protected against the Infection Caused by Aeromonas veronii via Enhancement of Immune Response in Zebrafish. Front. Microbiol. 2017, 8, 2406. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.M.; Kim, H.K.; Kim, O.J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immun. 2015, 42, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Katya, K.; Park, Y.J.; Won, S.H.; Seong, M.J.; Hamidoghli, A.; Bai, S.C. Comparative evaluation of dietary probiotics Bacillus subtilis WB60 and Lactobacillus plantarum KCTC3928 on the growth performance, immunological parameters, gut morphology and disease resistance in Japanese eel, Anguilla japonica. Fish Shellfish Immun. 2017, 61, 201–210. [Google Scholar] [CrossRef]
- Butprom, S.; Phumkhachorn, P.; Rattanachaikunsopon, P. Effect of Lactobacillus plantarum C014 on Innate Immune Response and Disease Resistance against Aeromonas hydrophila in Hybrid Catfish. Sci. World J. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Larsen, A.M.; Mohammed, H.H.; Arias, C.R. Characterization of the gut microbiota of three commercially valuable warmwater fish species. J. Appl. Microbiol. 2014, 116, 1396–1404. [Google Scholar] [CrossRef]
- Bledsoe, J.W.; Peterson, B.C.; Swanson, K.S.; Small, B.C. Ontogenetic Characterization of the Intestinal Microbiota of Channel Catfish through 16S rRNA Gene Sequencing Reveals Insights on Temporal Shifts and the Influence of Environmental Microbes. PLoS ONE 2016, 11, e166379. [Google Scholar] [CrossRef]
- Tanasomwang, V.; Muroga, K. Intestinal microflora of rockfish Sebastes schlegeli, tiger puffer Takifugu rubripes and red grouper Epinephelus akaara at their larval and juvenile stages. Nippon Suisan Gakk. 1989, 55, 1371–1377. [Google Scholar] [CrossRef] [Green Version]
- Xun, P.W.; Lin, H.Z.; Wang, R.X.; Huang, Z.; Zhou, C.P.; Yu, W.; Huang, Q.Q.; Tan, L.J.; Wang, Y.; Wang, J. Effects of dietary vitamin B1 on growth performance, intestinal digestion and absorption, intestinal microflora and immune response of juvenile golden pompano (Trachinotus ovatus). Aquaculture 2019, 506, 75–83. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Bouvier, C.; Bouvier, T.; Troussellier, M.; Escalas, A.; Mouillot, D. Genetic difference but functional similarity among fish gut bacterial communities through molecular and biochemical fingerprints. FEMS Microbiol. Ecol. 2012, 79, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.G.; Wang, G.T.; Angert, E.R.; Wang, W.W.; Li, W.X.; Zou, H. Composition, diversity, and origin of the bacterial community in grass carp intestine. PLoS ONE 2012, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pond, M.J.; Stone, D.M.; Alderman, D.J. Comparison of conventional and molecular techniques to investigate the intestinal microflora of rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 194–203. [Google Scholar] [CrossRef]
- Wu, S.; Gao, T.H.; Zheng, Y.Z.; Wang, W.W.; Cheng, Y.Y.; Wang, G.T. Microbial diversity of intestinal contents and mucus in yellow catfish (Pelteobagrus fulvidraco). Aquaculture 2010, 303, 1–7. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Proportion |
|---|---|
| Fish meal | 8.00 |
| Soybean meal | 45.00 |
| Corn meal | 25.00 |
| Wheat middling | 7.40 |
| Corn oil | 3.60 |
| Dicalcium phosphate | 1.00 |
| Vitamin mix1 | 0.50 |
| Mineral mix2 | 0.50 |
| Cellulose | 6.00 |
| Carboxymethyl cellulose | 3.00 |
| Proximate composition | |
| Moisture | 7.84 |
| Protein | 30.56 |
| Lipid | 5.76 |
| Ash | 6.84 |
| CG | Lc | Lp | |
|---|---|---|---|
| Initial weight (g) | 67.43 ± 1.31 | 67.18 ± 1.42 | 67.01 ± 1.64 |
| Final weight (g) | 108.40 ± 9.57 b | 114.67 ± 10.75 a | 110.57 ± 10.61 b |
| WG (g) | 40.95 ± 0.94 b | 47.49 ± 1.62 a | 43.56 ± 1.77 b |
| Survival rate (%) | 98.81 ± 2.06 a | 98.81 ± 2.06 a | 100.00 ± 0.00 a |
| SGR (%) | 1.69 ± 0.03 b | 1.90 ± 0.05 a | 1.79 ± 0.06 b |
| FCR (%) | 0.98 ± 0.07 a | 0.82 ± 0.02 b | 0.89 ± 0.04 ab |
| Sample | Shannon Index | ACE | Chao1 Index | Goods Coverage | Simpson |
|---|---|---|---|---|---|
| CG | 2.16 ± 0.29 ab | 1041.25 ± 199.74 a | 600.46 ± 53.19 ab | 0.997 ± 0.000 c | 0.21 ± 0.06 b |
| Lc | 0.98 ± 1.10 bc | 472.49 ± 316.03 b | 455.37 ± 324.30 b | 0.999 ± 0.001 a | 0.75 ± 0.28 a |
| Lp | 0.97 ± 0.68 bc | 531.92 ± 148.74 b | 466.16 ± 192.22 b | 0.998 ± 0.001 b | 0.73 ± 0.19 a |
| CG-2wk | 3.34 ± 0.88 a | 1048.15 ± 358.86 a | 1008.51 ± 489.45 a | 0.997 ± 0.000 bc | 0.16 ± 0.03 b |
| Lc-2wk | 3.28 ± 0.16 a | 513.82 ± 30.60 b | 518.77 ± 22.11 b | 0.999 ± 0.000 a | 0.10 ± 0.01 b |
| Lp-2wk | 0.69 ± 0.44 c | 808.84 ± 60.62 ab | 640.22 ± 126.39 ab | 0.997 ± 0.000 bc | 0.74 ± 0.25 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, H.; Hu, K.; Jiao, L.; Zhao, M.; Yang, X.; Xia, L. Effect of Dietary Supplementation of Lactobacillus Casei YYL3 and L. Plantarum YYL5 on Growth, Immune Response and Intestinal Microbiota in Channel Catfish. Animals 2019, 9, 1005. https://doi.org/10.3390/ani9121005
Zhang H, Wang H, Hu K, Jiao L, Zhao M, Yang X, Xia L. Effect of Dietary Supplementation of Lactobacillus Casei YYL3 and L. Plantarum YYL5 on Growth, Immune Response and Intestinal Microbiota in Channel Catfish. Animals. 2019; 9(12):1005. https://doi.org/10.3390/ani9121005
Chicago/Turabian StyleZhang, Hongyu, Haibo Wang, Kun Hu, Liting Jiao, Mingjun Zhao, Xianle Yang, and Lei Xia. 2019. "Effect of Dietary Supplementation of Lactobacillus Casei YYL3 and L. Plantarum YYL5 on Growth, Immune Response and Intestinal Microbiota in Channel Catfish" Animals 9, no. 12: 1005. https://doi.org/10.3390/ani9121005
APA StyleZhang, H., Wang, H., Hu, K., Jiao, L., Zhao, M., Yang, X., & Xia, L. (2019). Effect of Dietary Supplementation of Lactobacillus Casei YYL3 and L. Plantarum YYL5 on Growth, Immune Response and Intestinal Microbiota in Channel Catfish. Animals, 9(12), 1005. https://doi.org/10.3390/ani9121005