1. Introduction
As the global climate changes, the price of commodity feed ingredients, such as corn and soybean, rise continuously. Therefore, it is imperative to find new agricultural by-products as substitutes. Every year, millions of tons of wheat are produced all over the world, and wheat bran (WB) is the main by-product [
1]. WB is inexpensive and may become one of the replacement sources for animal feed ingredients. WB consists of about 15% crude protein, 4% crude fat, 28% carbohydrate, 42% insoluble fiber, and is low in energy content. WB contains many compounds, such as ferulic acid, tocopherol, and lutein [
2], which are good antioxidants. However, it contains high fiber content, 1% phytate, about 84% phytate-p and the total-p ratio [
3] and susceptibility to mycotoxin contaminations because of improper storage. These have negative impacts on the utilization of WB as a viable ingredient in animal feed.
It is well-known that yeast cell walls are rich in ß-glucan and mannan, which can regulate immunity and protect the host from mycotoxins damage [
4]. Although past studies have indicated that the addition of
S. cerevisiae powder does not improve the growth traits of animals, it can increase the villi height and reduce the number of
E. coli in the gut [
5].
S. cerevisiae also has considerable enzyme secretion capabilities, such as proteases, xylanases, and cellulases [
6]. In addition to
S. cerevisiae, a previous study also pointed out that
A. oryzae, generally recognized as safe (GRAS) probiotic, can be used as a probiotic for fermenting; it also has good enzyme secretion ability and can increase the crude protein content of the fermented material [
7]. Moreover, many studies indicated that WB could increase its utilization value after fermentation by probiotics such as
Bacillus amyloliquefaciens,
S. cerevisiae, or white rot fungi in broilers diet [
5,
8]. Furthermore, a previous study pointed out that probiotics can decrease the inflammation response in animals [
9]. Inflammation can cause many chronic diseases that affect animal health and longevity [
10]. The reduction of the inflammatory response can decrease energy loss and improve the cell survival rate [
11].
It is well-known that phytate possess high content of phosphorus and can chelate sodium, calcium or amino acids, thus affecting the absorption and utilization of nutrients by animals; however, phytase addition could solve the above-mentioned problem, and increase the digestibility of amino acids such as Lys, Met, Cys, and Thr [
12], and the weight of the tibia, as well as increase the retention of phosphorus [
13].
Few studies have been carried out on co-fermenting WB with probiotics combined with phytase to examine inflammation in broilers. Therefore, this study aims to investigate the S. cerevisiae fermented products (SCFP) and A. oryzae fermented products (AOFP) additions in broiler diets on the growth characteristics, intestinal morphology, microbial morphology, and inflammation-related mRNA expression in broilers.
2. Materials and Methods
2.1. Probiotic Characteristics
S. cerevisiae was cultured in Yeast-Mald (YM) broth at 30 °C for 0, 6, 12, 18, and 24 h, and then the number of S. cerevisiae colony-forming unit (CFU) on YM agar (Sparks, MD 21152 USA) was counted after sequence dilution to measure the growth curve of S. cerevisiae. A. oryzae was cultured in potato dextrose broth (PDB, Neogen, Lansing, MI, USA) at 30 °C for 0, 6, 12, 18, and 24 h, and the content of dry mycelia weight of A. oryzae in PDB was measured for the growth curve of A. oryzae. Moreover, 108 CFU/mL S. cerevisiae or 106 spores/mL A. oryzae fluid were incubated in 85 °C hot water for 3 min for heat tolerance measurement. After heating, the surviving S. cerevisiae and A. oryzae were counted on the YM agar or potato dextrose agar (PDA, Neogen, Lansing, MI, USA) respectively.
Acid, base, and gastrointestinal fluid tolerance were measured by pH-adjusted phosphate buffered saline (PBS, pH 3.0, 4.0, or 13.0, adjusted by citric acid or NaOH) (Merck, Darmstadt, Germany), simulation intestinal fluid (pH 13.0, 0.3% bile acid (Merck, Darmstadt, Germany), and 0.1% trypsin (Merck, Darmstadt, Germany)), or gastric fluid (pH 3.0, 0.3% pepsin). Here, 108 CFU/mL S. cerevisiae or 106 spores/mL A. oryzae fluid were incubated in the pH adjusted fluid, simulation intestinal fluid, or gastric fluid for 0 or 3 h at 30 °C. After incubating, the surviving S. cerevisiae and A. oryzae were counted on the YM agar or PDA, respectively.
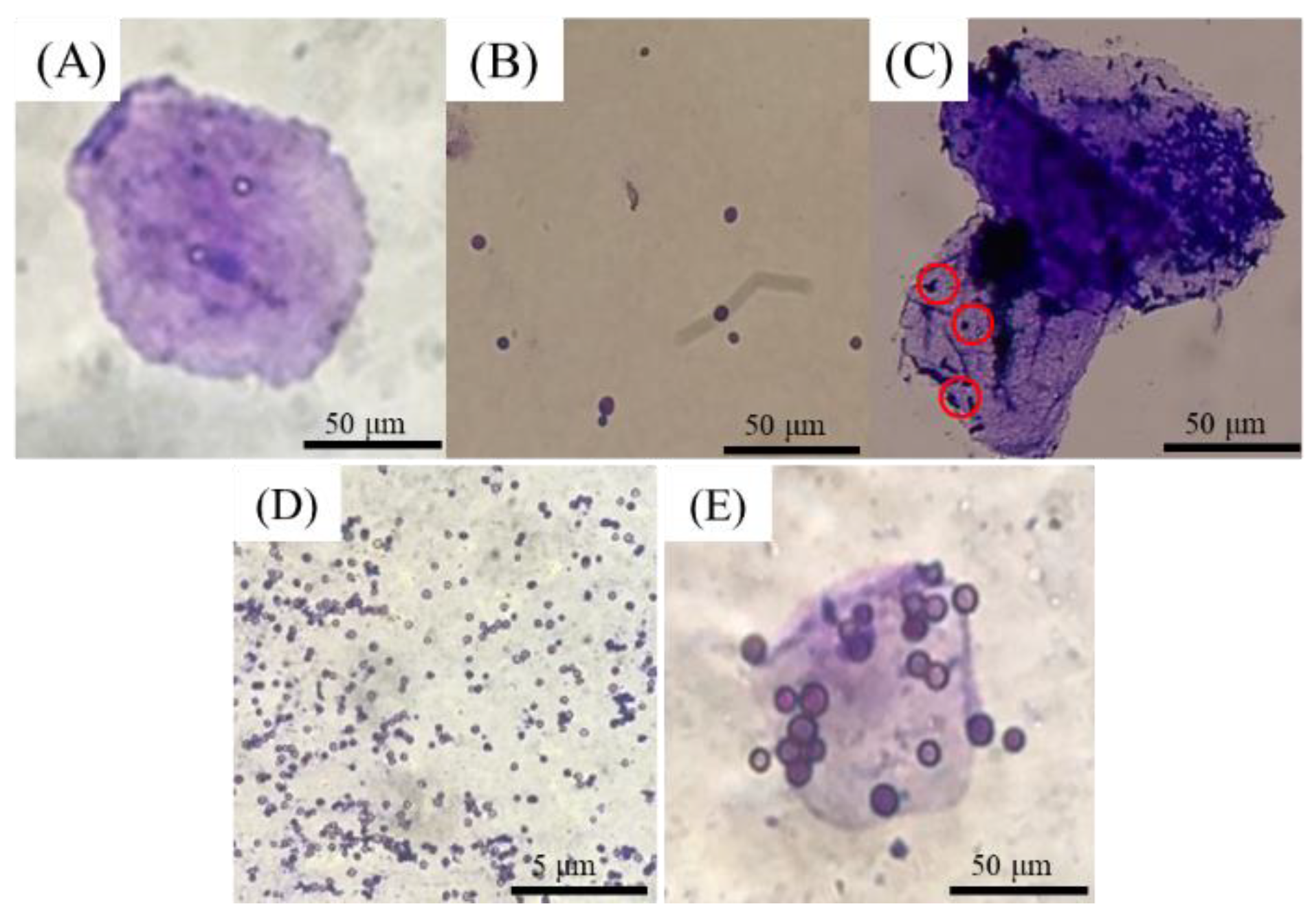
The adherence ability was measured by the method indicated by Annika et al. [
14], briefly, sacrificing the broiler and removing its crop. After dissection of the crop, sterile pH 7.2 PBS was used for rinsing. After washing, the crop epithelial cells were scraped with a slide and
S. cerevisiae or
A. oryzae spore solution was added. The state of attachment was confirmed under an optical microscope.
2.2. S. cerevisiae Fermented Products (SCFP) and A. oryzae Fermented Products (AOFP) Preparation and Characteristics
Then 108 CFU/ mL of S. cerevisiae was added to dilute the fluid mentioned above into sterilized wheat bran at a ratio of 1 mL dilute fluid per 3 g of wheat bran. The moisture of S. cerevisiae pre-fermentation wheat bran was adjusted to 60%. After that, 500 phytase units (FTU) of phytase were added to 30 g S. cerevisiae pre-fermentation wheat bran. S. cerevisiae pre-fermentation wheat bran (contain phytase) was incubated at 30 °C for 5 days, and placed in a stove at 50 °C for 1 day. The S. cerevisiae fermented product (SCFP) was stored in a −20 °C refrigerator for the other test.
The spore counts of
A. oryzae were diluted to 9 × 10
6 spores/mL with deionized water. The diluted fluid was added into sterilized WB at a ratio of 1 mL dilute fluid per 3 g wheat bran. The moisture of
A. oryzae pre-fermentation wheat bran was adjusted to 50%. After that, 500 FTU was added to 30 g
A. oryzae pre-fermentation wheat bran.
A. oryzae pre-fermentation wheat bran (contain phytase) was incubated at 30 °C for 7 days, and placed in a stove at 50 °C for 1 day. The
A. oryzae fermented product (AOFP) was stored in a −20 °C refrigerator for the other test. The xylanase [
15], protease [
16], cellulase, and ß-glucanase [
17] activities in SCFP or AOFP were analyzed before the animal experiment.
2.3. Peripheral Blood Mononuclear Cell Isolation
The assay was conducted and modified according to Kaiser et al. [
18]. Briefly, 5 mL whole blood was collected from a total of thirty-six 35-d-old broilers (3 for each pen, 9 for each treatment) in the Summax Single-use Containers for Human Venous Blood Specimen Collection (Lithium Heparin Tube), and centrifuged at 200×
g for 10 min to separate blood cells and plasma. The supernatant was replaced with sterilization PBS, and the sample was mixed uniformly. The mixed sample was added to the same amount of ficol, and centrifuged at 200×
g for 30 min. The separated PBMC were moved to the new RNase free Eppendorf tube (Gunster Biotech, Co., Ltd., Taipei, Taiwan), and washed by PBS for three times. After this, the sample was centrifuged at 200×
g for 10 min to remove the suspension PBS. RPMI-1640 (Merck, Darmstadt, Germany) with 10% fetal bovine serum (Merck, Darmstadt, Germany) (for cell test) or PBS (for qPCR) were used to re-suspend PBMCs and dilute it to 10
7 cells/mL.
2.4. Nitric Oxide Assay
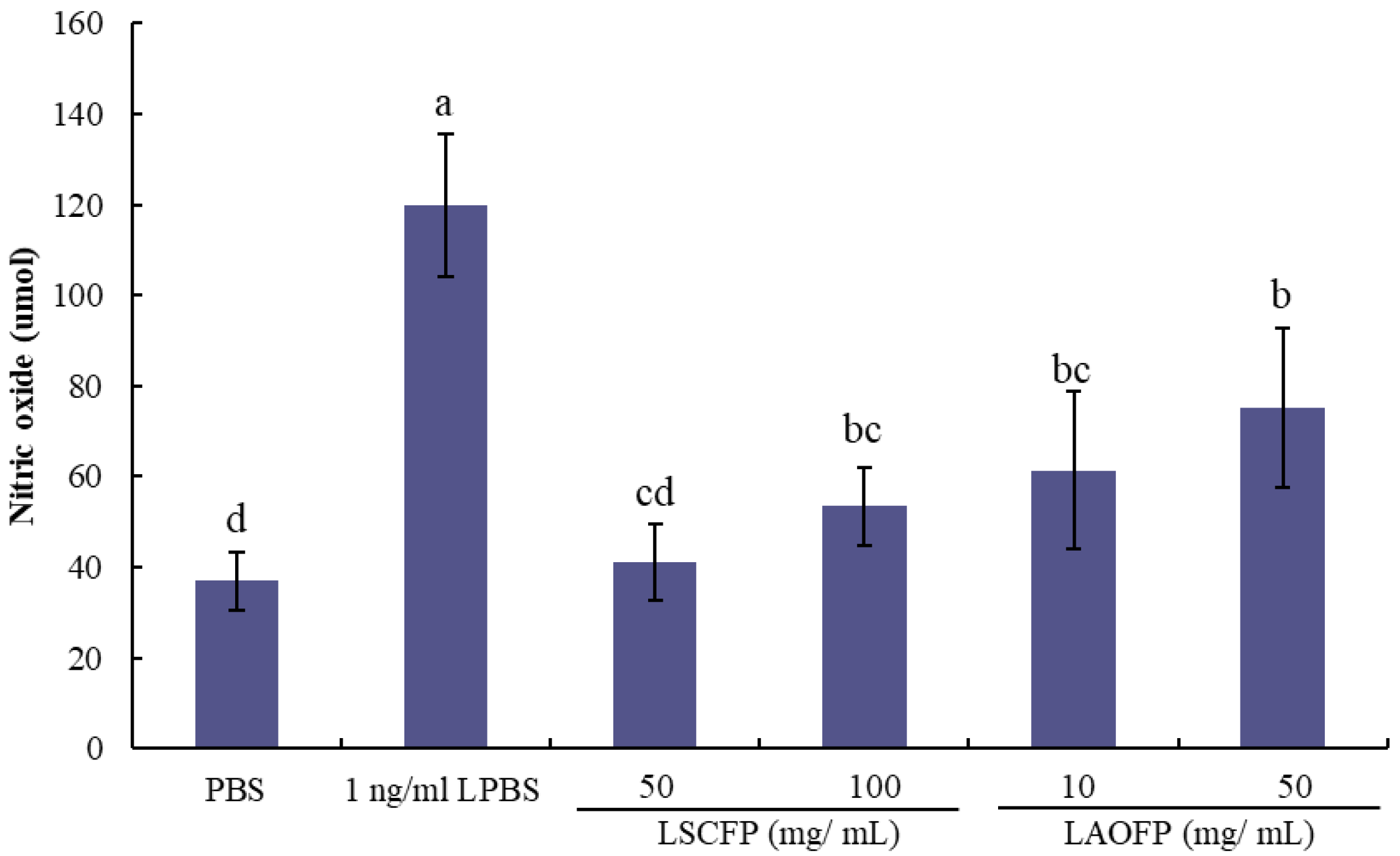
Isolation diluted PBMC (107 cells/mL) were cultured at 37 °C and 5% CO2 for 2 h. Then 10 μL 1 ng/mL lipopolysaccharides (LPS) and 10 μL sample solution, which had been filtrated by the 0.22 μm filter, were added. Sterilized PBS was used as the control group. After co-incubation for 24 h, 100 μL Griess reagent was added, and the absorbance was detected at 540 nm.
2.5. 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
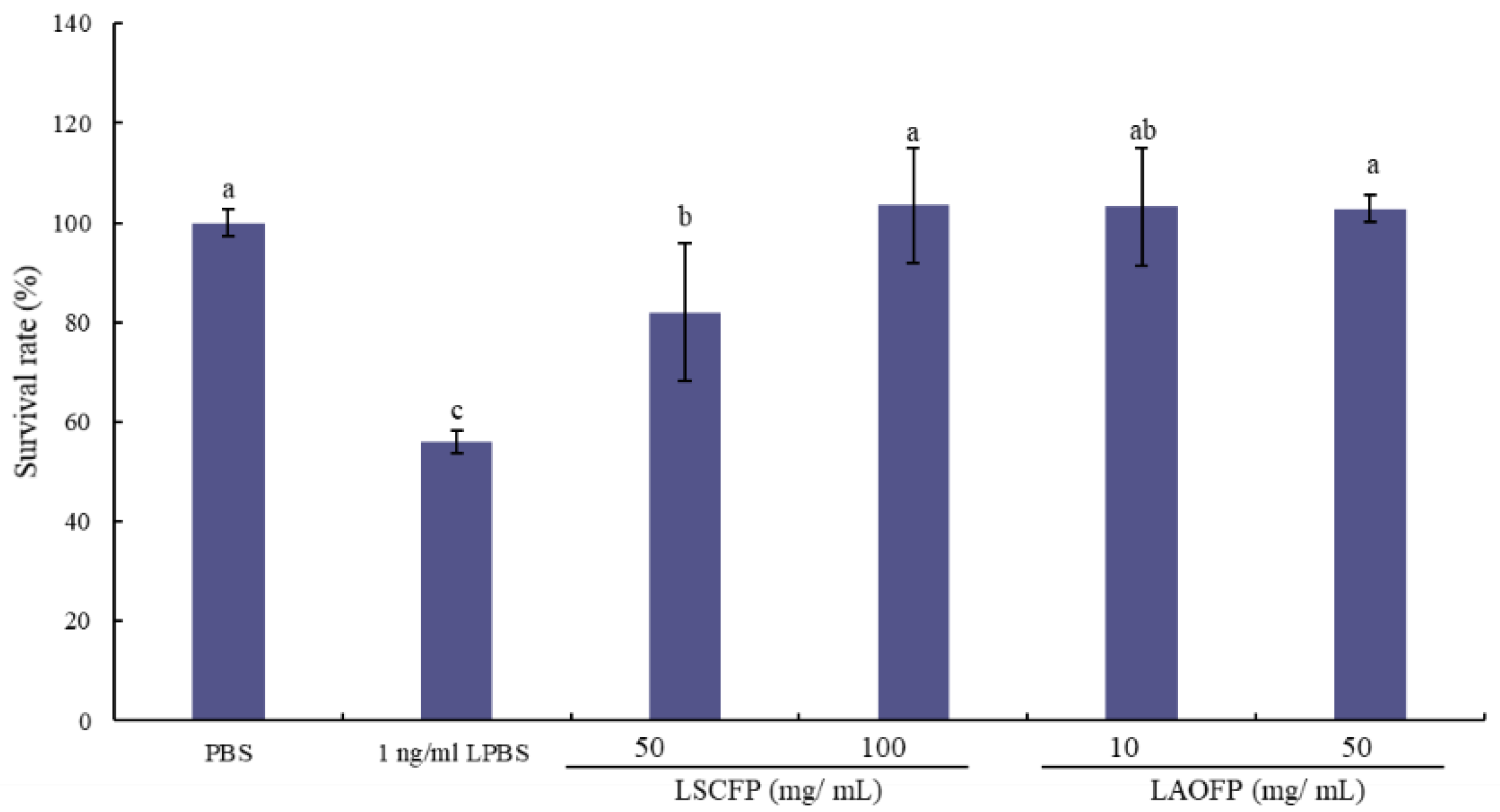
Isolation diluted PBMC (107 cells/mL) were cultured at 37 °C and 5% CO2 for 2 h. Then 10 μL 1 ng/mL lipopolysaccharides (LPS, Merck, Darmstadt, Germany) and 10 μL sample solution, which has been filtrated by 0.22 μm filter, were added. Sterilized PBS was used as the control group. After co-incubating for 48 h, 20 μL 0.5% MTT (Merck, Darmstadt, Germany) was added, and cultured for 4 h. After culturing, 100 μL dimethyl sulfoxide (DMSO, Merck, Darmstadt, Germany) was added, and the absorbance was detected at 570 nm.
2.6. Animal Experimental Design
To clarify the effect of SAFP, AOFP, and SAFP on broilers, the intestinal morphology, blood, and serum characteristics and mRNA expression in broilers’ PBMC were measured. This experiment was conducted at National Chung Hsing University, Taiwan. All of the protocols for animal use were followed by the Animal Care and Use Committee (IACUC: 107-013). The methods of animal design, intestinal morphology, blood and serum characteristics, and microbial parameter in intestinal content were slightly modified according to Teng et al. [
5]. A total of 240 male one-day-old broiler chickens (Ross 308) were used and divided into four groups: Control, 0.1% SCFP (SCFP), 0.05% SCFP + 0.05% AOFP (SAFP), and 0.1% AOFP (AOFP), respectively. Each pen had 20 broilers, and 3 replicate pens (a total of 60 birds per group). The average body weight (42.0 ± 0.5 g/bird) was similar among every pen initially. All birds were placed in the temperature control house. At 0–3 day-old, the temperature was 33 ± 0.5 °C, and then the temperature decreased as the broiler grew up. Temperature was controlled at 22 ± 1 °C after 30 days. Diets were divided into starter and finisher (
Table 1). Both of them met or exceeded the nutrient requirements of broilers (NRC, 1994) with addition of SCFP and AOFP. The proximate composition was analyzed according to the methods of Association of Official Analytical Communities (AOAC) [
19]. The starter and finisher diets were offered to the birds from 1–21 day-old and 22–35 day-old respectively. Body weight and feed consumption were recorded at 21 and 35 day-old. Body weight gain and feed conversion ratios (FCR) were calculated. Nine birds of each group were randomly selected to isolate PBMCs for qPCR and cell test.
2.7. Intestinal Morphology
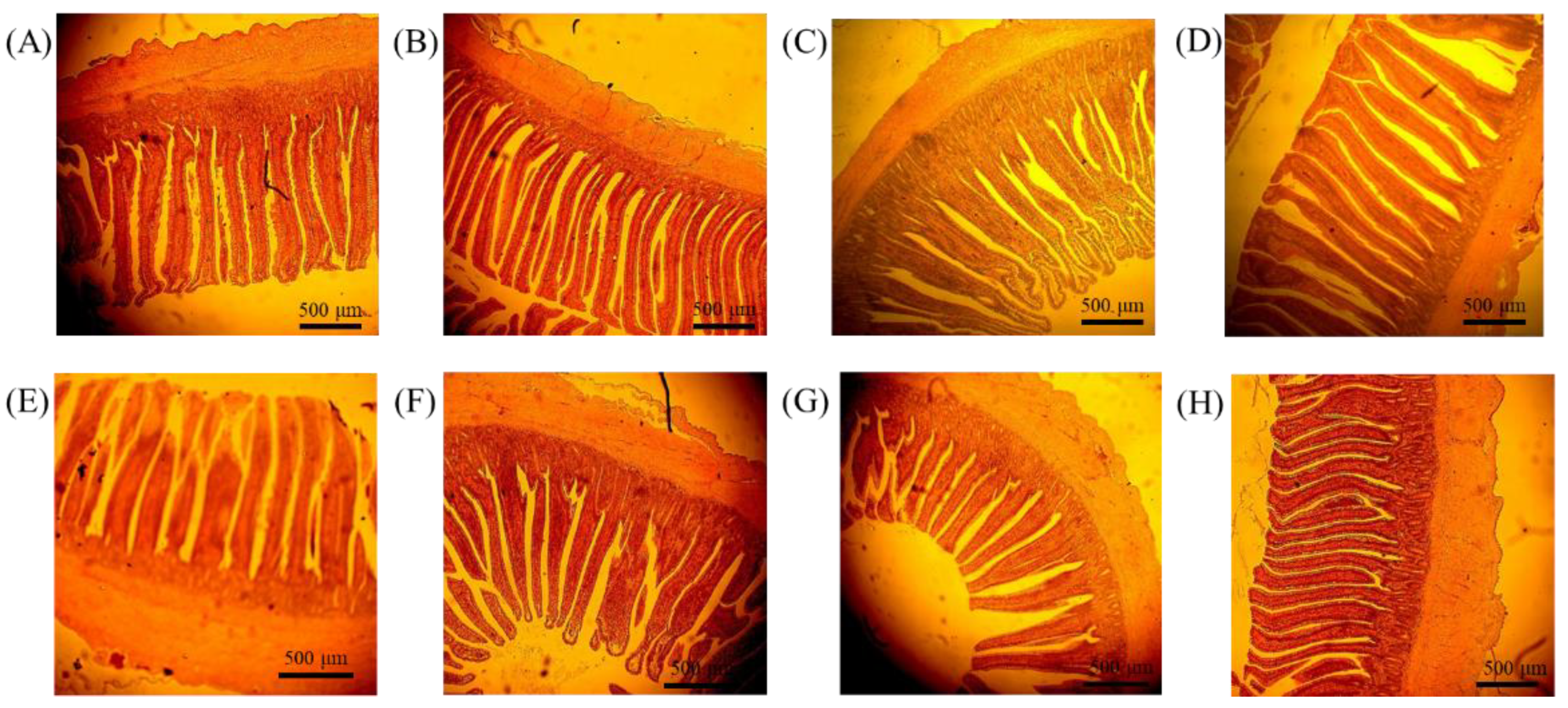
Twenty-four 35-day-old broilers (2 of each pen, 6 for each treatment, fasting for 1 day) were used for the measurement of intestinal morphology. The birds were euthanized, and the middle of the jejunum and ileum were excised, and the content was flushed by PBS and fixed in 10% formalin. Each sample was embedded by paraffin and stained with hematoxylin and eosin. The slice was observed with a light microscope. The villus height, crypt depth, and tunica muscular were measured by Motic Image Plus 2.0 analysis system (Motic Instruments, Richmond, BC, Canada).
2.8. Blood and Serum Characteristics
Thirty six 35-day-old broilers (3 of each pen, 9 for each treatment) blood was collected randomly for the blood and serum characteristic measurement. Blood samples stood for about 4 to 5 h at 4 °C, and then centrifuged at 3000 rpm for 10 min at 4 °C. Blood and serum biochemical parameters analyses were measured by automatic biochemical analyzer (Hitachi, 7150 auto-analyzer, Tokyo, Japan).
2.9. Microbial Parameter in Intestinal Content
Twenty-four 35-day-old broilers (2 of each pen, 6 for each treatment) were randomly selected for microbial analysis. Birds were euthanized, and the contents of ileum and cecum were squeezed out. Next, 1 g chyme was put into 9 mL PBS to sequence a dilution for the microbial parameter. Lactobacillus spp. were cultured with DeMan, Rogosa, and Sharpe agar (Difco™ Lactobacilli MRS Agar, BD, Franklin Lakes, NJ); Clostridium perfringens were cultured with tryptose sulfite cycloserine agar (GranuCult™ TSCagar, Merck KGaA, 64271, Darmstadt, Germany). Each microbe mentioned above was cultured at 37 °C under anaerobic conditions for 48 h. After culturing, the number of CFU on the agar was counted.
2.10. Total RNA Isolation and qPCR
Total RNA was isolated from PBMCs for determination of the mRNA expression level using SuperScript™ FirstStrand Synthesis System reagent (Thermo Fisher, Waltham, MA, USA) according to the manufacturer’s protocol. The method of determination of total RNA purity, cDNA synthesis, and qPCR analysis was slightly modified from Hu et al. [
20]. Briefly, 2× SYBR GREEN PCR Master Mix-ROX (Gunster Biotech, Co., Ltd., Taipei, Taiwan), cDNA, deionized water, and each primer were mixed at a ratio of 5:1.2:1.8:1. StepOnePlus™ Real-Time PCR System (Thermo Fisher, Waltham, MA, USA) was used to detect qRT-PCR performance, while the 2
−△△Ct method was used to calculate the relative mRNA expression level, and ß-actin was used as the housekeeping gene for normalization. Gene-specific primers are according to the genes of Gallus gallus (chicken) given as follows:
beta-actin (
ß-actin, 5′-CTGGCACCTAGCACAATGAA-3′ and 5′-ACATCTGCTGGAAGGTGGAC-3′, X00182.1);
interlukin-1 beta (
IL-1ß, 5′-GCTCTACATGTCGTGTGTGATGAG-3′ and 5′-TGTCGATGTCCCGCATGA-3′, NM_204524);
inducible nitric oxide synthase (iNOS, 5′-TACTGCGTGTCCTTTCAACG-3′ and 5′-CCCATTCTTCTTCCAACCTC-3′, U46504);
interferon-gamma (
IFN-γ (5′-CTCCCGATGAACGACTTGAG-3′ and 5′-CTGAGACTGGCTCCTTTTCC-3′, Y07922);
Nuclear factor kappa light chain enhancement of activated B cell p65 (NFκB p65, 5′-CCAGGTTGCCATCGTGTTCC-3′ and 5′-GCGTGCGTTTGCGCTTCT-3′, D13719.1).
2.11. Statistical Analysis
Data were analyzed for significance by analysis of variance (ANOVA) using SAS software (SAS® 9.4, 2018, SAS Institue Inc., Cary, NC, USA). Differences between treatment means were separated using Duncan’s multiple range test with p value less than 0.05.
4. Discussion
The high content of insoluble fiber and phytate could reduce the utilization of nutrition in animals. SCFP and AOFP contain quite a large amount of enzymes, such as cellulase, ß-glucanase, xylanase, and protease, thereby improving the nutrient utilization of broilers [
21]. Nevertheless, adding phytase to the fermentation process of SCFP and AOFP may also help to reduce the phytate content [
12]. From the results, the addition of SCFP and AOFP did not improve body weight, feed consumption, weight gain, and FCR. However, Santos et al. [
22] reported that phytase addition (500 FTU/kg) improved the body weight gain of 1–21 day-old broilers, but did not improve the body weight gain and FCR in 22–35 day-old broilers. Mountzouris et al. [
23] pointed out that
S. cerevisiae powder (50 mg (10
9 CFU)/kg diet) addition did not enhance the growth performance. Otherwise, there are few studies investigating the effects of
A. oryzae addition on broilers growth performance. As a previous study shows, replacing soybean meal by
A. oryzae fermented soybean meal (29.5% replacement in whole diet) can improve average daily gain and feed intake of broilers [
24]. As the replacement is far higher than in the current study, it is speculated that the addition of
A. oryzae should be sufficient to prove effective in growth performance.
The improvement of white blood cells (WBC) may be attributed to the joint effects of SCFP and AOFP because only SAFP increase WBC count in broilers blood. The SCFP and SAFP groups can significantly reduce the uric acid (UA) content in the blood, and the data number of SAFP is between SCFP and AOFP. This is because the protease yielded by SCFP was higher than AOFP and protease improved the protein utilization of the host, thereby decreasing the UA content in the blood. UA is a metabolite of protein that has an antioxidant function but is converted to a pro-oxidant in the cell or cytoplasm, and may be associated with cardiovascular disease when the concentration increases [
25]. In addition, UA also promotes inflammation and is associated with insulin resistance and metabolic disorders [
25]. Therefore, the addition of SCFP or AOFP to feed may prevent the development of cardiovascular disease, and reduce the oxidative stress of animal cells by the pathway of reduced UA.
In this study, villus height and the villus:crypt ratio in the jejunum were significantly increased and crypt depth significantly decreased on SAFP and AOFP, indicating that SAFP and AOFP may have an effect on improving nutrition absorption. One possible reason is that the enzyme yielded by SCFP and AOFP may degrade the anti-nutritional factor and make the energy utilization focus on villi growth and nutrient absorption [
26]. Kalantar et al. [
21] also reported that adding 0.1% commercial enzyme COMBO
® (including 1000 FTU/g phytase, 200 U/g xylanase, 200 U/g β-glucanase, and 80 U/g hemicellulase) in broilers diet can decrease the crypt depth and increase the villus length/width ratio and villus length/crypt depth ratio. Gao et al. [
27] showed that yeast supplement (2.5, 5.0, and 7.5 g/kg diet) can increase the villus height of the small intestine. Feng et al. [
24] found that replacing soybean meal by
A. oryzae fermented soybean meal can increase villus height on both duodenum and jejunum and decrease the crypt depth in jejunum. In addition, a decrease in the crypt depth indicates that the animal can spend less energy on the formation of the crypt, thereby reducing the energy loss [
26]. Furthermore, the results showed that Tunica muscularis decreased significantly on jejunum after adding SCFP, SAFP, and AOFP. Tunica muscularis consists of smooth muscle and is related to digestion and absorption of chyme. Cil et al. [
28] reported that inflammation causes the intestine to swell and increases Tunica muscularis weight as well as thickness. Therefore, by the anti-inflammatory effect of SCFP, SAFP, and AOFP confirmed by cell test and broilers PBMCs mRNA expression, the decreased thickness of Tunica muscularis is expected.
Previous studies pointed out that probiotic can reduce the number of harmful microbes in the gut [
5]. Mountzouris et al. [
23] indicated that
S. cerevisiae powder (50 mg (1 × 10
9 CFU)/kg diet) can protect broilers from
Salmonella Enteritidis (2 × 10
6 CFU/birds) challenge at day 15 and increase the growth performance of broilers. In this study, adding SCFP, SAFP, and AOFP can reduce
C. perfringens in the ileum. Among them, the SCFP is better than the AOFP based on the data, but with no significant difference, while the SAFP effect was in the middle. The antibacterial effect may be due to the growth of probiotics that crowd out the growth resources of
C. perfringens, or the glucan and mannan produced by the probiotics encapsulate the toxins yield from
C. perfringens and reduce the competitiveness of
C. perfringens. Furthermore, the number of
C. perfringens is positively correlated with the histochemistry score and necrotizing enterocolitis [
29], thereby decreasing the number of
C. perfringens may decrease the amount of pro-inflammatory cytokine.
LPS is an extracellular structure of gram-negative bacteria that can cause inflammation and stimulate the release amount of nitric oxide (NO) from PBMCs [
11]. However, SCFP and AOFP can protect PBMCs from LPS stimulus and decrease the NO production. The reason may be that the glucan and mannan in
S. cerevisiae and
A. oryzae cell wall acted as antitoxin [
3]. In addition, it is well-known that low-grade inflammatory response can protect the host from pathogens; however, high-grade inflammatory causes cell damage and apoptosis, so it is important to decrease the level of inflammation to prevent unexpected cell death. It is worth noting that AOFP has a similar effect as SCFP on lowering the concentration on NO production. Therefore, AOFP has similar effects as SCFP on lowering the concentration, which may represent that AOFP has a better anti-inflammatory effect than SCFP does.
For broilers, inflammation is a double-edged sword depending of its level [
11]. Lee et al. [
30] indicated that LPS challenge on broilers (1 mg/kg body weight) causes significant weight loss because of the immune response and decrease in feed intake. In broilers, there are two ways to decrease the inflammation level: one is to decrease the stimulants, such as LPS or bacterial toxin, and the other way is to block the pro-inflammatory cytokine pathway. From the results, SCFP and AOFP reduced the inflammation-related stimulant, including UA,
C. perfringens, and NO production from LPS stimulated PBMCs.
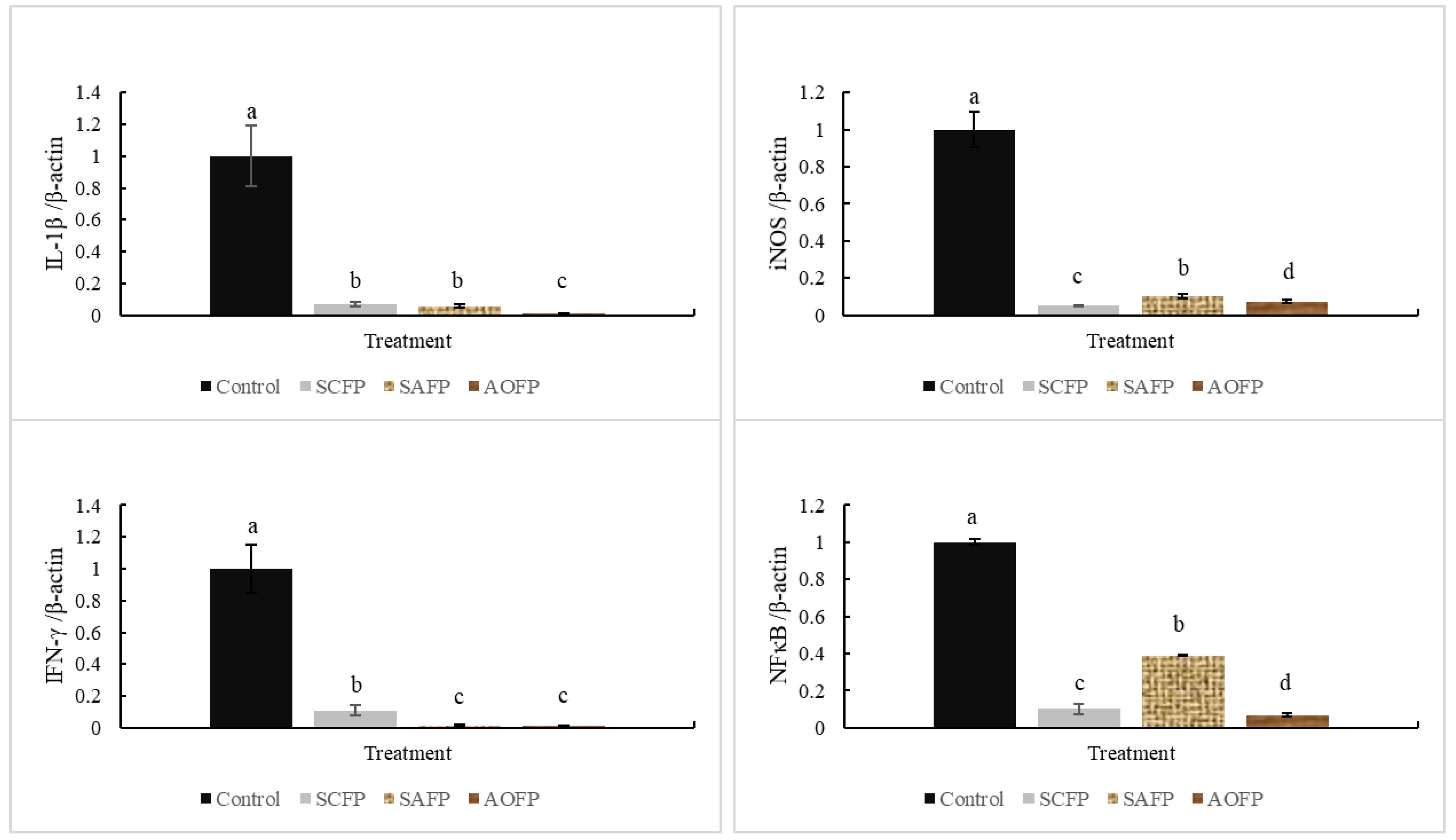
As a pro-inflammatory cytokine,
IL-1ß mediates many pathways involved in apoptosis or inflammation [
31]. It is well-known that
S. cerevisiae and
A. oryzae cell wall are rich in β-glucans [
3], which can inhibit
IL-1ß production [
32] and thereby reduce
NF-κB-mediated inflammatory responses [
33]. Therefore,
IL-1ß is positively correlated with
NF-κB.
NF-κB is a main inflammatory factor, which promotes reactive nitrogen species (RNS) content [
11]. In normal conditions,
NF-κB binds with the inhibitory protein named inhibitor of kappa B (IκB) and has no effect [
33]. With stimulants such as heat stress or infection,
IκB phosphorylates and releases the
NF-κB as a nuclear transcription factor to induce an inflammatory response [
33]. SCFP, SAFP, and AOFP addition can decrease the
IL-1ß and
NF-κB mRNA expression. Hegazy and Bedewy [
9] had similar results, indicating that 1 × 10
10 CFU of
Lactobacillus delbruekii and
L. fermentum can decrease the interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and
NF-κB expression. The results show that probiotics reduced inflammation by suppressing the
NF-κB and
IL-1ß expression. Furthermore, the expression of
iNOS can increase NO production, and is associated with infection [
34], while SCFP and AOFP reduced the
iNOS mRNA expression.
NF-κB can induce
iNOS expression, and increase the RNS production [
35]; therefore, decreasing the
NF-κB content can also suppress
iNOS expression.
IFN-γ is associated with infection and high concentration of
IFN-γ may contribute to autoimmune disease [
36]. When animals suffer from infection,
IFN-γ expression increases and induces
NF-κB expression to produce reactive oxygen species (ROS) and RNS to resist pathogens [
37]. The data shown above indicate that SCFP and AOFP can decrease the damage caused by LPS and the number of
C. perfringens in ileum, which are both related to the infection. Therefore,
IFN-γ suppressed by SCFP, SAFP, and AOFP in broilers also decreases the
NF-κB expression and reduces the inflammatory response [
37]. Decreasing the stimulant and blocking the inflammation-related mRNA expression mentioned above confirm that both SCFP and AOFP addition can reduce the inflammation, especially AOFP.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}