3.1. Variation of Infrared Body Surface Temperature by Treatment and Event
As the treatment × anatomical site interaction had no effect on IR body surface temperature (
P > 0.05) in this study, data were pooled across anatomical sites, and only the combined effects of handling type and number of laps by the event are presented and discussed (
Table 2).
A greater IR maximum body surface temperature was observed in GH pigs than in RH pigs at loading (32.52 ± 0.24 vs. 30.98 ± 0.24 °C;
P < 0.001), whereas at unloading, the RH group showed greater IR maximum body surface temperature compared to GH pigs (33.83 ± 0.25 vs. 33.09 ± 0.25 °C;
P = 0.04). The increased IR body surface temperature observed in GH pigs during loading may result from their psychological response to the novelties of the handling test course, since the gentle handling applied to this group allowed them to explore the novel environment and to react more to it compared to RH pigs (see later section on behavior results). Whereas, IR body surface temperature variation observed in RH pigs likely resulted from their fatigue caused by the additive effect of the handling intensity during loading and transport stress, as reported in a number of previous studies [
36,
37].
Except for IR body surface temperature recorded on pigs’ return to the home pen, the interaction handling type × number of laps had no effect on IR body surface temperature at any time in this study (
Table 2). On the return to the home pen, the IR body surface temperature of RH3 pigs was greater (+2.24 °C;
P = 0.02) than that of RH1 pigs. This result may be explained by the additive effect of the greater handling intensity and longer walking distance on the thermal condition of RH3 pigs. The increased IR body surface temperature in the pigs likely resulted from the greater heat produced by skeletal muscle activity [
38,
39,
40,
41,
42], cutaneous vasodilatation [
43], and short-term exchange in the convective delivery of central heat to the periphery [
44].
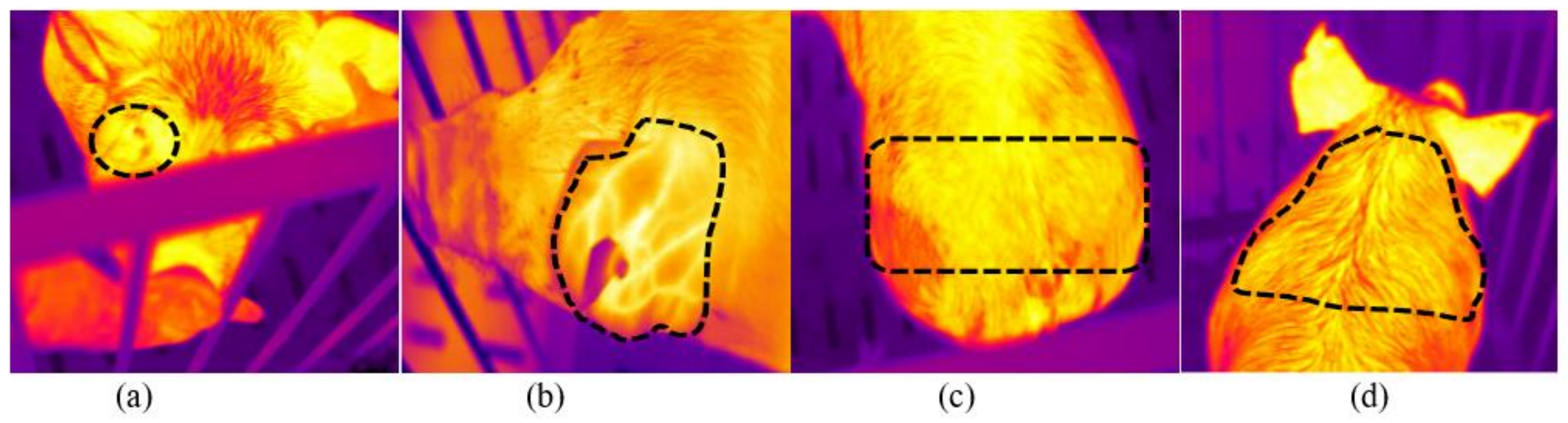
3.2. Variation of Infrared Body Surface Temperature by Anatomical Site and Event
As shown in
Table 3, IRNT and IRBET progressively increased from rest to loading and from loading to unloading (
P < 0.001 for both). In this study, IROT also increased from rest to unloading, but dropped on the return of pigs to the home pen (
P < 0.001). Whereas, IRRT increased from the rest to loading, dropped at unloading and increased again upon the return of pigs to the home pen (
P < 0.001). The temperature variation observed on the pig’s head may result from the effects of physical stress of loading and unloading on body temperature, as reported in previous studies [
36,
37], and the greater heat dissipation through blood circulation from the head [
4]. The lower temperature recorded in the rump region at unloading may be explained by the reduced heat dissipation, due to the insulation provided by the thicker fat cover in this region [
45].
As shown in
Table 3, among the anatomical sites, the greatest variation in IR skin surface temperature between rest and unloading was observed in the orbital region (IROT ΔT = +7.01 ± 0.29 °C;
P < 0.001) and behind the ears (IRBET ΔT = +5.86 ± 0.46 °C;
P < 0.001). The second largest variation between the rest and the return to the home pen IR values was recorded in the neck region (IRNT ΔT = +5.73 ± 0.30 °C;
P < 0.001) and behind the ears (IRBET ΔT = +5.67 ± 0.33 °C;
P < 0.001). Compared with IRNT, the lower variation in IRBET values may be due to the lower skin and subcutaneous fat thickness in the region behind the ears compared with the neck regions [
45]. Whereas, the greater hair density on the neck and rump surface compared to the orbital region and the ears [
45] may be a factor contributing to lower variation in IR temperature in these anatomical sites. A dense, dry hair coat, in fact, increases the emissivity of the skin area, because of the cavity effect [
46,
47].
In this study, the highest IR temperature was recorded behind the ears (IRBET) and in the orbital (IROT) regions at unloading (
P < 0.05;
Table 3)—which may be explained by the close proximity of the eyes and ears to the brain [
48], and their richer capillary beds innervated by the sympathetic system quickly responding to changes in the blood flow in stressful situations [
23,
49], the greater vasomotor response in the extremities than in the middle body regions [
50], and the greater heat dissipation in the region of the head [
16,
18,
19,
51].
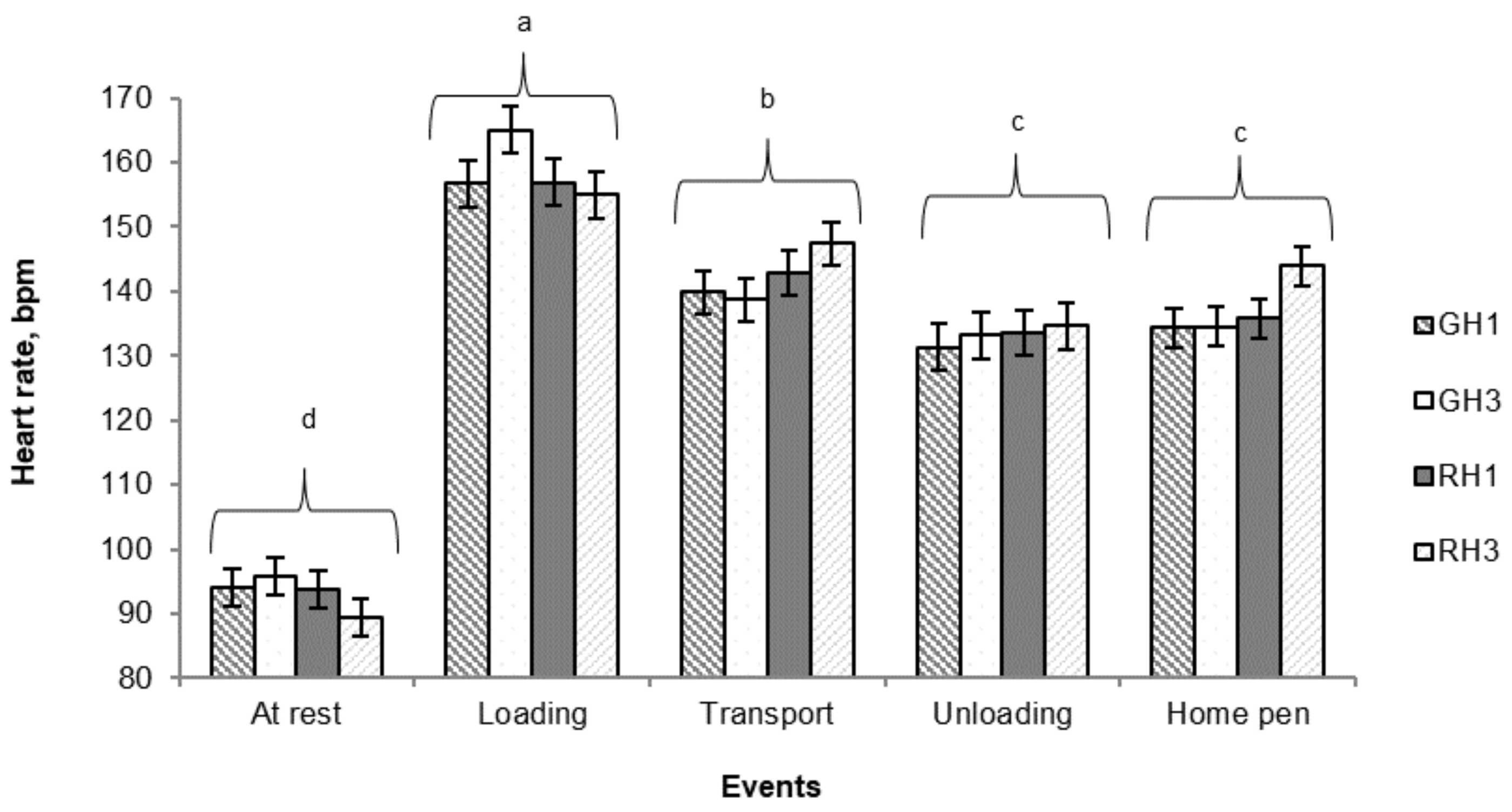
3.3. Heart Rate
Overall, in this study, pigs’ heart rate varied between events (
P < 0.001;
Figure 4), with the rate is the highest at loading (158 ± 1.56 bpm) and dropping during transport (142.4 ± 1.7 bpm), at unloading (133.0 ± 1.7 bpm) and up to the return at the home pen (137.0 ± 1.5 bpm). However, no significant change in heart rate was found in pigs between unloading and the return to the home pen (
P > 0.05).
As there was no interaction between handling type and number of laps, heart rate data were pooled across treatments (
Table 4). Heart rate was only affected by handling type, with RH pigs showing a greater (
P = 0.04) heart rate on their return to the home pen compared with GH pigs. No significant effect of the number of laps on pigs’ heart rate was observed in this study (
P > 0.05).
In agreement with a number of previous reports [
37,
52,
53,
54,
55], in this study, heart rates increased at loading and decreased during transport and at unloading. However, in this study, this effect was exacerbated by the additive effects of rough handling through an alley characterized by multiple corners and a sloped ramp. Increased heart rates have been previously reported in pigs handled with electric prods [
53], negotiating closed corners or bends [
56] and sloped ramps [
57,
58,
59] at loading.
3.4. Variation of Rectal Temperature by Event
No significant effect of the handling type or number of laps, either as a single factor or interaction, was found on rectal temperature during the events (
P > 0.05;
Table 5). This result is difficult to explain, since rectal temperature has been commonly used as an indicator of stress in several studies [
60,
61,
62]. In this study the lack of effect of the handling intensity or the distance moved on the rectal temperature may be related to the time lapse between unloading, driving pigs back to the home pen and rectal temperature measurement (approx. 10–15 min.). However, other studies also failed to report an effect of handling intensity or distance moved on the rectal temperature variation in pigs [
60,
63]. Additionally, it cannot be ruled out that the size differences between animals used in this study and those reported studies may be an important factor for the lack of results.
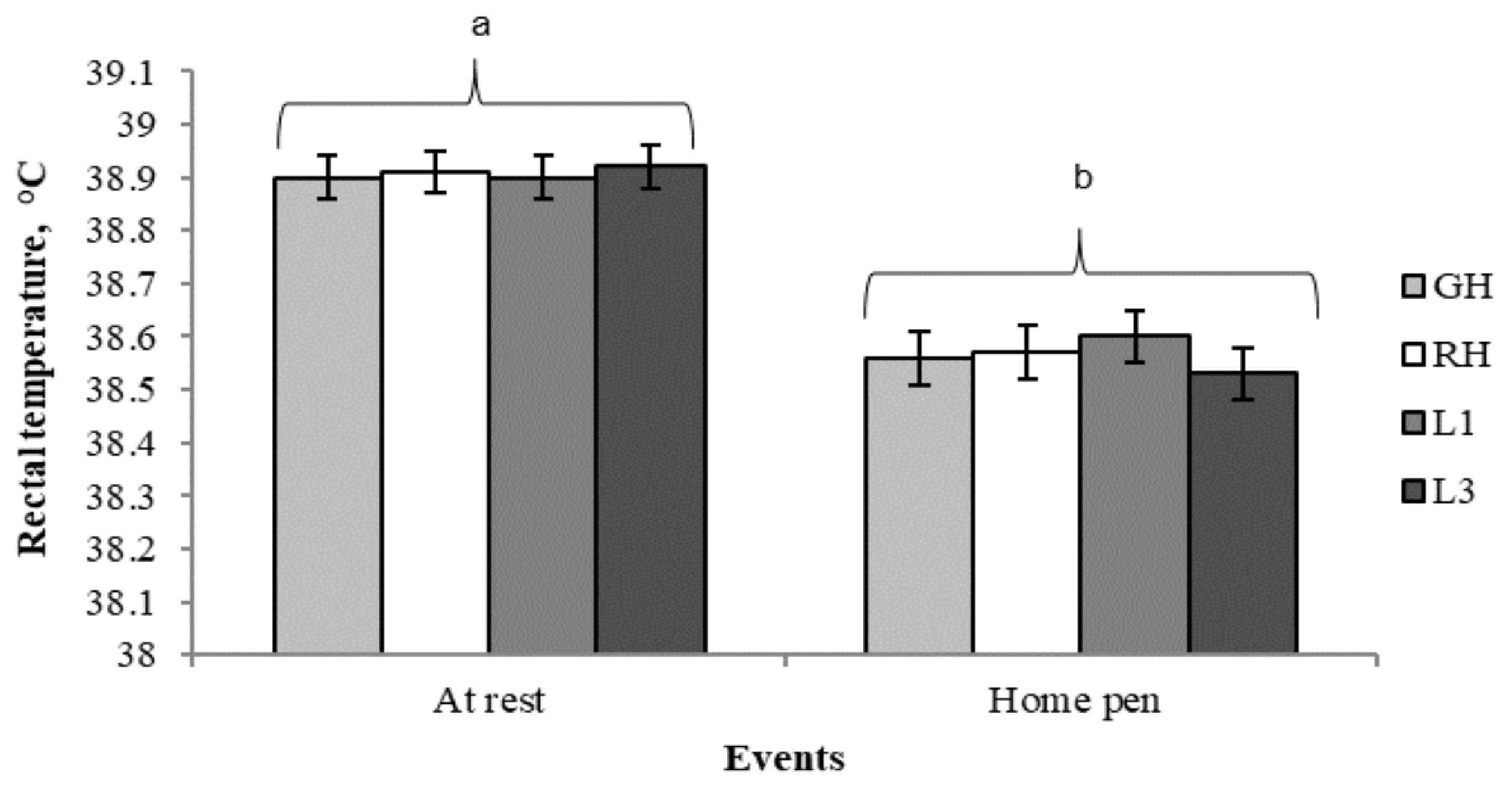
Therefore, data were pooled across treatments to show the variation of rectal temperature between rest and the end of the handling/transport test (
Figure 5). The average rectal temperature was slightly greater at rest than on pigs’ return to the home pen (38.9 ± 0.03 °C vs. 38.6 ± 0.03 °C;
P < 0.001), however, both values are within the reference range reported for slaughter pigs (39.3 ± 0.03 °C; [
14]). Additionally, the higher rectal temperature at rest may result from the relatively high basal blood flowing from hotter visceral structures to the rectum, making it, along with the gastrointestinal tract, the body site with the highest core temperature [
64].
3.6. Behavioral Response
In this study, GH pigs took longer to load than the RH pigs (4.03 ± 0.53 vs. 1.47 ± 0.05 min;
P < 0.001). Correa et al. [
53] also reported a shorter loading time for pigs handled harshly (i.e., electric prodding) compared with those driven gently (i.e., with paddles) as they are imposed to walk at a fast pace. As expected, in this study driving pigs through three laps instead of only one lap of the handling test course took longer (3.71 ± 0.47 vs. 1.79 ± 0.26 min;
P < 0.01).
As there was no interaction between handling type and number of laps on loading time and pig behaviors, data were pooled across treatments (
Table 7).
Similar to Correa et al. [
53] and Rabaste et al. [
66], in this study rough handling resulted in more pigs overlapping compared with more gentle handling (
P = 0.05;
Table 7). However, when compared with RH pigs, GH pigs backed-up, turned back and vocalized more, and were more reluctant to move (
P < 0.001). These behaviors may indicate that when gentle handling is applied, pigs are more inclined to walk at a pace that allows them to explore the novel environment and express fear behaviors, such as reluctance to move and turning-back [
67], towards the novelties encountered through the handling test course (i.e., ramp, bends and sharp corners in this study). Goumon et al. [
58] also reported a greater frequency of fear behaviors in near-market-weight pigs being handled through ramps or corners.
When compared to one lap, imposing pigs to walk three laps through the handling test course increased the frequency of physical contacts from the handler (
P = 0.05), reluctance to move (
P = 0.04) and turns-back (
P < 0.001). The physical effort to go through a longer walking distance and to go up and down a ramp three times may elicit increased frustration and fatigue in this group of pigs, leading to the higher incidence of those fear behaviors, and the need of more handler interventions to move forward. Previous studies [
68,
69,
70] also observed signs of physical signs of stress during loading in pigs moved over a long distance compared with those moved over a short one.
Overall in this study, pigs only stood and sat during transport, regardless of handling type or number of laps (
P > 0.05;
Table 8). These postures are normally observed in the truck during trips as short as that of this study [
34,
71,
72]. Pigs, in fact, start to accommodate in the truck and lie down only between 0.5 and 3 h from the departure from the farm, regardless of the loading conditions [
73,
74,
75].
3.7. Correlations between IR Temperature at Different Anatomical Sites, Other Physiological Stress Indicators and Behavior Measurements
Spearman correlation coefficients between IRNT, IRRT, IROT and IRBET, and physiological measurements are shown in
Table 9. Among the anatomical sites, the greatest correlation was found between IROT and IRBET (
r = 0.90;
P < 0.001), followed by that between IROT and IRNT (
r = 0.84;
P < 0.001) and IRNT and IRBET (
r = 0.82;
P < 0.001). Previous studies also reported strong correlations between IR temperature values at these anatomical sites in pigs [
19,
51].
The greater magnitude of the correlation between IROT and IRBET may be explained by the greater heat dissipation through blood circulation from the head [
44]. Additionally, according to Ng et al. [
76], the accuracy of surface temperature measurements at the orbital and ears locations is even better compared with other anatomical regions, due to the lower fat thickness and hair coat density reducing the distance between the blood flow and the body skin in these regions. This difference in measurement accuracy between anatomical sites was also observed in our study, where low correlations were observed between IROT or IRBET and IRNT regions.
In this study, all correlations between physiological stress indicators and IR temperature values at different anatomical sites were significant (
P < 0.001), except for IRNT and heart rate (
r = 0.13;
P > 0.05), but were from weak to moderate (
r = −0.27 to 0.56). The greatest correlation was found between IROT and heart rate (
r = 00.56;
P < 0.001), which is not surprising as heart rate and IR ocular temperature are both indicators of physical stress [
22,
58]. Greater correlations were also found between IR temperatures at all anatomical sites and salivary cortisol (
r = 0.49–0.51;
P < 0.001). Similar or higher correlations (
r = 0.55 to 0.71) between the IR maximum ocular temperature and salivary cortisol have also been reported in horses [
77,
78]. However, our results are in disagreement with those reported by Warriss et al. [
17] who failed to find a relationship between IR ear temperature and serum cortisol level. The discrepancy in the results between the two studies may be explained by the greater responsiveness of salivary cortisol to stress-related adrenocortical activity compared to plasma cortisol [
77]. Therefore, based on its association with salivary cortisol variation, it may be concluded that the variation in the IR body surface temperature—especially that assessed in the head region, may also be influenced by the hypothalamic-pituitary-adrenal axis activity in response to the physical stress or exercise during handling and transport in pigs, leading to an alteration in heat production and loss from the animal [
3].
Significant, although rather weak, the correlation was found between IROT and rectal temperature (
r =−0.32;
P < 0.001). This result suggests that a greater variation in IROT during physical effort may be explained by the different dissipation rate between core, i.e., rectal temperature, and superficial temperatures (i.e., orbital IR skin surface temperature) resulting from the difference in blood flow redistribution during physical effort. In this study, IROT temperature increased from rest to loading (29.05 ± 0.26 °C vs. 36.06 ± 0.26 °C), while rectal temperature (38.9 ± 0.03 °C vs. 38.6 ± 0.03 °C) remained stable. Indeed, this result indicates that during physical effort the sympathetic nervous system is activated and catecholamines are released into the bloodstream resulting in vasodilation and increased blood flow in the region of the head [
79]. As the blood flow changes in response to physical activity, the amount of radiated heat that is lost from the orbital region is increased, whereas the parasympathetic system reduces gastrointestinal activity by decreasing blood flow towards the intestinal tract [
80], likely caused by the vasoconstriction of the rectal wall in contrast to the orbital region [
81].
No significant correlations between IR body surface temperature at different anatomical sites and behavioral measurements were found in this study (P > 0.05).


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}