Forgotten, But Not Lost—Alloparental Behavior and Pup–Adult Interactions in Companion Dogs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Development of the Survey
2.3. Subjects
2.4. Variables
- -
- Alloparental behaviors (apart from feeding by regurgitation); e.g., when another female dog nursed, or attempted to nurse the puppies. (Multiple choice, where the possible answers were: No alloparental behaviors were observed; puppies were nursed by—the daughter, the mother, the sister of the mother dog; an unrelated adult female nursed the puppies.)
- -
- Feeding with regurgitation (we asked the participants to indicate separately whether the mother of the puppies and/or another dog regurgitated food for the puppies). Multiple choice item.
- -
- Puppies’ reaction to adult dogs’ barking: Breeders could select one of three options: Puppies show no reaction, puppies become frightened (run and hide, become quiet), or puppies start to bark when they hear the adults’ barking.
- -
- Adult dogs’ reaction to the puppies’ whining: We again used a three grade scale (single choice): Adults show no reaction to puppies’ whining, they sniff the puppies, or the adults show alloparental behavior (mother-like caretaking behavior towards the puppies: Licks them, lays down next to them, plays with them).
- -
- Adult dogs’ reaction to puppies in general situations: During play, eating, drinking, and chewing on toys (single choice). We got the majority of answers from the following two categories: Adults show no reaction to the puppies, or adults are friendly with the puppies. Other categories, such as adults start barking, show aggression towards the puppies, puppies show aggression towards adults, were excluded due to the very sporadic or missing answers to these options.
- -
- Mother dogs’ reaction to adult dogs around her puppies (single choice). Breeders could again indicate their answers on a three-grade scale: She shows no reaction, she is aggressive with other dogs, or she is friendly with other dogs.
- -
- The way the puppies are separated from the other dogs. Three alternatives were given: Puppies and adult dogs are kept together, puppies are separated with a fence or barrier, or puppies are separated completely from adult dogs. (i.e., they live in a separate room of the house).
- -
- Age of the puppies when they are separated from adult dogs. Participants could choose from three main options: The puppies and adult dogs are kept together, the puppies are kept separated only until a defined age, or the puppies are separated completely from other adult dogs the entire time they are at the breeder.
- -
- Aggression towards puppies. The breeders were asked whether they experienced any aggression towards the puppies in general situations (yes/no).
- -
- The age of the other dog: Younger than one year, between one and eight years, and over eight years old.
- -
- Paternal status of the other dogs (whether it was the father or not of the current puppies).
- -
- Previous parental experience of adult dogs (i.e., has the dog already reproduced, in the case of both male and female dogs); yes or no answers could be selected.
- -
- Sexual status of the adult dogs: Breeders could select whether the particular dog was neutered/spayed or intact.
2.5. Data Analysis
3. Results
3.1. Alloparental Behaviors (Including Nursing, Licking, Cleaning, But Not Regurgitation)
3.2. Feeding of the Puppies with Regurgitation
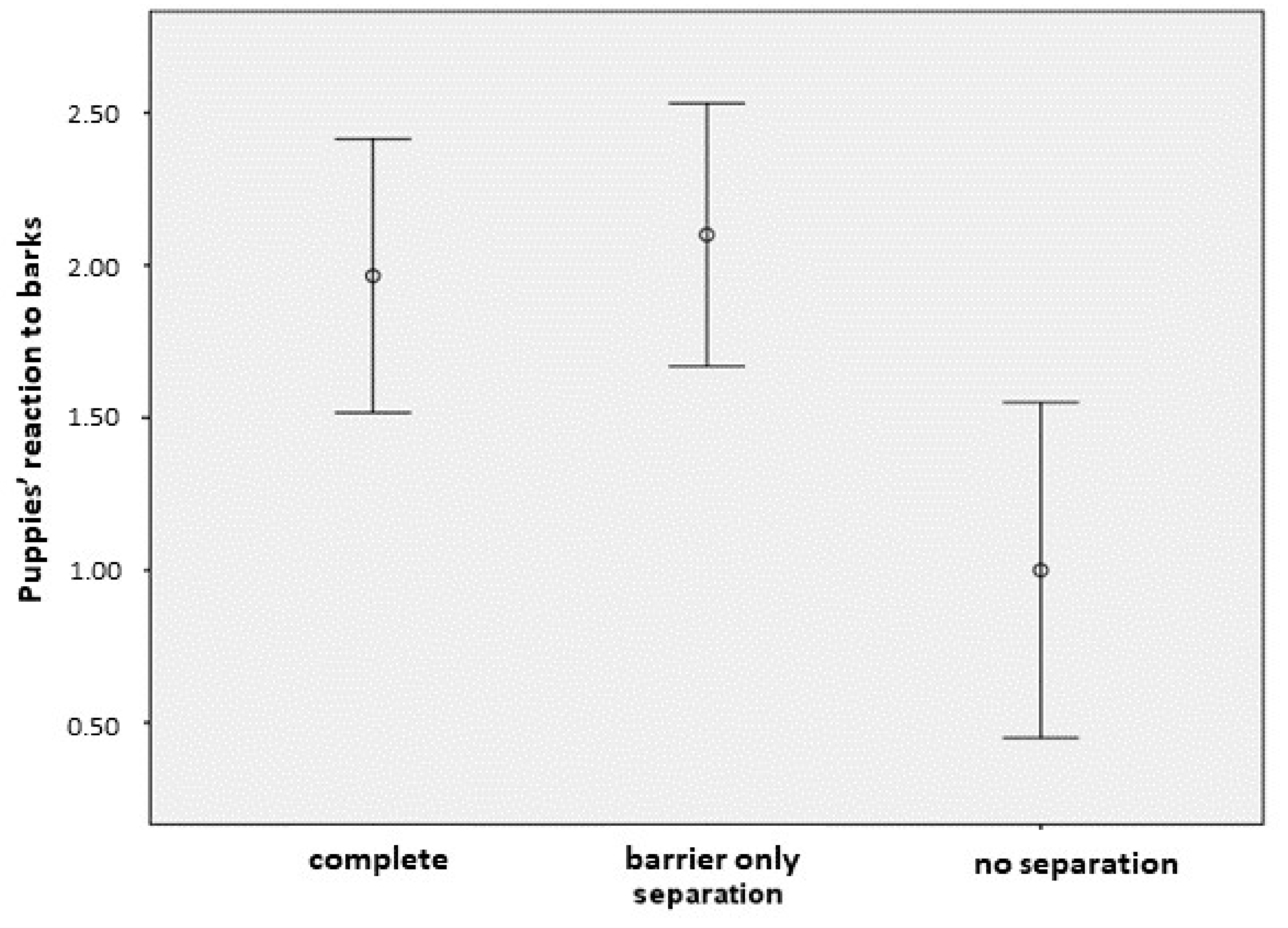
3.3. The Puppies’ Reaction to the Other Dogs’ Barking
3.4. Adult Dogs’ Reaction to the Puppies’ Whining
3.5. Adult Dogs’ Reaction to Puppies in General Situations
3.6. The Reaction of the Mother of the Puppies to Other Dogs in the Household
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Topál, J.; Miklósi, Á.; Gácsi, M.; Dóka, A.; Pongrácz, P.; Kubinyi, E.; Csányi, V. The dog as a model for understanding human social behavior. Adv. Study Behav. 2009, 39, 71–116. [Google Scholar]
- Udell, M.A.; Dorey, N.R.; Wynne, C.D. What did domestication do to dogs? A new account of dogs’ sensitivity to human actions. Biol. Rev. 2010, 85, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Topál, J.; Gácsi, M.; Miklósi, Á.; Virányi, Z.; Kubinyi, E.; Csányi, V. Attachment to humans: A comparative study on hand-reared wolves and differently socialized dog puppies. Anim. Behav. 2005, 70, 1367–1375. [Google Scholar] [CrossRef]
- Marshall-Pescini, S.; Virányi, Z.; Range, F. The effect of domestication on inhibitory control: Wolves and dogs compared. PLoS ONE 2015, 10, e0118469. [Google Scholar] [CrossRef]
- Clutton-Brock, J. Origins of the dog: Domestication and early history. In The Domestic Dog: Its Evolution, Behaviour and Interactions with People; Serpell, J., Barrett, P., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 7–20. [Google Scholar]
- Gácsi, M.; McGreevy, P.; Kara, E.; Miklósi, Á. Effects of selection for cooperation and attention in dogs. Behav. Brain Funct. 2009, 5, 31. [Google Scholar] [CrossRef]
- Belyaev, D.K.; Plyusnina, I.Z.; Trut, L.N. Domestication in the silver fox (Vulpes fulvus Desm): Changes in physiological boundaries of the sensitive period of primary socialization. Appl. Anim. Behav. Sci. 1985, 13, 359–370. [Google Scholar] [CrossRef]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.-L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, Å.; Lindblad-Toh, K. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 2013, 495, 360. [Google Scholar] [CrossRef]
- Miklósi, A.; Topál, J.; Csányi, V. Comparative social cognition: What can dogs teach us? Anim. Behav. 2004, 67, 995–1004. [Google Scholar] [CrossRef]
- Range, F.; Virányi, Z. Tracking the evolutionary origins of dog-human cooperation: The Canine Cooperation Hypothesis. Front. Psychol. 2015, 5, 1582. [Google Scholar] [CrossRef]
- Arendt, M.; Fall, T.; Lindblad-Toh, K.; Axelsson, E. Amylase activity is associated with AMY 2B copy numbers in dog: Implications for dog domestication, diet and diabetes. Anim. Genet. 2014, 45, 716–722. [Google Scholar] [CrossRef]
- Arendt, M.; Cairns, K.M.; Ballard, J.W.O.; Savolainen, P.; Axelsson, E. Diet adaptation in dog reflects spread of prehistoric agriculture. Heredity 2016, 117, 301. [Google Scholar] [CrossRef] [PubMed]
- Belyaev, D.K. Destabilizing selection as a factor in domestication. J. Hered. 1979, 70, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Harrop, A.E. Reproduction in the Dog; Williams and Wilkins: Philadelphia, PA, USA, 1960. [Google Scholar]
- Hoffman, J.M.; Creevy, K.E.; Promislow, D.E. Reproductive capability is associated with lifespan and cause of death in companion dogs. PLoS ONE 2013, 8, e61082. [Google Scholar] [CrossRef] [PubMed]
- McGreevy, P.D.; Nicholas, F.W. Some practical solutions to welfare problems in dog breeding. Anim. Welf. 1999, 8, 329–342. [Google Scholar]
- Kustritz, M.V.R. Determining the optimal age for gonadectomy of dogs and cats. J. Am. Vet. Med. Assoc. 2007, 231, 1665–1675. [Google Scholar] [CrossRef]
- Asa, C.S.; Valdespino, C. Canid reproductive biology: An integration of proximate mechanisms and ultimate causes. Am. Zool. 1998, 38, 251–259. [Google Scholar] [CrossRef]
- Paul, M.; Majumder, S.S.; Bhadra, A. Grandmotherly care: A case study in Indian free-ranging dogs. J. Ethol. 2014, 32, 75–82. [Google Scholar] [CrossRef]
- Paul, M.; Bhadra, A. The great Indian joint families of free-ranging dogs. PLoS ONE 2018, 13, e0197328. [Google Scholar] [CrossRef]
- Coppinger, R.; Coppinger, L. What is a Dog? University of Chicago Press: Chicago, IL, USA, 2016. [Google Scholar]
- Matter, H.C.; Daniels, T.J. Dogs, Zoonoses and Public Health. In Dog Ecology and Population Biology; McPherson, C.N.L., Meslin, F.X., Wandeler, A.I., Eds.; CABI: Wallingford, UK, 2000; pp. 17–62. [Google Scholar]
- Coppinger, R.; Coppinger, L. Dogs: A New Understanding of Canine Origin, Behavior and Evolution; University of Chicago Press: Chicago, IL, USA, 2002. [Google Scholar]
- Pal, S.K. Reproductive behaviour of free-ranging rural dogs in West Bengal, India. Acta Theriol. 2003, 48, 271–281. [Google Scholar] [CrossRef]
- Lord, K.; Feinstein, M.; Smith, B.; Coppinger, R. Variation in reproductive traits of members of the genus Canis with special attention to the domestic dog (Canis familiaris). Behav. Proc. 2013, 92, 131–142. [Google Scholar] [CrossRef]
- Sacks, B.N.; Brown, S.K.; Stephens, D.; Pedersen, N.C.; Wu, J.T.; Berry, O. Y chromosome analysis of dingoes and Southeast Asian village dogs suggests a Neolithic continental expansion from Southeast Asia followed by multiple Austronesian dispersals. Mol. Biol. Evol. 2013, 30, 1103–1118. [Google Scholar] [CrossRef] [PubMed]
- Thomson, P.C. The behavioural ecology of dingoes in north-western Australia. II. Activity patterns, breeding season and pup rearing. Wildl. Res. 1992, 19, 519–529. [Google Scholar] [CrossRef]
- Seksel, K. Puppy socialization classes. Vet. Clin. N. Am. Small 1997, 27, 465–477. [Google Scholar] [CrossRef]
- Gazzano, A.; Mariti, C.; Alvares, S.; Cozzi, A.; Tognetti, R.; Sighieri, C. The prevention of undesirable behaviors in dogs: Effectiveness of veterinary behaviorists’ advice given to puppy owners. J. Vet. Behav. 2008, 3, 125–133. [Google Scholar] [CrossRef]
- Howell, T.J.; King, T.; Bennett, P.C. Puppy parties and beyond: The role of early age socialization practices on adult dog behavior. Vet. Med. Res. Rep. 2015, 6, 143. [Google Scholar] [CrossRef]
- Řezáč, P.; Viziová, P.; Dobešová, M.; Havlíček, Z.; Pospíšilová, D. Factors affecting dog–dog interactions on walks with their owners. Appl. Anim. Behav. Sci. 2011, 134, 170–176. [Google Scholar] [CrossRef]
- Ward, C.; Bauer, E.B.; Smuts, B.B. Partner preferences and asymmetries in social play among domestic dog, Canis lupus familiaris, littermates. Anim. Behav. 2008, 76, 1187–1199. [Google Scholar] [CrossRef]
- Slabbert, J.M.; Rasa, O.A.E. Observational learning of an acquired maternal behaviour pattern by working dog pups: An alternative training method? Appl. Anim. Behav. Sci. 1997, 53, 309–316. [Google Scholar] [CrossRef]
- Foyer, P.; Wilsson, E.; Jensen, P. Levels of maternal care in dogs affect adult offspring temperament. Sci. Rep. 2016, 6, 19253. [Google Scholar] [CrossRef]
- Malm, K. Regurgitation in relation to weaning in the domestic dog: A questionnaire study. Appl. Anim. Behav. Sci. 1995, 43, 111–122. [Google Scholar] [CrossRef]
- Pal, S.K. Parental care in free-ranging dogs, Canis familiaris. Appl. Anim. Behav. Sci. 2005, 90, 31–47. [Google Scholar] [CrossRef]
- Scott, J.P.; Fredericson, E.; Fuller, J.L. Experimental exploration of the critical period hypothesis. Personality 1951, 1, 162–183. [Google Scholar]
- Gácsi, M.; Győri, B.; Miklósi, Á.; Virányi, Z.; Kubinyi, E.; Topál, J.; Csányi, V. Species-specific differences and similarities in the behavior of hand-raised dog and wolf pups in social situations with humans. Dev. Psychobiol. 2005, 47, 111–122. [Google Scholar] [CrossRef]
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic analyses reveal the influence of geographic origin, migration, and hybridization on modern dog breed development. Cell Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Wilsson, E.; Sundgren, P.E. The use of a behaviour test for the selection of dogs for service and breeding, I: Method of testing and evaluating test results in the adult dog, demands on different kinds of service dogs, sex and breed differences. Appl. Anim. Behav. Sci. 1997, 53, 279–295. [Google Scholar] [CrossRef]
- King, T.; Marston, L.C.; Bennett, P.C. Describing the ideal Australian companion dog. Appl. Anim. Behav. Sci. 2009, 120, 84–93. [Google Scholar] [CrossRef]
- Kustritz, M.V.R. Reproductive behavior of small animals. Theriogenology 2005, 64, 734–746. [Google Scholar] [CrossRef]
- Lezama-García, K.; Mariti, C.; Mota-Rojas, D.; Martínez-Burnes, J.; Barrios-García, H.; Gazzano, A. Maternal behaviour in domestic dogs. Int. J. Vet. Sci. Med. 2019, 7, 20–30. [Google Scholar] [CrossRef]
- Riedman, M.L. The evolution of alloparental care and adoption in mammals and birds. Q. Rev. Biol. 1982, 57, 405–435. [Google Scholar] [CrossRef]
- Boitani, L.; Ciucci, P.; Ortolani, A. Behaviour and social ecology of free-ranging dogs. In The Behavioural Biology of Dogs; Jensen, P., Ed.; CAB International: Wallingford, UK, 2007; pp. 147–165. [Google Scholar]
- Pal, S.K.; Ghosh, B.; Roy, S. Dispersal behaviour of free-ranging dogs (Canis familiaris) in relation to age, sex, season and dispersal distance. Appl. Anim. Behav. Sci. 1998, 61, 123–132. [Google Scholar] [CrossRef]
- Scott, J.P. The process of primary socialization in canine and human infants. Monogr. Soc. Res. Child Dev. 1963, 28, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Duxbury, M.M.; Jackson, J.A.; Line, S.W.; Anderson, R.K. Evaluation of association between retention in the home and attendance at puppy socialization classes. J. Am. Vet. Med. Assoc. 2003, 223, 61–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dendoncker, P.A.; De Keuster, T.; Diederich, C.; Dewulf, J.; Moons, C.P. On the origin of puppies: Breeding and selling procedures relevant for canine behavioural development. Vet. Record. 2019, 184, 710. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.E. The Effects of Differential Early Treatment on the Social and Exploratory Behavior of Puppies. Ph.D. Thesis, Pennsylvania State University, University Park, PA, USA, 1955. [Google Scholar]
- Howell, T.J.; Bennett, P.C. Puppy power! Using social cognition research tasks to improve socialization practices for domestic dogs (Canis familiaris). J. Vet. Behav. 2011, 6, 195–204. [Google Scholar] [CrossRef]
- Wormald, D.; Lawrence, A.J.; Carter, G.; Fisher, A.D. Analysis of correlations between early social exposure and reported aggression in the dog. J. Vet. Behav. 2016, 15, 31–36. [Google Scholar] [CrossRef]
- Lenkei, R.; Pogány, Á.; Fugazza, C. Social behavior in dog puppies: Breed differences and the effect of rearing conditions. Biol. Fut. 2019, 70, 134–142. [Google Scholar] [CrossRef]
- Lehoczki, F.; Szamosvölgyi, Z.; Miklósi, Á.; Faragó, T. Dogs’ sensitivity to strange pup separation calls: Pitch instability increases attention regardless of sex and experience. Anim. Behav. 2019, 153, 115–129. [Google Scholar] [CrossRef]
- Smith, M.S.; Mc Donald, L.E. Serum levels of luteinizing hormone and progesterone during the estrous cycle, pseudopregnancy and pregnancy in the dog. Endocrinology 1974, 94, 404–412. [Google Scholar] [CrossRef]
- Schradin, C.; Vuarin, P.; Rimbach, R. The neoteny-helper hypothesis: When to expect and when not to expect endocrine mechanisms to regulate allo-parental care? Physiol. Behav. 2018, 193, 127–134. [Google Scholar] [CrossRef]
- Pal, S.K. Urine marking by free-ranging dogs (Canis familiaris) in relation to sex, season, place and posture. Appl. Anim. Behav. Sci. 2003, 80, 45–59. [Google Scholar] [CrossRef]
- Pal, S.K.; Ghosh, B.; Roy, S. Agonistic behaviour of free-ranging dogs (Canis familiaris) in relation to season, sex and age. Appl. Anim. Behav. Sci. 1998, 59, 331–348. [Google Scholar] [CrossRef]
- McLeod, P.J. Infanticide by female wolves. Can. J. Zool. 1990, 68, 402–404. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | FCI Breed Group | Total | Questionnaire 2 only | Questionnaires 1 and 2 | Country |
|---|---|---|---|---|---|
| Australian Shepherd | FCI-1 | 4 | 2 | Hungary, Germany | |
| Belgian Shepherd Dog/Malinois | FCI-1 | 2 | 1 | 1 | Hungary, UK |
| Border Collie | FCI-1 | 1 | 1 | Hungary | |
| Bouvier des Flandres | FCI-1 | 1 | 1 | Hungary | |
| Collie Rough | FCI-1 | 1 | 1 | Hungary | |
| Collie Smooth | FCI-1 | 1 | Germany | ||
| German Shepherd Dog | FCI-1 | 4 | Hungary | ||
| Mudi | FCI-1 | 5 | 1 | 1 | Hungary |
| Old English Sheepdog | FCI-1 | 1 | Poland | ||
| Pumi | FCI-1 | 1 | Sweden | ||
| Shetland Sheepdog | FCI-1 | 2 | Hungary | ||
| Cane Corso | FCI-2 | 1 | Hungary | ||
| Doberman | FCI-2 | 4 | 2 | 2 | Hungary, Germany |
| Great Dane | FCI-2 | 2 | Hungary | ||
| Kangal | FCI-2 | 1 | 1 | Hungary | |
| Rottweiler | FCI-2 | 3 | 1 | 1 | Hungary, US |
| Schnauzer—Giant | FCI-2 | 3 | Hungary | ||
| Schnauzer—Middle | FCI-2 | 1 | 1 | Hungary | |
| Shar Pei | FCI-2 | 1 | Hungary | ||
| Am. Pitbull Terrier | FCI-3 | 3 | 1 | Hungary | |
| Am. Staffordshire Terrier | FCI-3 | 2 | Hungary | ||
| Biewer Yorkshire Terrier | FCI-3 | 1 | Hungary | ||
| Jack Russell Terrier | FCI-3 | 1 | Hungary | ||
| Yorkshire Terrier | FCI-3 | 1 | Hungary | ||
| Dachshund Mini | FCI-4 | 1 | Finland | ||
| Dachshund Short Haired | FCI-4 | 1 | Belgium | ||
| Akita Inu | FCI-5 | 1 | 1 | Hungary | |
| German Spitz Klein | FCI-5 | 2 | Hungary, UK | ||
| Hokkaido | FCI-5 | 1 | Hungary | ||
| Keeshond (Wolfspitz) | FCI-5 | 1 | Germany | ||
| Shiba Inu | FCI-5 | 1 | Lithuania | ||
| Siberian Husky | FCI-5 | 3 | 1 | Hungary | |
| Rhodesian Ridgeback | FCI-6 | 1 | 1 | Germany | |
| Braque d’Auvergne | FCI-7 | 1 | Hungary | ||
| Hungarian Vizsla | FCI-7 | 2 | 1 | Hungary | |
| English Cocker Spaniel | FCI-8 | 1 | 1 | Hungary | |
| Flatcoated Retriever | FCI-8 | 1 | 1 | Finland | |
| Labrador Retriever | FCI-8 | 5 | 1 | Hungary, UK, Italy | |
| Chinese Crested Dog | FCI-9 | 3 | 1 | Hungary | |
| Bichon Frise | FCI-9 | 1 | 1 | Hungary | |
| Bolognese | FCI-9 | 2 | 1 | Hungary, Slovenia | |
| Coton de Tulear | FCI-9 | 1 | 1 | Hungary | |
| French Bulldog | FCI-9 | 2 | 1 | Hungary | |
| Havanese | FCI-9 | 4 | 1 | Hungary | |
| Lhasa Apso | FCI-9 | 1 | Hungary | ||
| Poodle—all sizes | FCI-9 | 3 | 1 | Hungary | |
| Shih Tzu | FCI-9 | 1 | Hungary | ||
| Tibetan Terrier | FCI-9 | 3 | 1 | 1 | Hungary |
| Borzoi | FCI-10 | 1 | 1 | Hungary | |
| Whippet | FCI-10 | 3 | 1 | Hungary, Austria |
| Parameter | Estimate | Std. Error | Wald | df | Sig. | 95% Confidence Interval | ||
|---|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | |||||||
| Threshold | [barkreact = 0] | −0.317 | 0.472 | 0.450 | 1 | 0.502 | −1.243 | 0.609 |
| [barkreact = 1] | 0.893 | 0.486 | 3.367 | 1 | 0.067 | −0.061 | 1.846 | |
| [barkreact = 2] | 1.496 | 0.504 | 8.809 | 1 | 0.003 | 0.508 | 2.484 | |
| Location | [separation = 1] | 1.461 | 0.589 | 6.154 | 1 | 0.013 | 0.307 | 2.615 |
| [separation = 2] | 1.651 | 0.592 | 7.771 | 1 | 0.005 | 0.490 | 2.812 | |
| [separation = 3] | 0 * | . | . | 0 | . | . | . | |
| Parameter | B | Std. Error | 95% Wald Confidence Interval | Hypothesis Test | ||||
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Wald Chi-Square | df | Sig. | ||||
| Threshold | [react_whine = 1] | −1.485 | 0.4545 | −2.375 | −0.594 | 10.671 | 1 | 0.001 |
| [react_whine = 2] | 0.912 | 0.4410 | 0.047 | 1.776 | 4.275 | 1 | 0.039 | |
| [age = below 1 year] | 2.113 | 7824 | 0.579 | 3.646 | 7.292 | 1 | 0.007 | |
| [age = between 1–8 years] | 0.579 | 0.5375 | −0.474 | 1.633 | 1.162 | 1 | 0.281 | |
| [age = older than 8 years] | 0 * | . | . | . | . | . | . | |
| [spayed] | −1.152 | 0.5439 | −2.218 | −0.086 | 4.488 | 1 | 0.034 | |
| [intact] | 0 | . | . | . | . | . | . | |
| [parous = 0] | −1.140 | 0.4780 | −2.077 | −0.203 | 5.690 | 1 | 0.017 | |
| [parous = 1] | 0 * | . | . | . | . | . | . | |
| Parameter | B | Std. Error | 95% Wald Confidence Interval | Hypothesis Test | ||||
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Wald Chi-Square | df | Sig. | ||||
| Threshold | [react_whine = 1] | −2.322 | 0.3886 | −3.083 | −1.560 | 35.694 | 1 | 0.000 |
| [react_whine = 2] | 0.396 | 0.3295 | −0.250 | 1.041 | 1.441 | 1 | 0.230 | |
| [male] | −0.963 | 0.4643 | −1.873 | −0.053 | 4.302 | 1 | 0.038 | |
| [female] | 0 * | . | . | . | . | . | . | |
| [parous = 0] | −1.273 | 0.4984 | −2.250 | −0.296 | 6.527 | 1 | 0.011 | |
| [parous = 1] | 0 * | . | . | . | . | . | . | |
| Parameter | B | Std. Error | 95% Wald Confidence Interval | Hypothesis Test | |||
|---|---|---|---|---|---|---|---|
| Lower | Upper | Wald Chi-Square | df | Sig. | |||
| (Intercept) | 0.241 | 0.4029 | −0.549 | 1.031 | 0.358 | 1 | 0.549 |
| [age = below 1 year] | −2.044 | 0.7752 | −3.564 | −0.525 | 6.955 | 1 | 0.008 |
| [age = 1–8 yeas] | −1.259 | 0.5064 | −2.252 | −0.266 | 6.181 | 1 | 0.013 |
| [age = older than 8 years] | 0 * | . | . | . | . | . | . |
| [spayed] | 0.935 | 0.3727 | 0.204 | 1.666 | 6.293 | 1 | 0.012 |
| [intact] | 0 | . | . | . | . | . | . |
| [parous = 0] | 0.194 | 0.3723 | −0.536 | 0.923 | 0.271 | 1 | 0.603 |
| [parous = 1] | 0 * | . | . | . | . | . | . |
| Parameter | B | Std. Error | 95% Wald Confidence Interval | Hypothesis Test | ||||
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Wald Chi-Square | df | Sig. | ||||
| Threshold | [react_female = −1] | −2.019 | 0.3824 | −2.768 | −1.269 | 27.876 | 1 | 0.000 |
| [react_female = 0] | −0.751 | 0.3556 | −1.448 | −0.054 | 4.463 | 1 | 0.035 | |
| [age = less than 1 year] | 1.744 | 0.6716 | 0.428 | 3.061 | 6.743 | 1 | 0.009 | |
| [age = 1–8 years] | −0.195 | 0.3999 | −0.979 | 0.588 | 0.239 | 1 | 0.625 | |
| [age = older than 8 years] | 0 * | . | . | . | . | . | . | |
| [spayed] | −0.646 | 0.4022 | −1.434 | 0.142 | 2.579 | 1 | 0.108 | |
| [intact] | 0 * | . | . | . | . | . | . | |
| [parous = 0] | −2.003 | 0.4294 | −2.844 | −1.161 | 21.748 | 1 | 0.000 | |
| [parous = 1] | 0 * | . | . | . | . | . | . | |
| [reproductive = 0] * [parous = 1] | 2.117 | 0.6065 | 0.928 | 3.306 | 12.183 | 1 | 0.000 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pongrácz, P.; Sztruhala, S.S. Forgotten, But Not Lost—Alloparental Behavior and Pup–Adult Interactions in Companion Dogs. Animals 2019, 9, 1011. https://doi.org/10.3390/ani9121011
Pongrácz P, Sztruhala SS. Forgotten, But Not Lost—Alloparental Behavior and Pup–Adult Interactions in Companion Dogs. Animals. 2019; 9(12):1011. https://doi.org/10.3390/ani9121011
Chicago/Turabian StylePongrácz, Péter, and Sára S. Sztruhala. 2019. "Forgotten, But Not Lost—Alloparental Behavior and Pup–Adult Interactions in Companion Dogs" Animals 9, no. 12: 1011. https://doi.org/10.3390/ani9121011
APA StylePongrácz, P., & Sztruhala, S. S. (2019). Forgotten, But Not Lost—Alloparental Behavior and Pup–Adult Interactions in Companion Dogs. Animals, 9(12), 1011. https://doi.org/10.3390/ani9121011