Fossil History of Curculionoidea (Coleoptera) from the Paleogene


Abstract
:1. Introduction
2. Materials and Methods
3. Results
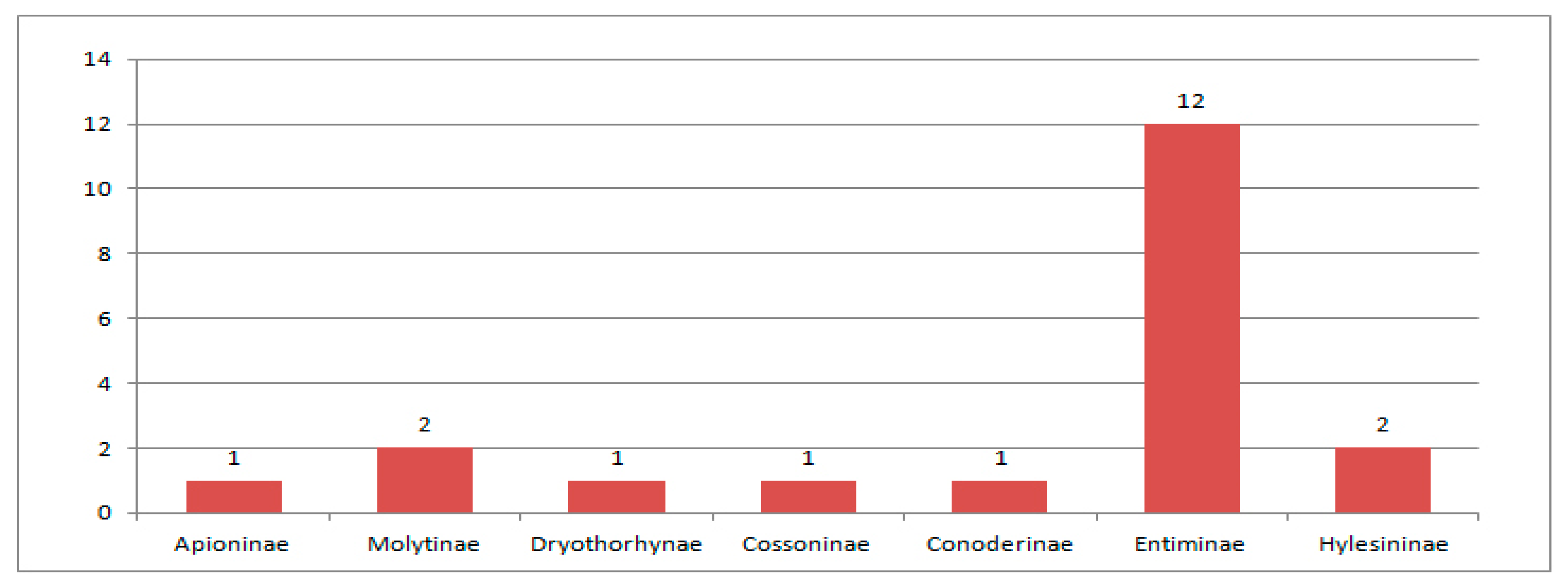
3.1. Paleocene Weevil Fauna
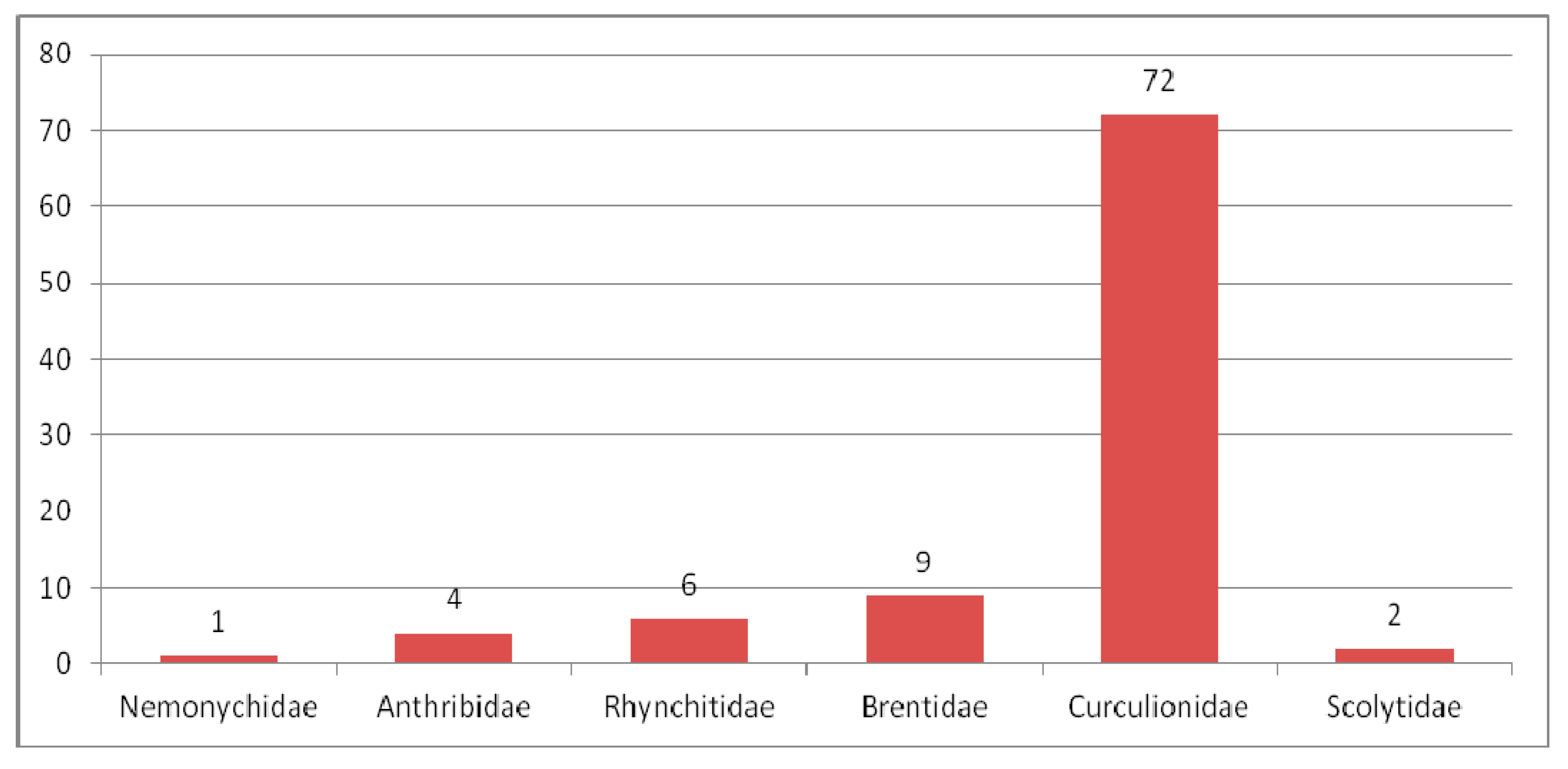
3.2. Eocene Weevil Fauna
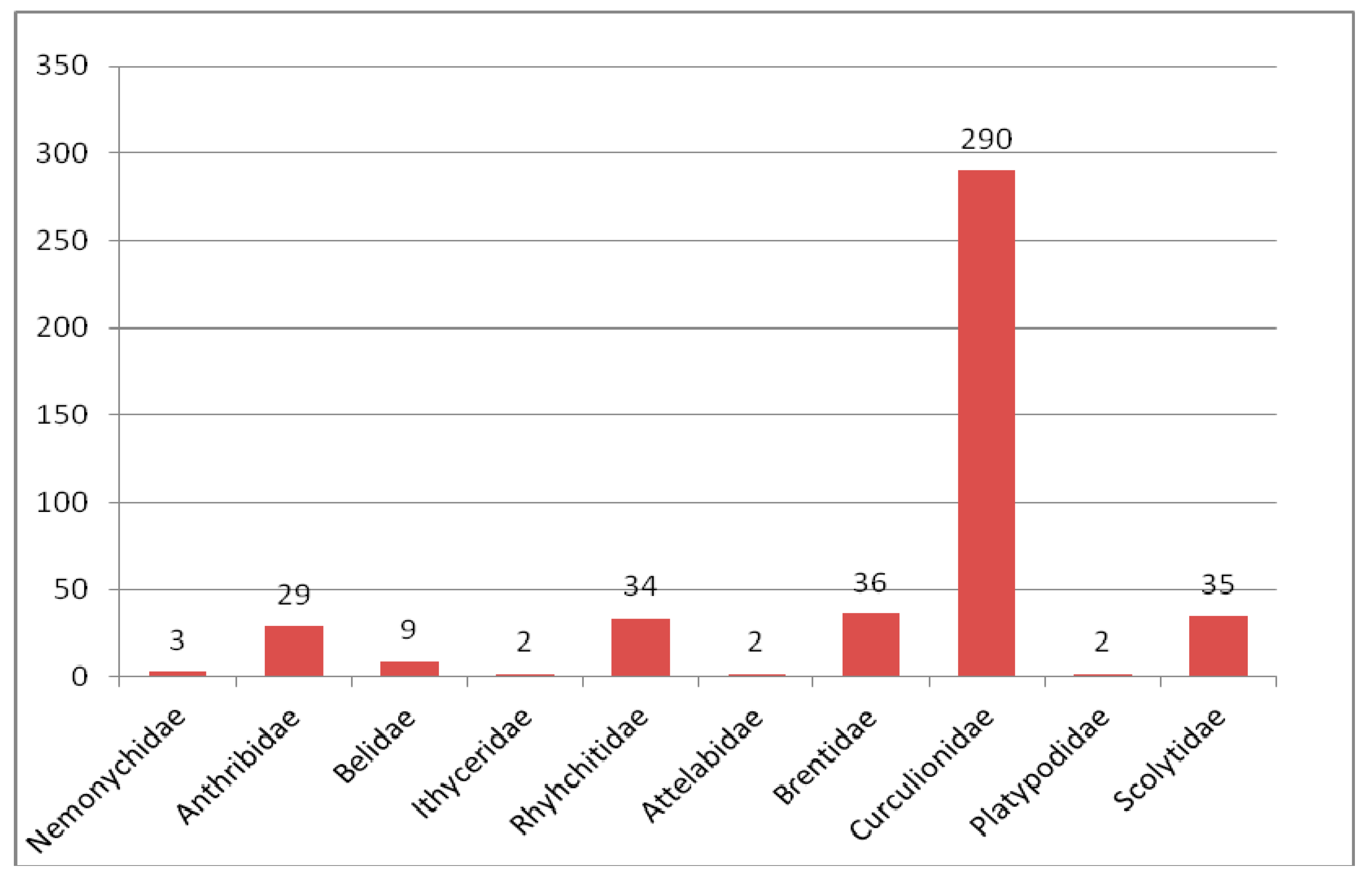
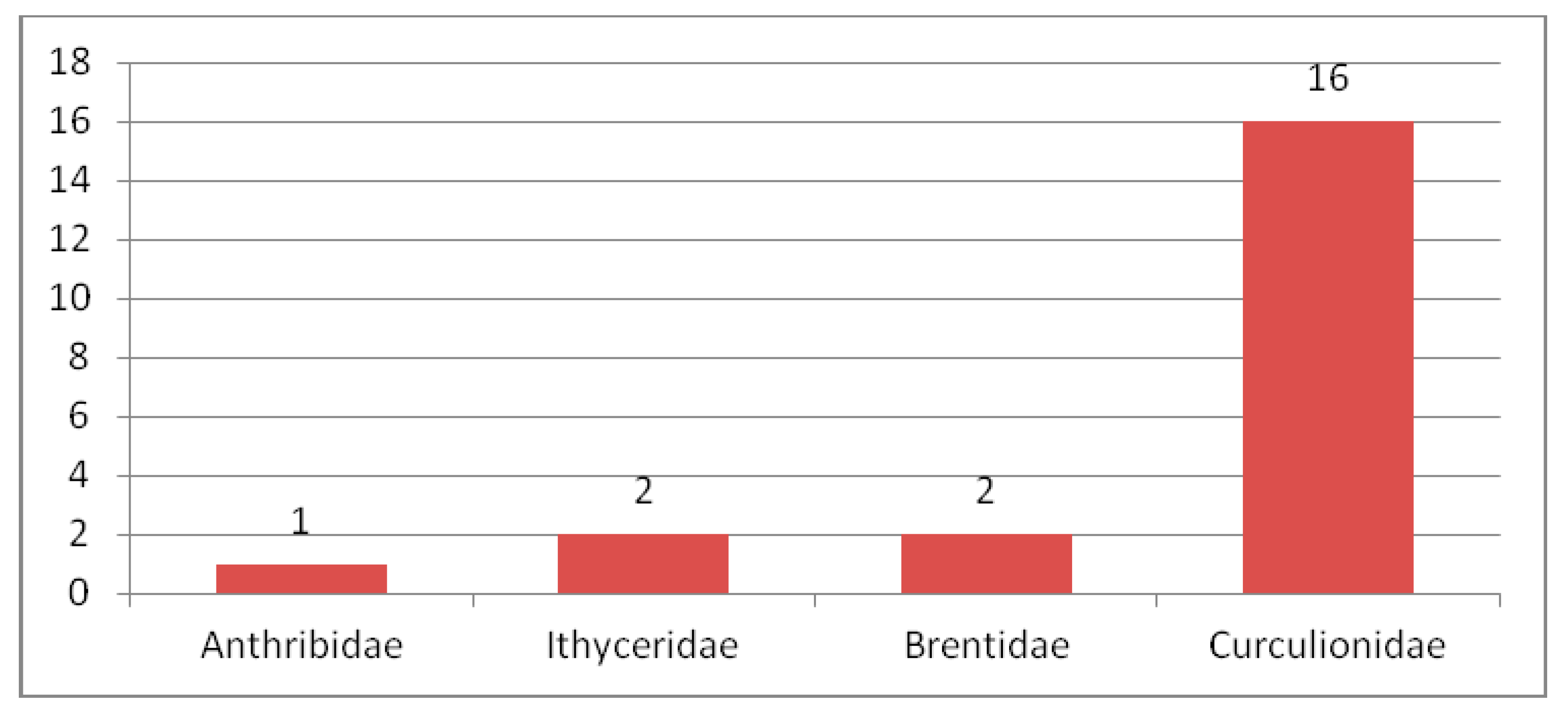
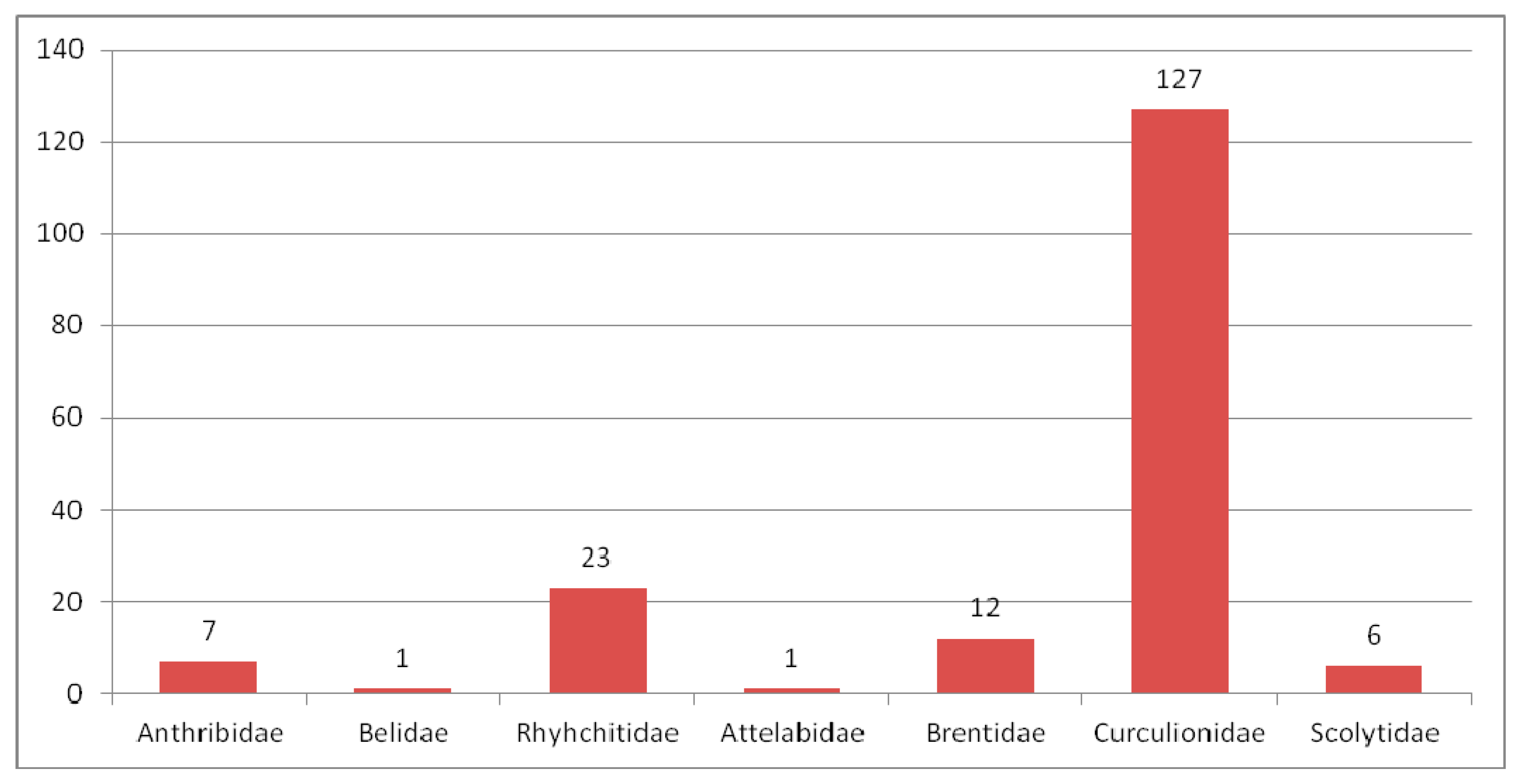
3.2.1. Review of Curculionoidea Families in the Eocene
Nemonychidae in the Eocene
Anthribidae in the Eocene
Ithyceridae in the Eocene
Belidae in the Eocene
Rhynchitidae in the Eocene
Attelabidae in the Eocene
Brentidae in the Eocene
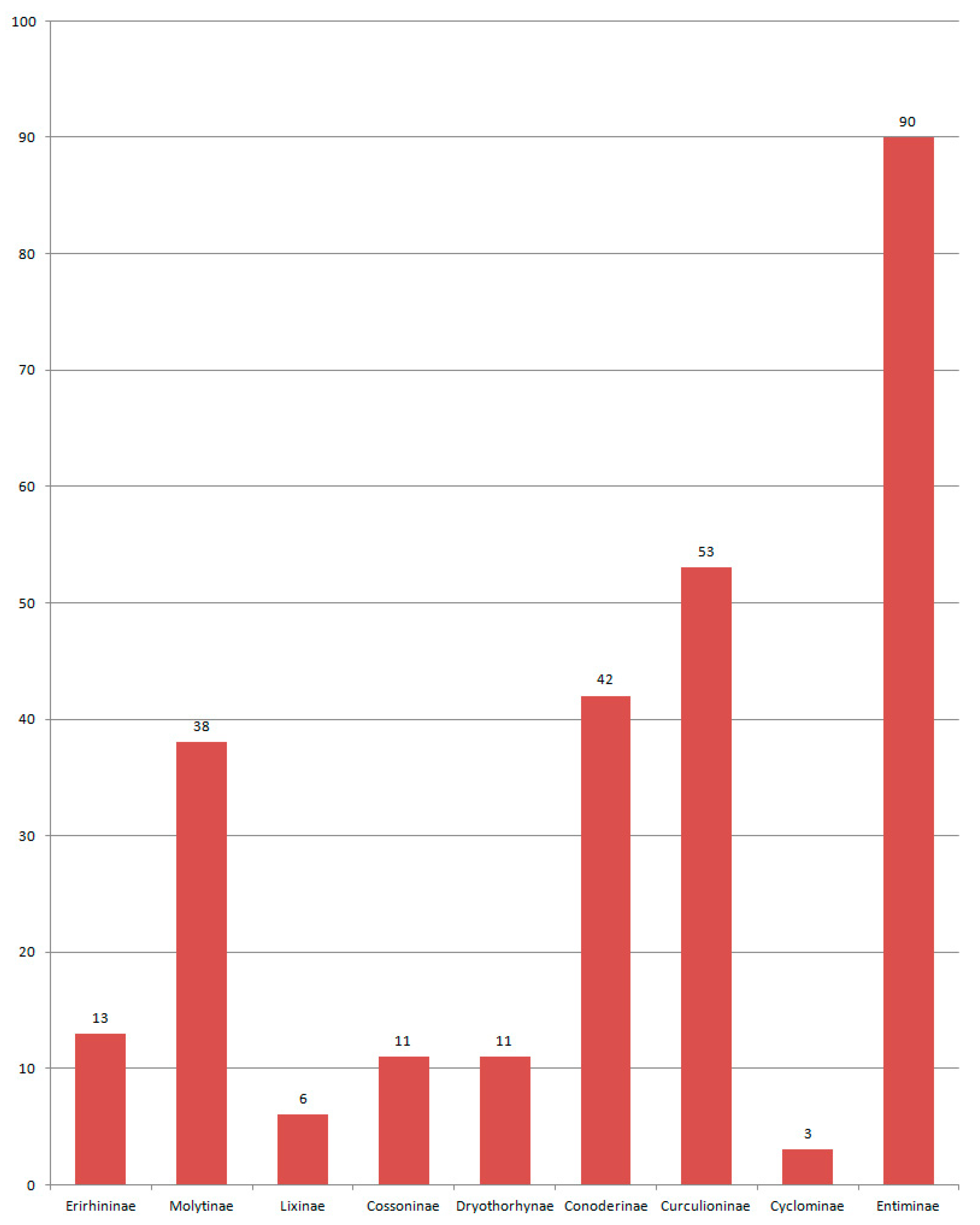
Curculionidae in the Eocene
Platypodidae in the Eocene
Scolytidae in the Eocene
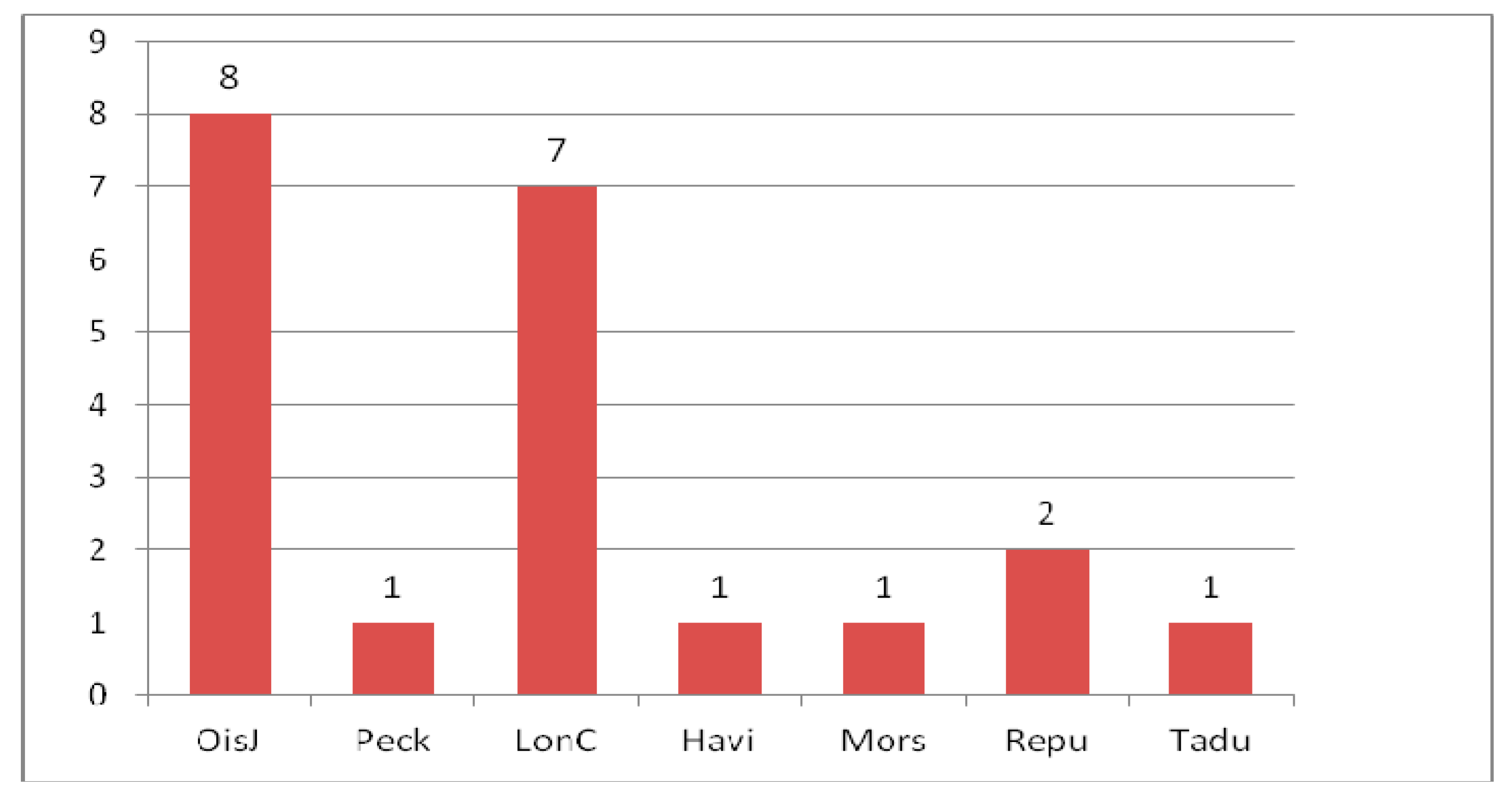
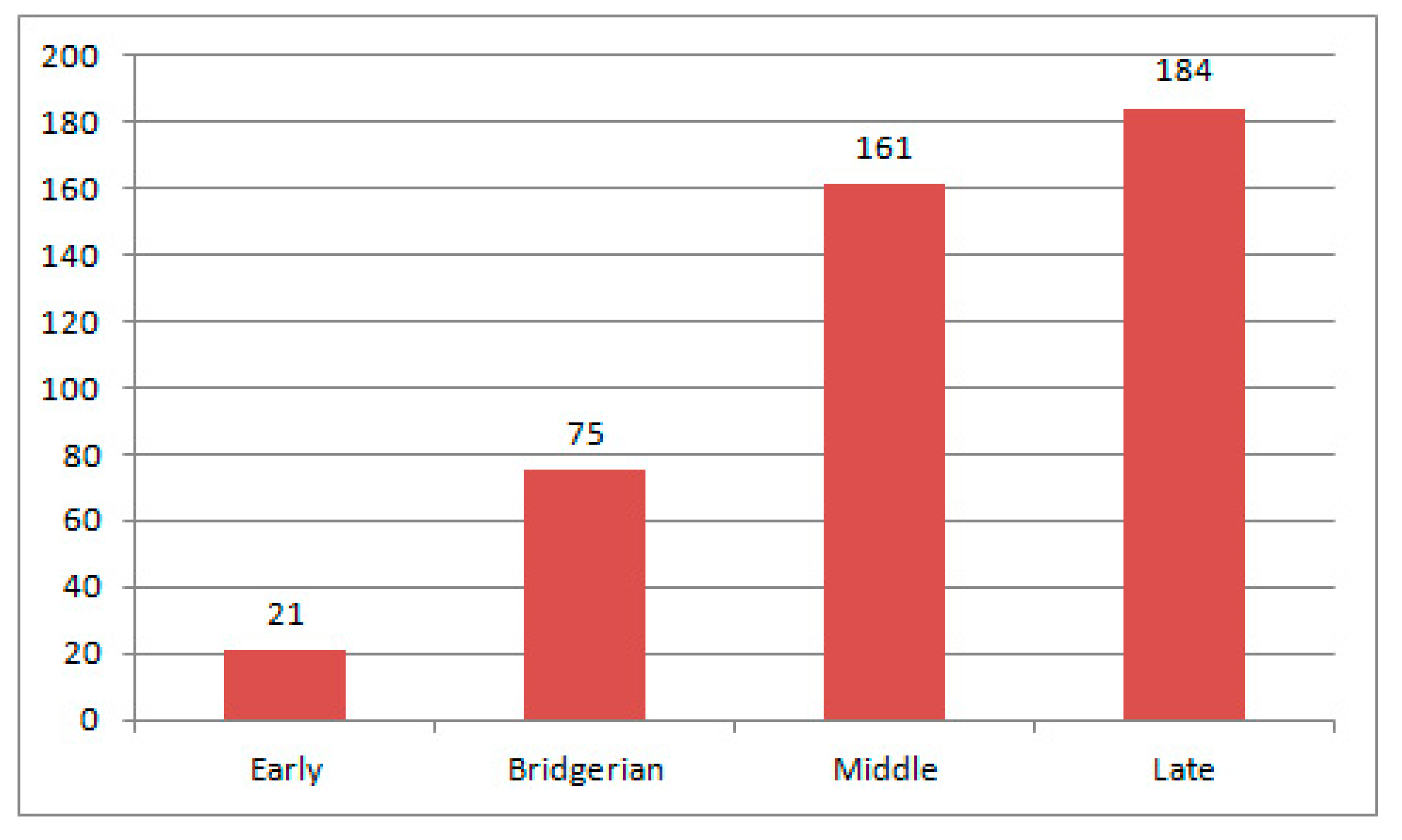
3.2.2. Early Eocene Weevil Fauna
3.2.3. Bridgerian Weevil Fauna
3.2.4. Middle Eocene Weevil Fauna
3.2.5. Late Eocene Weevil Fauna
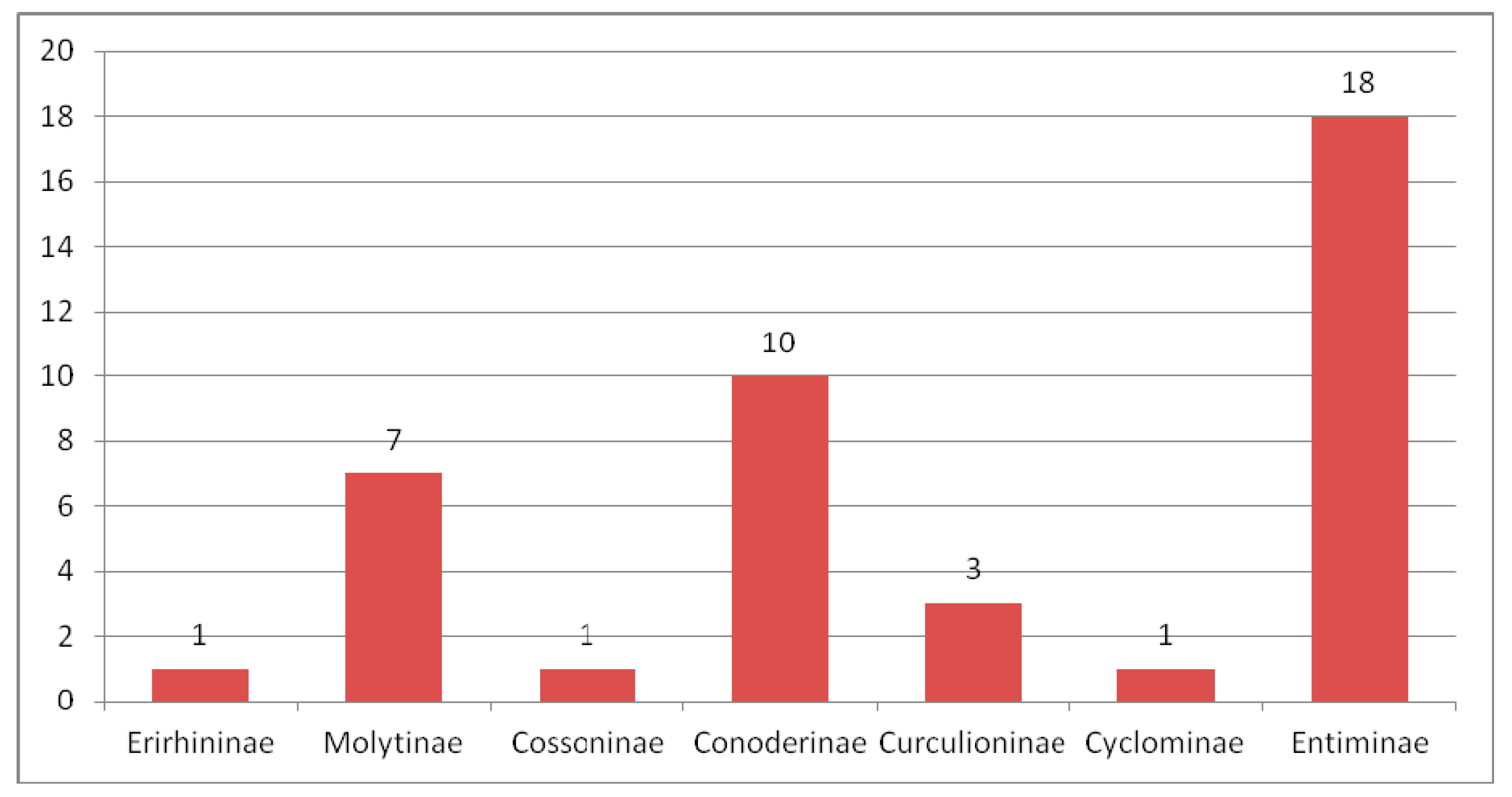
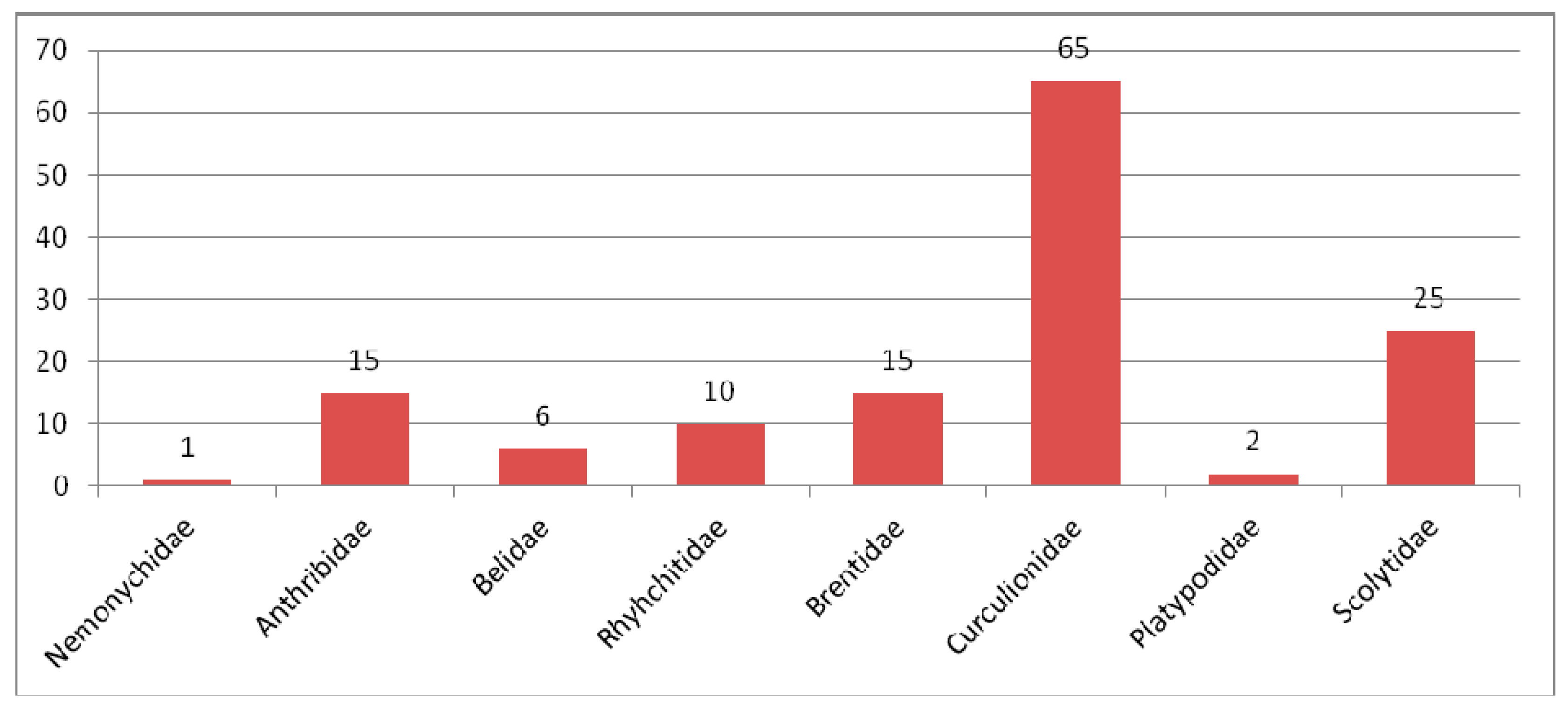
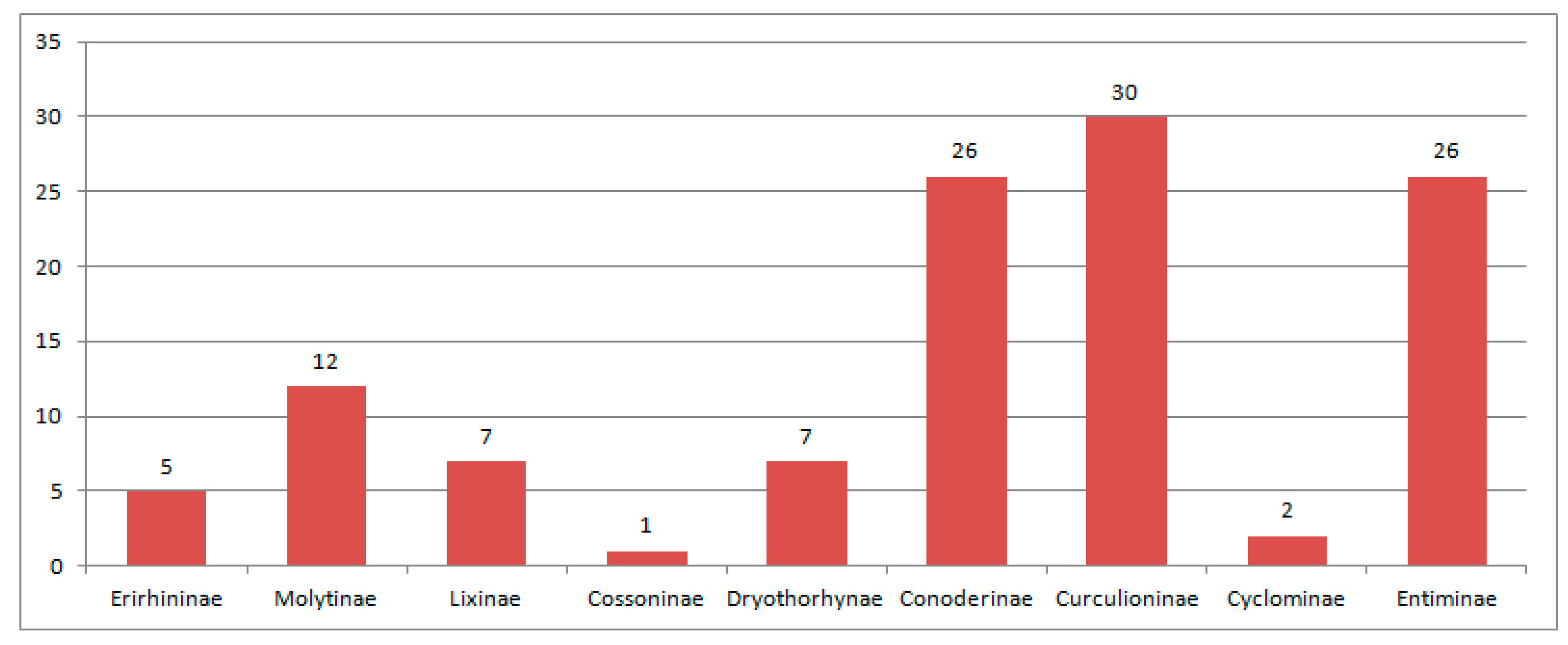
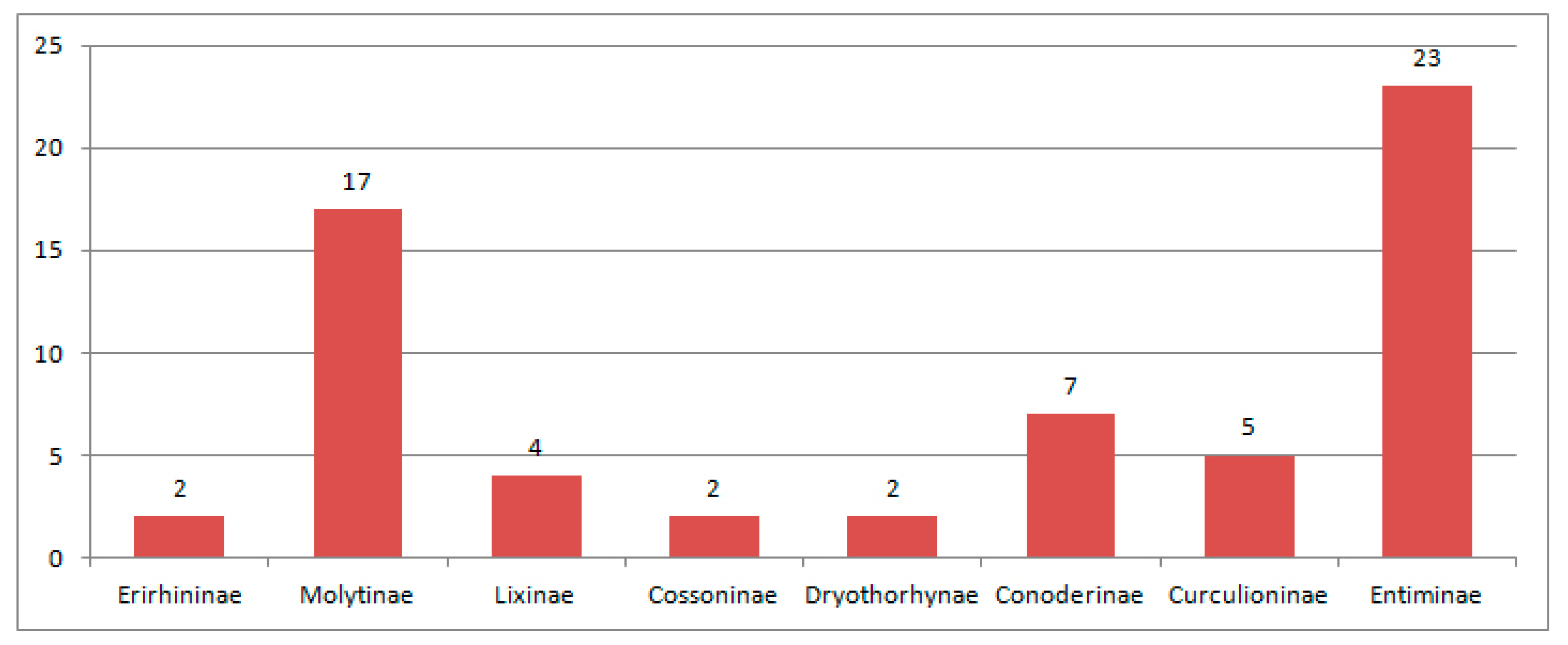
3.2.6. Comparison of the Eocene Weevil Faunas
3.3. Oligocene Weevil Fauna
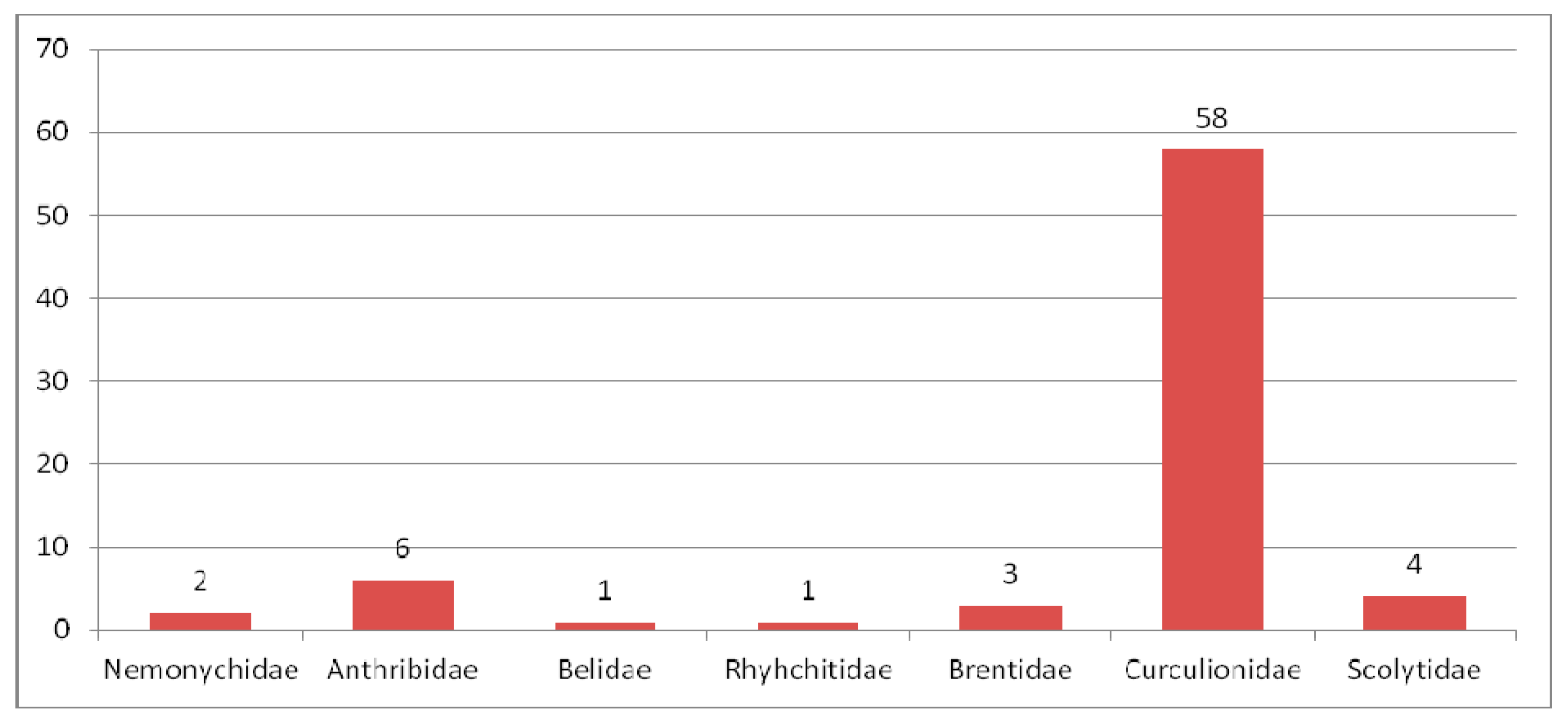
3.3.1. Review of Curculionoidea Families in the Oligocene
Nemonychidae in the Oligocene
Anthribidae in the Oligocene
Rhynchitidae in the Oligocene
Attelabidae in the Oligocene
Brentidae in the Oligocene
Curculionidae in the Oligocene
3.3.2. Early Oligocene Weevil Fauna
3.3.3. Middle Oligocene Weevil Fauna
3.3.4. Late Oligocene Weevil Fauna
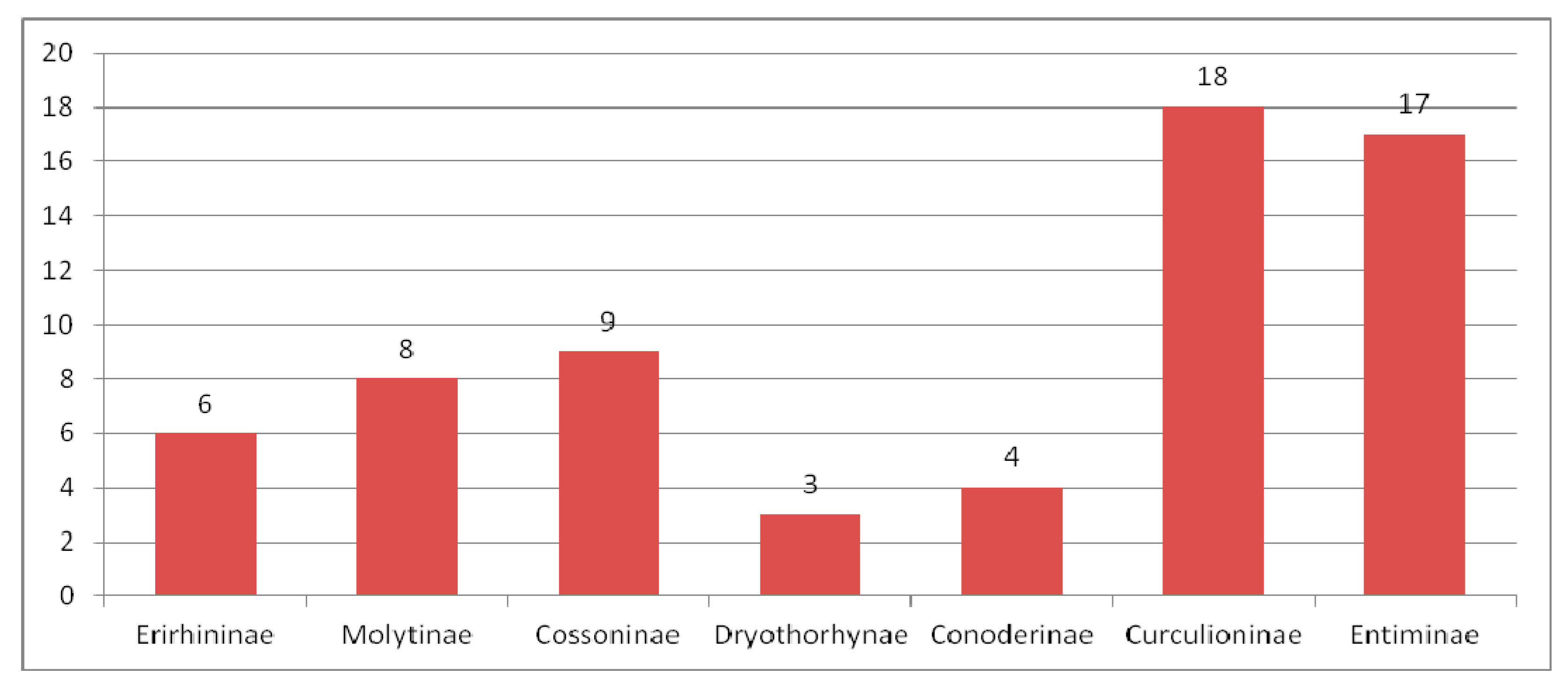
3.3.5. Comparing of the Oligocene Weevil Faunas
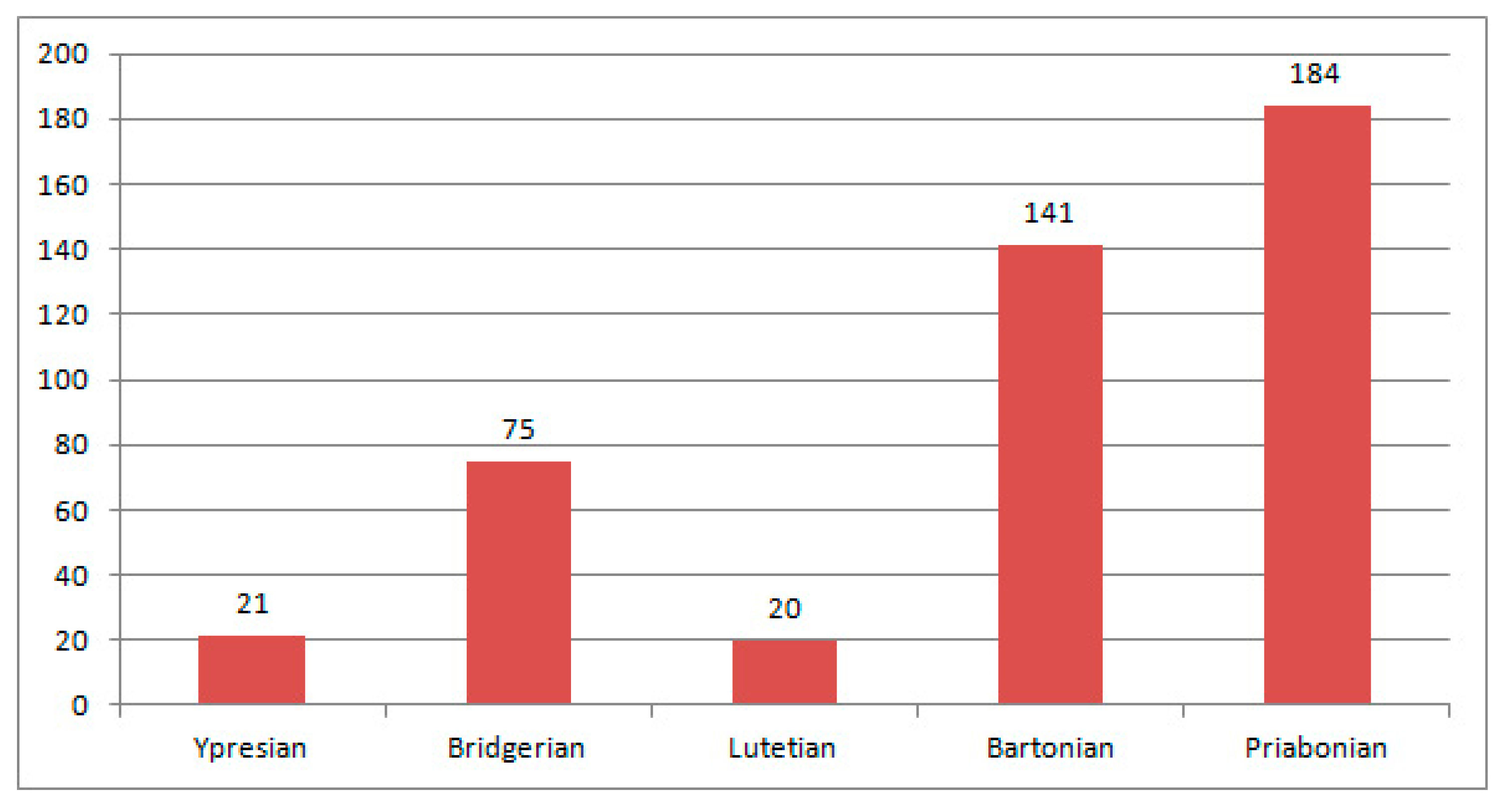
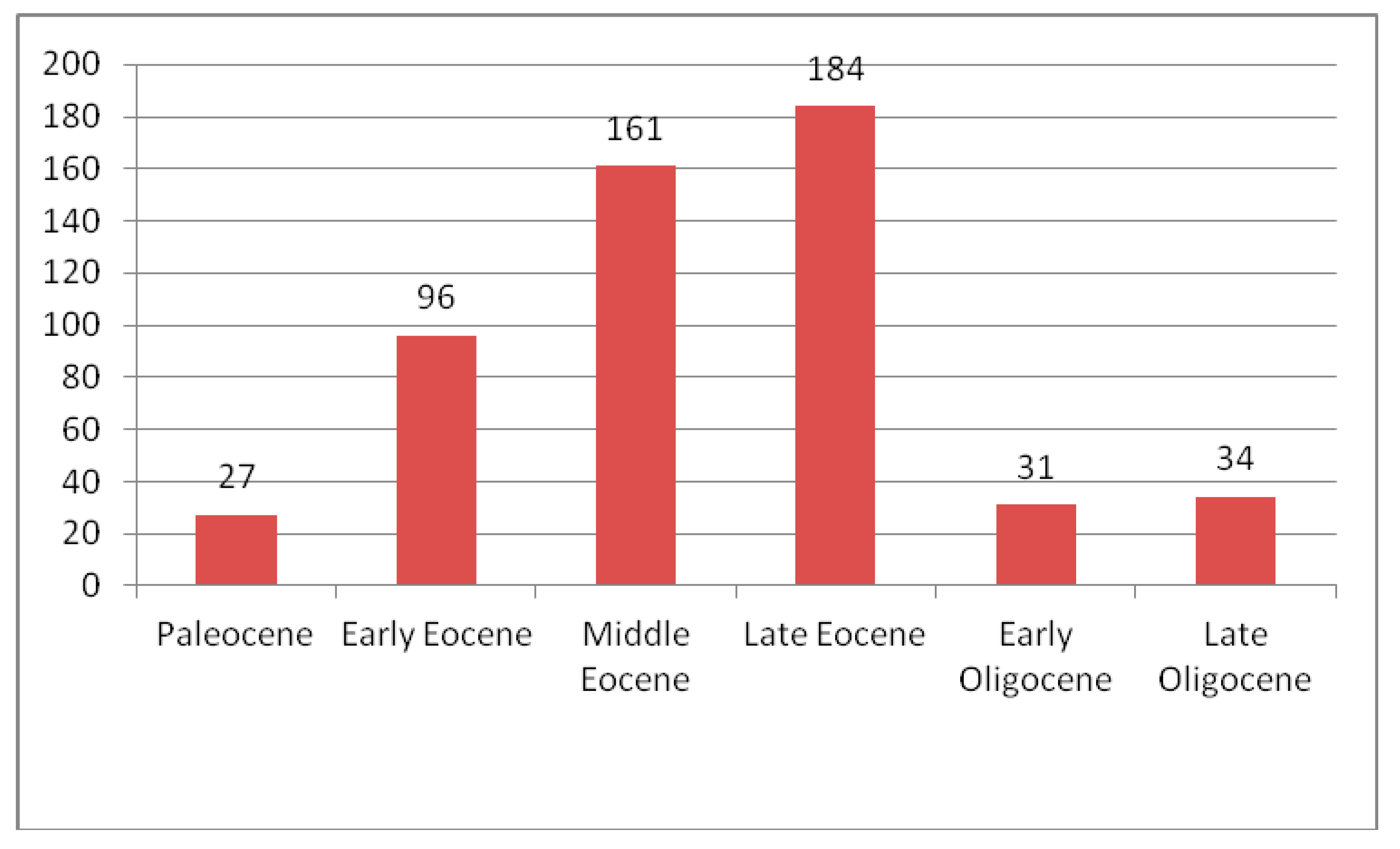
4. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Scotese, C.R. Atlas of Paleogene Paleogeographic Maps (Mollweide Projection), Maps 8–15, Volume 1, The Cenozoic, PALEOMAP Atlas for ArcGIS, PALEOMAP Project, Evanston, IL. 2014. Available online: https://www.researchgate.net/publication/267569784_Atlas_of_Paleogene_Paleogeographic_Maps_Mollweide_Projection_Maps_8-15_Volume_1_The_Cenozoic_PALEOMAP_Atlas_for_ArcGIS_PALEOMAP_Project_Evanston_IL (accessed on 6 September 2020).
- Blythe, A.E.; Kleinspehn, K.L. Tectonically versus climatically driven Cenozoic exhumation of the Eurasian plate margin, Svalbard: Fission track analyses. Tectonics 1998, 17, 621–639. [Google Scholar] [CrossRef]
- Nel, A. The oldest bee fly in the French Paleocene (Diptera: Bombyliidae). C.R.Palevol 2008, 7, 401–405. [Google Scholar] [CrossRef]
- Wappler, T.; Currano, E.D.; Wilf, P.; Rust, J.; Labandeira, C.C. No post-Cretaceous ecosystem depression in European forests? Rich insect feeding damage on diverse middle Palaeocene plants, Menat, France. Proc. R. Soc. B: Biol. Sci. 2009, 276, 4271–4277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyke, G.; Lindow, B. Taphonomy and abundance of birds from the Lower Eocene Fur Formation of Denmark. Geol. J. 2009, 44, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.; Beckett, H.T.; Close, R.A.; Johanson, Z. The English Chalk and London Clay: Two remarkable British bony fish Lagerstätten. Geolog. Soc., London, Special Publ. 2015, 430, 165–200. [Google Scholar] [CrossRef]
- Smith, M.E.; Singer, B.; Carroll, A. 40Ar/39Ar geochronology of the Eocene Green River Formation, Wyoming. GSA Bull. 2003, 115, 549–565. [Google Scholar] [CrossRef] [Green Version]
- Kirejtshuk, A.G.; Legalov, A.A.; Nel, A. A new genus of the subfamily Apioninae (Coleoptera: Brentidae) from the Lower Eocene Oise amber. Paleontol. J. 2015, 49, 1436–1441. [Google Scholar] [CrossRef]
- Wilf, P.; Singer, B.S.; Zamaloa, M.C.; Johnson, K.R.; Cúneo, N.R. Early Eocene 40Ar/39Ar age for the Pampa de Jones plant, frog and insect biota (Huitrera Formation, Neuquén Province, Patagonia, Argentina). Ameghiniana 2010, 47, 207–216. [Google Scholar] [CrossRef]
- Mathewes, R.W.; Greenwood, D.R.; Archibald, S.B. Paleoenvironment of the Quilchena flora, British Columbia, during the Early Eocene Climatic Optimum. Can. J. Earth Sci. 2016, 53, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Mustoe, G.E. Geologic history of Eocene Stonerose Fossil Beds, Republic, Washington, USA. Geosciences 2015, 5, 243–263. [Google Scholar] [CrossRef] [Green Version]
- Lenz, O.K.; Wilde, M.; Mertz, D.F.; Riegel, W. New palynology-based astronomical and revised 40Ar/39Ar ages for the Eocene maar lake of Messel (Germany). Int. J. Earth. Sci. 2015, 104, 873–889. [Google Scholar] [CrossRef]
- West, I.M. The Eocene Cliffs of Bournemouth Dorset. Geology of the Wessex Coast. Available online: www.southampton.ac.uk/~imw/Bournemouth-Geology.htm (accessed on 9 July 2016).
- Ring, S.J.; Bocherens, H.; Wings, O.; Rabi, M. Divergent mammalian body size in a stable Eocene greenhouse climate. Sci. Rep. 2020, 10, 3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachse, M. A remarkable fossiliferous mass flow deposit in the Eocene Eckfeld Maar (Germany)—Sedimentological, taphonomical and palaeoecological considerations. Facies 2005, 51, 173–184. [Google Scholar] [CrossRef]
- Evanoff, E.; McIntosh, W.C.; Murphey, P.C. Stratigraphic summary and 40Ar/39Ar geochronology of the Florissant Formation, Colorado. Denver Mus. Nat. Sci. Proc. 2001, 4, 1–16. [Google Scholar]
- Scott, J.W.; Bowring, S.A. High Precision U/Pb Geochronology of Oligocene Tuffs from the White River Formation, Douglas, Wyoming. J. Vertebr. Paleontol. 2000, 20, 69A. [Google Scholar]
- Walther, H.; Kvaček, Z. Early Oligocene flora of Seifhennersdorf (Saxony). Acta Mus. Nat. Pragae, B, Hist. Nat. 2007, 63, 85–174. [Google Scholar]
- Schindler, T.; Wuttke, M. Geology and limnology of the Enspel Formation (Chattian, Oligocene; Westerwald, Germany). Palaeobiodiversity Palaeoenvironments 2010, 90, 21–27. [Google Scholar] [CrossRef]
- Petrulevičius, J.F.; Wappler, T.; Nel, A.; Rust, J. The diversity of Odonata and their endophytic ovipositions from the Upper Oligocene Fossillagerstätte of Rott (Rhineland, Germany). ZooKeys 2011, 130, 67–89. [Google Scholar] [CrossRef]
- Gaudant, J.; Nel, A.; Nury, D.; Véran, M.; Carnevale, G. The uppermost Oligocene of Aix-en-Provence (Bouches-du-Rhône, Southern France): A Cenozoic brackish subtropical Konservat-Lagerstätte, with fishes, insects and plants. C.R.Palevol 2018, 17, 460–478. [Google Scholar] [CrossRef]
- Seyfullah, L.J.; Beimforde, C.; Dal Corso, J.; Perrichot, V.; Rikkinen, J.; Schmidt, A.R. Production and preservation of resins—Past and present. Biol. Rev. 2018, 93, 1684–1714. [Google Scholar] [CrossRef]
- Bukejs, A.; Alekseev, V.I.; Pollock, D.A. Waidelotinae, a new subfamily of Pyrochroidae (Coleoptera: Tenebrionoidea) fromBaltic amber of the Sambian peninsula and the interpretation of Sambian amber stratigraphy, age and location. Zootaxa 2019, 4664, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.T. Observations on the morphology and classification of weevils (Coleoptera, Curculionoidea) with a key to major groups. J. Nat. Hist. 1992, 26, 835–891. [Google Scholar] [CrossRef]
- Kuschel, G. A phylogenetic classification of Curculionoidea to families and subfamilies. Mem. Entomol. Soc. Wash. 1995, 14, 5–33. [Google Scholar]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A world catalogue of families and genera Curculionoidea (Insecta: Coleoptera) (excepting Scolytidae and Platypodidae); Entomopraxis: Barcelona, Spain, 1999; p. 315. [Google Scholar]
- Oberprieler, R.G.; Marvaldi, A.E.; Anderson, R.S. Weevils, weevils, weevils everywhere. Zootaxa 2007, 1668, 491–520. [Google Scholar] [CrossRef] [Green Version]
- Oberprieler, R.G.; Anderson, R.S.; Marvaldi, A.E. Curculionoidea Latreille, 1802: Introduction, Phylogeny. In Handbook of Zoology. Arthropoda: Insecta; Coleoptera, Beetles. Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2014; pp. 285–300. [Google Scholar]
- Gratshev, V.G.; Legalov, A.A. The Mesozoic stage of evolution of the family Nemonychidae (Coleoptera, Curculionoidea). Paleontol. J. 2014, 48, 851–944. [Google Scholar] [CrossRef]
- Legalov, A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea). Paleontol. J. 2015, 49, 1442–1513. [Google Scholar] [CrossRef]
- Zherichin, V.V.; Gratshev, V.G. Obrieniidae, fam. nov., the oldest Mesozoic weevils (Coleoptera, Curculionoidea). Paleontol. J. 1993, 27, 50–69. [Google Scholar]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Reid, C.A.M.; Schmitt, M.; Slipiński, S.A.; et al. Family group names in Coleoptera (Insecta). Zookeys 2001, 88, 1–972. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A. New Obrieniidae from the Jurassic of Kazakhstan (Coleoptera, Obrienioidea). Paleontol. J. 2012, 46, 73–78. [Google Scholar] [CrossRef]
- Wood, S.L. A reclasification of the genera of Scolytidae (Coleoptera). Great Basin Nat. Mem. 1986, 10, 1–126. [Google Scholar]
- Sanborne, M. Biology of Ithycerus noveboracensis (Forster) (Coleoptera) and weevil phylogeny. Evol. Monogr. 1981, 4, 1–80. [Google Scholar]
- Marvaldi, A.E.; Sequeira, A.S.; O’Brien, C.W.; Farrell, B.D. Molecular and morphological phylogenetics of weevils (Coleoptera: Curculionoidea): Do niche shifts accompany diversification? Syst. biol. 2002, 51, 761–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legalov, A.A. Contribution to the knowledge of the family Nemonychidae (Coleoptera) with descriptions of new taxa. Ukr. J. Ecol. 2017, 7, 64–87. [Google Scholar] [CrossRef]
- Legalov, A.A. Annotated key to weevils of the world. Part 1. Families Nemonychidae, Anthribidae, Belidae, Ithyceridae, Rhynchitidae, Brachyceridae and Brentidae. Ukr. J. Ecol. 2018, 8, 780–831. [Google Scholar] [CrossRef]
- Legalov, A.A. Annotated key to weevils of the world. Part 2. Subfamily Molytinae (Coleoptera, Curculionidae). Ukr. J. Ecol. 2018, 8, 340–350. [Google Scholar]
- Legalov, A.A. Annotated key to weevils of the world. Part 3. Subfamily Conoderinae (Coleoptera, Curculionidae). Ukr. J. Ecol. 2018, 8, 494–503. [Google Scholar]
- Legalov, A.A. Annotated key to weevils of the world. Part 4. Subfamilies Erirhininae, Dryophthorinae and Cossoninae (Curculionidae). Ukr. J. Ecol. 2020, 10, 319–331. [Google Scholar] [CrossRef]
- Legalov, A.A. Annotated key to weevils of the world. Part 5. Subfamily Entiminae (Curculionidae). Ukr. J. Ecol. 2020, 10, 332–346. [Google Scholar] [CrossRef]
- Kuschel, G. The Nearctic Nemonychidae (Coleoptera, Curculionoidea). Entomol. Scand. 1989, 20, 121–171. [Google Scholar] [CrossRef]
- Seunggwan, S.; Clarke, D.J.; Lemmon, A.R.; Lemmon, E.M.; Aitken, A.L.; Haddad, S.; Farrell, B.D.; Marvaldi, A.E.; Oberprieler, R.G.; McKenna, D.D. Phylogenomic data yield new and robust insights into the phylogeny and evolution of weevils. Mol. Biol. Evol. 2018, 35, 823–836. [Google Scholar] [CrossRef]
- Rheinheimer, J. Illustrierter Katalog und Bibliographie der Anthribidae der Welt (Insecta: Coleoptera). Mitt. Entomol. Ver. Stuttg. 2004, 39, 1–288. [Google Scholar]
- Gratshev, V.G.; Legalov, A.A. New Mesozoic Ithyceridae beetles (Coleoptera). Paleontol. J. 2011, 45, 76–81. [Google Scholar] [CrossRef]
- Legalov, A.A. New and little known weevils (Coleoptera: Curculionoidea) from the Paleogene and Neogene. Hist. Biol. 2013, 25, 59–80. [Google Scholar] [CrossRef]
- Legalov, A.A.; Poinar, G., Jr. New tribes of the superfamily Curculionoidea (Coleoptera) in Burmese amber. Hist. Biol. 2015, 27, 558–564. [Google Scholar] [CrossRef]
- Poinar, G., Jr.; Brown, A.E.; Legalov, A.A. A new weevil tribe, Mekorhamphini trib. nov. (Coleoptera, Ithyceridae) with two new genera in Burmese amber. Ukr. J. Ecol. 2016, 6, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A.; Nel, A.; Kirejtshuk, A.G. New and little known weevils (Coleoptera: Curculionoidea) from the Paleocene of Menat (France). C.R. Palevol. 2017, 16, 248–256. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr.; Brown, A.E.; Legalov, A.A. A new weevil, Periosomerus tanyorhynchus gen. et sp. nov. (Coleoptera; Ithyceridae) in mid-Cretaceous Burmese amber. Cret. Res. 2019, 104, 104195. [Google Scholar] [CrossRef]
- Zimmerman, E.C. Australian Weevils (Coleoptera. Curculionoidea). I. Anthribidae to Attelabidae; CSIRO Publishing: Melbourne, Australia, 1994. [Google Scholar]
- Hamilton, R.W. Taxanomic use of endophallic structures in some Attelabidae and Rhynchitidae of America, North of Mexico (Coleoptera: Curculionoidea), with notes on nomenclature. Ann. Entomol. Soc. America. 1979, 72, 29–34. [Google Scholar] [CrossRef]
- Sawada, Y. A systematic study of the family Rhynchitidae of Japan (Coleoptera, Curculionoidea). Hum. Nat. 1993, 2, 1–93. [Google Scholar]
- Legalov, A.A. Taxonomy, classification and phylogeny of the leaf-rolling weevils (Coleoptera: Rhynchitidae, Attelabidae) of the world fauna. Novosib. CD-R 2003. № 0320301200. [Google Scholar]
- Legalov, A.A. Reconstruction of phylogeny in leaf-rolling weevils (Coleoptera, Rhynchitidae, Attelabidae) using the Synap method. Report 1. Zool. Zhurnal. 2004, 83, 1427–1432. [Google Scholar]
- Legalov, A.A. Reconstruction of phylogeny in leaf-rolling weevils (Coleoptera, Rhynchitidae, Attelabidae) using the SYNAP method. Report 2. Zool. Zhurnal. 2005, 84, 190–194. [Google Scholar]
- Legalov, A.A. Trophic relations of leaf-rolling weevils (Coleoptera, Rhynchitidae, Attelabidae). Zool. Zhurnal. 2005, 84, 352–361. [Google Scholar]
- Legalov, A.A. Phylogenetic reconstruction of weevils superfamily Curculionoidea (Coleoptera) using the SYNAP method. Biol. Bull. 2006, 33, 127–134. [Google Scholar] [CrossRef]
- Legalov, A.A. Leaf-Rolling Weevils (Coleoptera: Rhynchitidae, Attelabidae) of the World Fauna; Agro-Siberia: Novosibirsk, Russia, 2007. [Google Scholar]
- Legalov, A.A. The family Rhynchitidae (Insecta: Coleoptera) in the Himalayas. Biodiversitäd und Nat. im Himalaya 2015, 5, 479–486. [Google Scholar]
- Zimmerman, E.C. Australian Weevils (Coleoptera. Curculionoidea). III. Nanophyidae, Rhynchophoridae, Erirhinidae, Curculionidae: Amycterinae, Literature Consulted; CSIRO Publications: Melbourne, Australia, 1993. [Google Scholar]
- Zimmerman, E.C. Australian Weevils (Coleoptera. Curculionoidea). II. Brentidae, Eurhynchidae, Apionidae and a Chapter on Immature Stages by Brenda May; CSIRO Publications: Melbourne, Australia, 1994. [Google Scholar]
- Wanat, M. Genera of Australo-Pacific Rhadinocybinae and Myrmacicelinae, with Biogeography of the Apionidae (Coleoptera: Curculionoidea) and Phylogeny of the Brentidae (s. lato); Mantis: Olsztyn, Poland, 2001. [Google Scholar]
- Zherikhin, V.V.; Egorov, A.B. Weevils (Coleoptera, Curculionidae) from Russian Far East; Institute Biology Soil Science: Vladivostok, Russia, 1991. [Google Scholar]
- Morimoto, K.; Kojima, H. Systematic position of the tribe Phylloplatypodini, with remarks on the definitions of the families Scolytidae, Platypodidae, Dryophthoridae and Curculionidae (Coleoptera: Curculionoidea). Esakia 2004, 44, 153–168. [Google Scholar]
- Legalov, A.A. New and little known Apioninae (Coleoptera, Brentidae) in Eocene Baltic amber. Evraz. Entomol. Zhurn. 2012, 11, 219–222. [Google Scholar]
- Legalov, A.A. A review of the tribe Auletini (Coleoptera, Rhynchitidae) from the Russian fauna. 2. Subtribe Pseudomesauletina. Zool. Zhurnal 2011, 90, 149–155. [Google Scholar] [CrossRef]
- Legalov, A.A. New weevils (Coleoptera, Curculionoidea) from the Eocene of the Green River, United States: Part 1. Paleontol. J. 2018, 52, 294–302. [Google Scholar] [CrossRef]
- Legalov, A.A. New weevils (Coleoptera, Curculionoidea) from the Eocene of the Green River, United States: Part 2. Paleontol. J. 2018, 52, 421–428. [Google Scholar] [CrossRef]
- Legalov, A.A. A review of the Curculionoidea (Coleoptera) from European Eocene ambers. Geosciences 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A. A new tribe of the subfamily Cretonemonychinae (Coleoptera, Nemonychidae) from the Middle Eocene of the Green River. Paleontol. J. 2013, 47, 410–413. [Google Scholar] [CrossRef]
- Scudder, S.H. Fossil Coleoptera from the Rocky mountain tertiaries. Bull. US Geol. Geogr. Surv. Terr. 1876, 2, 77–87. [Google Scholar]
- Scudder, S.H. The fossil insects of the Green River shales. Bull. US Geol. Geogr. Surv. Terr. 1878, 4, 747–776. [Google Scholar]
- Riedel, A. A new tribe, genus and species of Nemonychidae from Baltic amber (Coleoptera: Curculionoidea: Nemonychidae: Cimberidinae). Insect Syst. Evol. 2010, 41, 29–38. [Google Scholar] [CrossRef]
- Wickham, H.F. Fossil Coleoptera from Florissant, with descriptions of several new species. Bull. Am. Mus. Nat. Hist. 1911, 30, 53–69. [Google Scholar]
- Scudder, S.H. Tertiary rhynchophorus Coleoptera of the United States. Monogr. US Geol. Surv. Tert. 1893, 21, 1–206. [Google Scholar] [CrossRef] [Green Version]
- Scudder, S.H. The Tertiary insects of North America. US Geol. Surv. Terr. 1890, 13, 1–734. [Google Scholar]
- Zherikhin, V.V. On weevils (Insecta, Coleoptera) from the Baltic amber. Tr. Paleontol. Inst. Akad. Nauk. SSSR. 1971, 130, 197–209. [Google Scholar]
- Voss, E. Einige Rhynchophoren der Bernsteinfauna (Col.). Mitt. aus dem Geol. Staatsinst. Hambg. 1953, 22, 119–140. [Google Scholar]
- Legalov, A.A.; Kirejtshuk, A.G.; Nel, A. New weevils (Coleoptera, Curculionoidea) from the earlymost Eocene Oise amber. Paleontol. J. 2019, 53, 729–751. [Google Scholar] [CrossRef]
- Legalov, A.A. New Curculionoid beetles (Coleoptera: Curculionoidea) from the Baltic amber. Paleontol. J. 2012, 46, 262–272. [Google Scholar] [CrossRef]
- Gratshev, V.G.; Perkovsky, E.E. New species of the genus Glaesotropis (Insecta: Coleoptera: Anthribidae) from Rovno amber. Paleontol. J. 2008, 42, 60–63. [Google Scholar]
- Gratshev, V.G.; Zherikhin, V.V. A new anthribid genus from the Baltic amber (Insecta: Coleoptera: Anthribidae). Mitt. aus dem Geol. -Paläontologischen Intitut der Univ. Hambg. 1995, 78, 149–157. [Google Scholar]
- Heyden, C.; Heyden, L. Käfer und Polypen aus der Braunkohle des Siebengebirges, XXII–XXIV. Palaeontographica 1866, 15, 131–156. [Google Scholar]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. A new genus of fungus weevils (Coleoptera: Anthribidae) in Rovno amber. Foss. Rec. 2018, 21, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Foerster, B. Die Insekten des ‘Plattigen Steinmergels’ von Brunstatt, XI–XVI. Abh. Geol. Spec. Elsass Lotheringen 1891, 3, 333–594. [Google Scholar]
- Heyden, C.H.G. Gliederthiere aus der Braunkohle des Niederrhein’s, der Wetterau und der Röhn. Palaeontographica 1862, 10, 62–82. [Google Scholar]
- Schlechtendal, D.H.R. Beiträge zur Kenntnis Fossiler Insekten aus dem Braunkohlengebirge von Rott am Sie bengebirge, XII–XIV. Abh. Nat. Gesellsch. Halle 1894, 20, 197–228. [Google Scholar]
- Legalov, A.A. Two new genera and four new species of fossil weevils (Coleoptera: Curculionoidea) in Baltic amber. Entomol. Fenn. 2016, 27, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A.; Poinar, G.O., Jr. Two new species and new findings of Curculionoidea (Insecta: Coleoptera) in Baltic amber. Ukr. J. Ecol. 2020, 10, 357–364. [Google Scholar] [CrossRef]
- Legalov, A.A. A new weevil genus of the tribe Metrioxenini (Coleoptera: Belidae) in Eocene Baltic amber. Hist. Biol. 2012, 24, 213–217. [Google Scholar] [CrossRef]
- Riedel, A.; dos Santos Rolo, T.; Cecilia, A.; van de Kamp, T. Sayrevilleinae Legalov, a newly recognised subfamily of fossil weevils (Coleoptera, Curculionoidea, Attelabidae) and the use of synchrotron microtomography to examine inclusions in amber. Zool. J. Linn. Soc. 2012, 165, 773–794. [Google Scholar] [CrossRef] [Green Version]
- Kania, J.; Legalov, A.A. A new genus of tooth-nosed snout weevils (Coleoptera: Rhynchitidae) in Baltic amber. Paleontol. J. 2019, 53, 1040–1044. [Google Scholar] [CrossRef]
- Bukejs, A.; Legalov, A.A. The first record of Rhynchitidae (Coleoptera) from Rovno amber. Entomol. Fenn. 2019, 30, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.F. A report on some recent of fossil Coleoptera from the Miocene Shales of Florissant, I–VIII. Bull. Lab. Nat. Hist. St. Univ. Iowa 1912, 6, 3–38. [Google Scholar]
- Wickham, H.F. Fossil Coleoptera from the Wilson Ranch near Florissant, Colorado, I–VII. Bull. Lab. Nat. Hist. State. Univ. Iowa. 1913, 6, 3–29. [Google Scholar]
- Zherikhin, V.V. Tertiary weevils (Insecta, Coleoptera: Curculionoidea), identified from the collections of the Senckenberg Museum, Senckenberg. Lethaea 1992, 72, 169–178. [Google Scholar]
- Piton, L. Paléontologie du gisement Éocène de Menat (PuydeDôme) (Flore et faune); Imprim. Paul Vallier: ClermontFerrand, France, 1940. [Google Scholar]
- Wickham, H.F. New fossil Coleoptera from the Florissant beds Bull. Lab. Nat. Hist. St. Univ. Iowa 1916, 7, 3–19, pls. I–IV. [Google Scholar]
- Wickham, H.F. On some fossil Rhynchophorus Coleoptera from Florissant Colorado, I–IV. Bull. Am. Mus. Nat. Hist. 1912, 31, 41–55. [Google Scholar]
- Rheinheimer, J. Neue fossile Rüsselkäfer (Coleoptera: Curculionidae) aus dem Eozän des Baltischen Bernsteins und der Grube Messel bei Darmstadt. Staatliches. Mus. Nat. 2007, 365, 1–24. [Google Scholar]
- Legalov, A.A. The oldest Brentidae and Curculionidae (Coleoptera: Curculionoidea) from the Aptian of Bon-Tsagaan. Hist. Biol. 2014, 26, 6–15. [Google Scholar] [CrossRef]
- Legalov, A.A. Two new weevil genera of the family Brentidae (Coleoptera) in Baltic amber. Entomol. Fennica 2018, 29, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Voss, E. Einige Rüsselkäfer der Tertiärzeit aus baltischen Bernstein (Coleoptera, Curculionidea). Steenstupia 1972, 2, 167–181. [Google Scholar]
- Legalov, A.A.; Bukejs, A. Succinapion telnovi n. gen. et n. sp. of the tribe Kalcapiini (Coleoptera: Brentidae: Apioninae) in Baltic amber. Hist. Biol. 2014, 26, 603–607. [Google Scholar] [CrossRef]
- Wagner, H. Ein neues Apion aus dem baltischen Bernstein (Coleoptera, Curculionidae). Deut. Entomol. Zeitsch. 1924, 1924, 134–136. [Google Scholar]
- Bukejs, A.; Legalov, A.A. The first record of Brentidae (Coleoptera) in Eocene Rovno amber with description of a new fossil species of Toxorhynchus Scudder, 1893. Fossil Record 2020, 23, 169–177. [Google Scholar] [CrossRef]
- Legalov, A.A. New weevils of the family Brentidae (Coleoptera) in Baltic amber. Paleontol. J. (In Press).
- Oustalet, M.E. Recherches sur les insectes fossiles des terraines tertiaires de la France. 2me partie. Insectes fossiles d’Aix en Provence. Ann. Sci. Géol. Paris 1874, 5, 1–347. [Google Scholar]
- Legalov, A.A. First record of the weevil subfamily Nanophyinae (Coleoptera, Brentidae) from the Eocene of the Green River, United States. Paleontol. J. 2015, 49, 399–401. [Google Scholar] [CrossRef]
- Théobald, N. Les insectes fossiles des terrains oligocènes de France. Bull. Mensuel. Soc. Sci. Nancy 1937, 1, 1–473. [Google Scholar]
- Legalov, A.A.; Wappler, T. The oldest record of straight-snouted weevils (Coleoptera: Curculionoidea: Brentidae: Brentinae) from the Eocene of Germany. Hist. Biol. 2019, 1–9. [Google Scholar] [CrossRef]
- Britton, E.B. Beetles from the London clay (Eocene) of Bognor regis, Sussex. Bull. Br. Mus. Nat. Hist. Geol. 1960, 4, 27–50. [Google Scholar]
- Legalov, A.A. New weevils (Curculionidae) in Baltic amber. Paleontol. J. 2016, 50, 970–985. [Google Scholar] [CrossRef]
- Bukejs, A.; Legalov, A.A. A new species of the genus Dorytomus Germar, 1817 (Coleoptera, Curculionidae) from Baltic amber. Entomol. Fenn. 2019, 30, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. New weevils (Coleoptera: Curculionidae) from Rovno amber. Paleontol. J. 2019, 53, 1045–1059. [Google Scholar] [CrossRef]
- Legalov, A.A. Checklist of Mesozoic Curculionoidea (Coleoptera) with description of new taxa. Balt. J. Col. 2010, 10, 71–101. [Google Scholar]
- Oustalet, F.E. Recherches sur les insectes fossiles des terraines tertiaires de la France. Ann. Sci. Géol. Paris 1870, 2, 1–178. [Google Scholar]
- Beier, M. Miozäne und oligozäne Insekten aus Österreich und den unmitulbar angrenzenden Gebieten. Sitzungsberichte Österreichische Akademie der Wissenschaften, Mathe matischenNaturwissensschaftliche Klasse, Abteilung 1: Biologie, Mineralogie, Erdkunde und vewandte Wissenschaften 1952, 161, 129–134. [Google Scholar]
- Hustache, A. Un curculionide de l’ambre de la Baltique. Bull. Mens. Soc. Lin. Lyon. 1942, 11, 108–109. [Google Scholar] [CrossRef]
- Ulke, T. A new genus and species of Curculionidae (Coleoptera) in Baltic amber. Not. Nat. 1947, 19, 1–5. [Google Scholar]
- Cockerell, T.D.A. Some Eocene insects from Colorado and Wyoming. Proc. Unit. Stat. Nat. Mus. 1921, 59, 29–39. [Google Scholar] [CrossRef]
- Haupt, H. Die Käfer (Coleoptera) aus der Eozänen Braunkohle des Geiseltales. Geologica 1950, 6, 1–168. [Google Scholar]
- Bukejs, A.; Alekseev, V.I.; Legalov, A.A. A new Eocene genus of the subtribe Tylodina (Coleoptera: Curculionidae) and notes concerning local differences of Baltic amber in the Kaliningrad Region. Foss. Rec. 2020, 23, 75–81. [Google Scholar] [CrossRef]
- Deichmueller, J.V. Fossile Insecten aus dem Diatomeenschiefir von Kutschlin bei Bilin, Böhmen. Nova Acta Leopold. Verh. Kais. Leopold.Carol. Dtsch. Akad. Nat. 1881, 42, 293–331. [Google Scholar]
- Meunier, F. Sur quelques nouveaux insectes des lignites oligocenès (Aquitanien) de Rott, Siebengebirge (Rhénanie). Konin. Akad. Wetensch. Amst. 1924, 26, 605–612. [Google Scholar]
- Heyden, C.H.G. Fossile Insekten aus der Braunkohle der Sieblos. Palaeontographica 1858, 5, 115–120. [Google Scholar]
- Nazarenko, V.Y.; Perkovsky, E.E. A new genus and species of Dryophthorid weevils (Coleoptera, Dryophthoridae: Stromboscerinae) from the Rovno amber. Paleontol. J. 2009, 43, 1097–1100. [Google Scholar] [CrossRef]
- Piton, L.; Theobald, N. La faune entomologique des gisements miopliocènes du Massif Central. Rev. Sci. Nat. Auvergne 1935, 2, 1–81. [Google Scholar]
- Cockerell, T.D.A. Some tertiary fossil insects. An. Mag. Nat. Hist. 1926, 18, 313–324. [Google Scholar] [CrossRef]
- Kuska, A. Three new species of beetles (Coleoptera: Cantharidae, Anobiidae, Curculionidae) from the Baltic amber. Ann. Up. Sil. Mus. Bytom Entomol. 1992, 3, 107–113. [Google Scholar]
- Nazarenko, V.Y.; Legalov, A.A.; Perkovsky, E.E. A new species of the genus Caulophilus Woll. (Coleoptera: Curculionidae: Cossoninae) from the Rovno amber. Paleontol. J. 2011, 45, 287–290. [Google Scholar] [CrossRef]
- Cockerell, T.D.A. Tertiary insects from Argentina, I. Proc. Unit. Stat. Nat. Mus. 1925, 68, 1–5. [Google Scholar] [CrossRef]
- Meunier, F. Über einige fossile Insekten aus den Braunkohlenschichten (Aquitanien) von Rott (Siebengebirge). Z. Deut. Geol. Gesellsch. 1916, 67, 205–225. [Google Scholar]
- Wickham, H.F. New species of fossil beetles from Floris sant, Colorado. Proc. US Nat. Mus. 1917, 52, 463–472. [Google Scholar] [CrossRef]
- Cockerell, T.D.A. Fossil arthropods in the British Museum. II. An. Mag. Nat. Hist. 1920, 9, 273–279. [Google Scholar] [CrossRef]
- Legalov, A.A.; Bukejs, A. A new genus of the tribe Ceutorhynchini (Coleoptera: Curculionidae) in Baltic amber. Entomol. Fenn. 2018, 29, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Kuska, A. New beetle species (Coleoptera, Cantharidae, Curculionidae) from the Baltic amber. Pr. Muz. Ziemi 1996, 44, 13–23. [Google Scholar]
- Legalov, A.A. A new tribe Palaeoanoplini trib. nov. (Coleoptera: Curculionidae) in Baltic amber. Paleontol. J. (in press).
- Legalov, A.A. A new genus of the tribe Curculionini (Coleoptera: Curculionidae) from Baltic amber. Paleontol. J. 2019, 53, 1036–1039. [Google Scholar] [CrossRef]
- Zherikhin, V.V. A weevil of the tribe Curculionini (Insecta: Coleoptera, Curculionidae) from the Paleogene of Havighorst, Germany. Mitt. Geol.–Paläontol. Inst. Hambg. 1995, 78, 145–148. [Google Scholar]
- Cockerell, T.D.A. New species of North American fossil beetles, cockroaches and tsetse flies. Proc. Unit. Stat. Nat. Mus. 1918, 54, 301–311, pls. 54–55. [Google Scholar] [CrossRef]
- Bukejs, A.; Legalov, A.A. Groehnius, a new genus of Eugnomini (Coleoptera: Curculionidae) from Eocene Baltic amber. Foss. Rec. 2019, 22, 45–49. [Google Scholar] [CrossRef]
- Nazarenko, V.Y.; Perkovsky, E.E. A new species of derelomine weevils (Coleoptera, Curculionidae, Curculioninae: Acalyptini) from the Rovno amber. Paleontol. J. 2016, 50, 991–996. [Google Scholar] [CrossRef]
- Heer, O. Ueber die fossilen Insecten von Aix in der Provence. Vierteljahr. Naturforsch. Gesllschaft. Zürich 1856, 5, 1–40. [Google Scholar]
- Germar, E.F. Ueber einige Insekten aus Tertiärgebildungen. Z. Deut. Geol. Gesellsch. 1849, 1, 52–66. [Google Scholar] [CrossRef]
- Nicolas, M. Insectes fossiles d’Aix (Provence). Descriptions de Quelques Nouvelles Espèces (Collection de M. Matheron). Assoc. Fr. pour l’Avancement Sci. 1891, 2, 425–438. [Google Scholar]
- Giebel, C.G.A. Fauna der Vorwelt mit steter Berücksichtigung der lebenden Thiere. Insecten und Spinnen. Monogr. Dargestellt 1856, 2, 1–511. [Google Scholar]
- Zherikhin, V.V. Oligocene seed beetles and weevils (Coleoptera: Bruchidae, Curculionidae) from Bol’shaya Svetlovodnaja River (northern Primorie). In Kainozoi Dal’nego Vost. (Cenozoic of the Far East); Institute Biology Soil Science: Vladivostok, Russia, 1989; pp. 145–150. [Google Scholar]
- Legalov, A.A.; Bukejs, A. A new species of the genus Archaeocallirhopalus (Coleoptera: Curculionidae) in Baltic amber. Entomol. Fenn. 2015, 26, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Haupt, H. Beitrag zur Kenntnis der Eozänen Arthropodenfauna des Gieseltales. Nova Acta Leopoldiana 1956, 18, 1–90. [Google Scholar]
- Wanat, M.; Borovec, L. New genus of weevil (Coleoptera, Curculionidae) from Baltic amber. Pol. Pismo Entomol. 1986, 56, 243–247. [Google Scholar]
- Yunakov, N.N.; Kirejtshuk, A.G. New genus and species of broad-nosed weevils from Baltic amber and notes on fossils of the subfamily Entiminae (Coleoptera, Curculionidae). ZooKeys 2011, 160, 73–96. [Google Scholar] [CrossRef] [PubMed]
- Bukejs, A.; Legalov, A.A. First record of the tribe Naupactini (Coleoptera: Curculionidae) in Rovno amber. Foss. Rec. 2019, 22, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.F. New Miocene Coleoptera from Florissant. Bull. Mus. Compar. Zool. 1914, 58, 423–494, pls. 1–16. [Google Scholar]
- Germar, E.F. Fauna Insectorum europae. 19. Insectorum Protogaea Specimen Systems Insecta Carbonum Fossilum; Kuemmelii: Halae, Germany, 1837; Available online: http://dl.ub.uni-freiburg.de/diglit/germar1837 (accessed on 6 September 2020).
- Williams, M.Y. The stratigraphy and palaeontology of Hong Kong and the new territories. Trans. Royal. Soc. Can. 1943, 3, 93–116. [Google Scholar]
- Cockerell, T.D.A. The fauna of the Sunchal (or Margas Verdes) Formation, Northern Argentina. Am. Mus. Nov. 1936, 886, 1–9. [Google Scholar]
- Wickham, H.F. Coleoptera from the Lower Eocene (Wilcox) clays. J. Wash. Acad. Sci. 1929, 19, 148–150. [Google Scholar]
- Heer, O. Die miocene Flora and Fauna Spitsbergens, I–XVI. Kongl. Sven. Vetensk.–Akad. Handl. 1870, 8, 1–98. [Google Scholar]
- Cockerell, T.D.A. Fossil arthropods in the British Museum. III. An. Mag. Nat. Hist. 1920, 9, 65–72. [Google Scholar] [CrossRef]
- Cockerell, T.D.A. Some American fossil Insects. Proc. Unit. Stat. Nat. Mus. 1917, 51, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Piton, L.; Rudel, M.A. Sur de nouveaux gisemente d’Insectes fossiles dans l’Oligocene de la Limagne. Bull. Mens. Soc. Linn. Lyon 1936, 5, 78–81. [Google Scholar]
- Cockerell, T.D.A. Eocene fossils from Green River, Wioming. Am. J. Sci. 1909, 28, 447–448. [Google Scholar] [CrossRef]
- Piton, L.; Theobald, N. Les insectes fossiles du PuydeMur (Limagne). Bull. Menseul. Soc. Sci. Nancy 1937, 9, 202–214. [Google Scholar]
- Peris, D.; Solórzano Kraemer, M.M.; Smith, S.M.; Cognato, A.I. Eoplatypus jordali gen.n. et sp.n., the first described Platypodinae (Coleoptera: Curculionidae) from Baltic amber. Arthropod Syst. Phyl. 2017, 75, 85–194. [Google Scholar]
- Schedl, K. Die Borkenkäfer des baltischen Bernsteins. Zent. Ges. Entomol. 1947, 2, 12–45. [Google Scholar]
- Hagedorn, M. Borkenkäfer des baltischen Bersteins. Schr. Phys.-Ökonom. Gesell. König. Prus. 1906, 47, 115–121. [Google Scholar]
- Germar, E.F. Insekten in Bernstein eingeschlossen, beschrieben aus dem academischen Mineralien Cabinet zu Halle. Mag. Entomol. 1813, 1, 11–18. [Google Scholar]
- Petrov, A.V.; Zherikhin, V.V. A new species of bark beetles of the genus Hylesinus (Insecta: Coleoptera: Scolytidae) from the Oligocene of France. Paleontol. J. 2004, 38, 187–189. [Google Scholar]
- Petrov, A.V.; Perkovsky, E.E. New species of bark beetles from the Rovno amber (Insecta: Coleoptera: Scolytidae). Paleontol. J. 2008, 42, 406–408. [Google Scholar] [CrossRef]
- Petrov, A.V.; Perkovsky, E.E. A new genus and species of Scolytinae (Coleoptera: Curculionidae) from the Rovno amber. Paleontol. J. 2018, 52, 164–167. [Google Scholar] [CrossRef]
- Martins-Neto, R.G. Review of some Insecta from Mesozoic and Cenozoic Brazilian deposits with descriptions of new taxa. Acta Geol. Leop. 2001, 24, 115–124. [Google Scholar]
- Motschulsky, V. Voyages. Lettres de M. de Motschulsky a M. Ménétriés, 4. Étud. Entomol. 1857, 5, 21–38. [Google Scholar]
- Lin, Q.B.; Lee, C.M. Early Tertiary insects from Peng Chau, Hong Kong. Palaeontol. Strat. Hong Kong 1997, 1, 185–199. [Google Scholar]
- Legalov, A.A. Fossil history of Mesozoic weevils (Coleoptera: Curculionoidea). Insect Sci. 2012, 19, 683–698. [Google Scholar] [CrossRef]
- Kuschel, G. Reappraisal of the Baltic Amber Curculionoidea described by E. Voss. Mitt. Geol.-Paläontol. Inst. Hambg. 1992, 73, 191–215. [Google Scholar]
- Legalov, A.A. A review of fossil and recent species of the family Ithyceridae (Coleoptera) from the world fauna, I–IV. Amur. Zool. Zh. 2009, 1, 117–131. [Google Scholar]
- Legalov, A.A. A new classification of ecological groups of the leaf-rolling weevils (Coleoptera: Rhynchitidae, Attelabidae). Evraz. Entomol. Zhur. 2004, 3, 43–45. [Google Scholar]
- Caldara, R.; Franz, N.M.; Oberprieler, R.G. Chapter 2. Curculioninae Latreille 1802. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles; Volume 3: Morphology and systematics (Phytophaga); Kristensen, N.P., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2014; Volume 3, pp. 589–628. [Google Scholar]
- Meregalli, M. 3.7.6. Lixinae Schoenherr 1823. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles; Volume 3: Morphology and Systematics (Phytophaga); Kristensen, N.P., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2014; Volume 3, pp. 523–528. [Google Scholar]
- Alonso-Zarazaga, M.A. Revision of the supraspecific taxa in the Palaearctic Apionidae Schoenherr 1823 (Coleoptera, Curculionoidea). 2. Subfamily Apioninae Schoenherr, 1823: Introduction, keys and descriptions. Graellsia 1990, 46, 19–156. [Google Scholar]
- Gruas-Cavagnetto, C. Etude palynologique del’Eocene du Bassin Anglo-Parisien. Mem. Soc. Geol. Fr. 1978, 56, 1–64. [Google Scholar]
- Archibald, S.B.; Mathewes, R.W. Early Eocene insects from Quilchena, British Columbia and their paleoclimatic implications. Can. J. Zool. 2000, 78, 1441–1462. [Google Scholar] [CrossRef]
- Gratshev, V.G.; Zherikhin, V.V. The fossil record of weevils and related beetle families (Coleoptera, Curculionoidea). Acta Zool. Cracov. 2003, 46, 129–138. [Google Scholar]
- Wilf, P.; Donovan, M.P.; Cúneo, N.R.; Gandolfo, M.A. The fossil flip-leaves (Retrophyllum, Podocarpaceae) of southern South America. Am. J. Bot. 2017, 104, 1344–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyal, C.H.C. 3.7.7. Molytinae Schoenherr 1823. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles; Volume 3: Morphology and Systematics (Phytophaga); Kristensen, N.P., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2014; Volume 3, pp. 529–569. [Google Scholar]
- Klebs, R. Über Bernsteinschlüsse im allgemein und die Coleopteren meiner Bernsteinsammlung. Schr. Phys.-ökonom. Gesellsch. Königsberg in Pr. 1910, 51, 217–242. [Google Scholar]
- Kirejtshuk, A.G.; Nel, A. Current knowledge of Coleoptera (Insecta) from the lowermost Eocene Oise amber. Insect Syst. Evol. 2013, 44, 175–201. [Google Scholar] [CrossRef]
- Alekseev, V.I. Coleoptera from the middle-upper Eocene European ambers: Generic composition, zoogeography and climatic implications. Zootaxa 2017, 4290, 401–443. [Google Scholar] [CrossRef]
- O’Brien, C.W.; Tang, W. Revision of the New World cycad weevils of the subtribe Allocorynina, with description of two new genera and three new subgenera (Coleoptera: Belidae: Oxycoryninae). Zootaxa 2015, 3970, 1–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poinar, G.J.; Legalov, A.A. Pleurambus strongylus n. gen., n. sp. (Coleoptera: Belidae) in Dominican amber. Hist. Biol. 2014, 26, 670–674. [Google Scholar] [CrossRef]
- Hassan, M.Y. Tertiary faunas from Kap Brewster, East Greenland. Medd. om Grønland 1953, 111, 1–42. [Google Scholar]
- Lutz, H. Beitrag zur Kenntnis der unteroligozänen Insektenfauna von Cereste (Süd-Frankreich). Doc. Nat. 1984, 21, 1–26. [Google Scholar]
- Prokop, J.; Fikacek, M. Early Oligocene insect fauna from Seifhennersdorf (Saxony, Germany). Acta Mus. Nat. Pragae 2007, 63, 205–213. [Google Scholar]
- Cifuentes-Ruiz, P.; Vega, F.J.; Cevallos Ferriz, S.R.S.; Gonzalez-Soriano, E.; Zaragoza Caballero, S.; Garibayromero, L. Oligocene Scorpion and insects (Plecoptera and Coleoptera) from the Los Ahuehuetes Locality, Puebla, Mexico. Ameghiniana 2007, 44, 673–679. [Google Scholar]
- Prokop, J. Remarks on palaeoenviromental changes based on reviewed Tertiary insect associations from the Krušnéhory piedmont basins and the Èeské støedohoøí Mts in northwestern Bohemia (Czech Republic). Acta Zool. Cracov. 2003, 46, 329–344. [Google Scholar]
- Wedmann, S.; Poschmann, M.; Hörnschemeyer, T. Fossil insects from the Late Oligocene Enspel Lagerstätte and their palaeobiogeographic and palaeoclimatic significance. Palaeobio. Palaeoenv. 2010, 90, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Kirejtshuk, A.G.; Ponomarenko, A.G.; Kurochkin, A.S.; Alexeev, A.V.; Gratshev, V.G.; Solodovnikov, A.V.; Krell, F.-T.; Soriano, C. The beetle (Coleoptera) fauna of the Insect Limestone (late Eocene), Isle of Wight, southern England. Earth Env. Sci. Trans. R. Soc. Edinb. 2019, 110, 405–492. [Google Scholar] [CrossRef]



























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
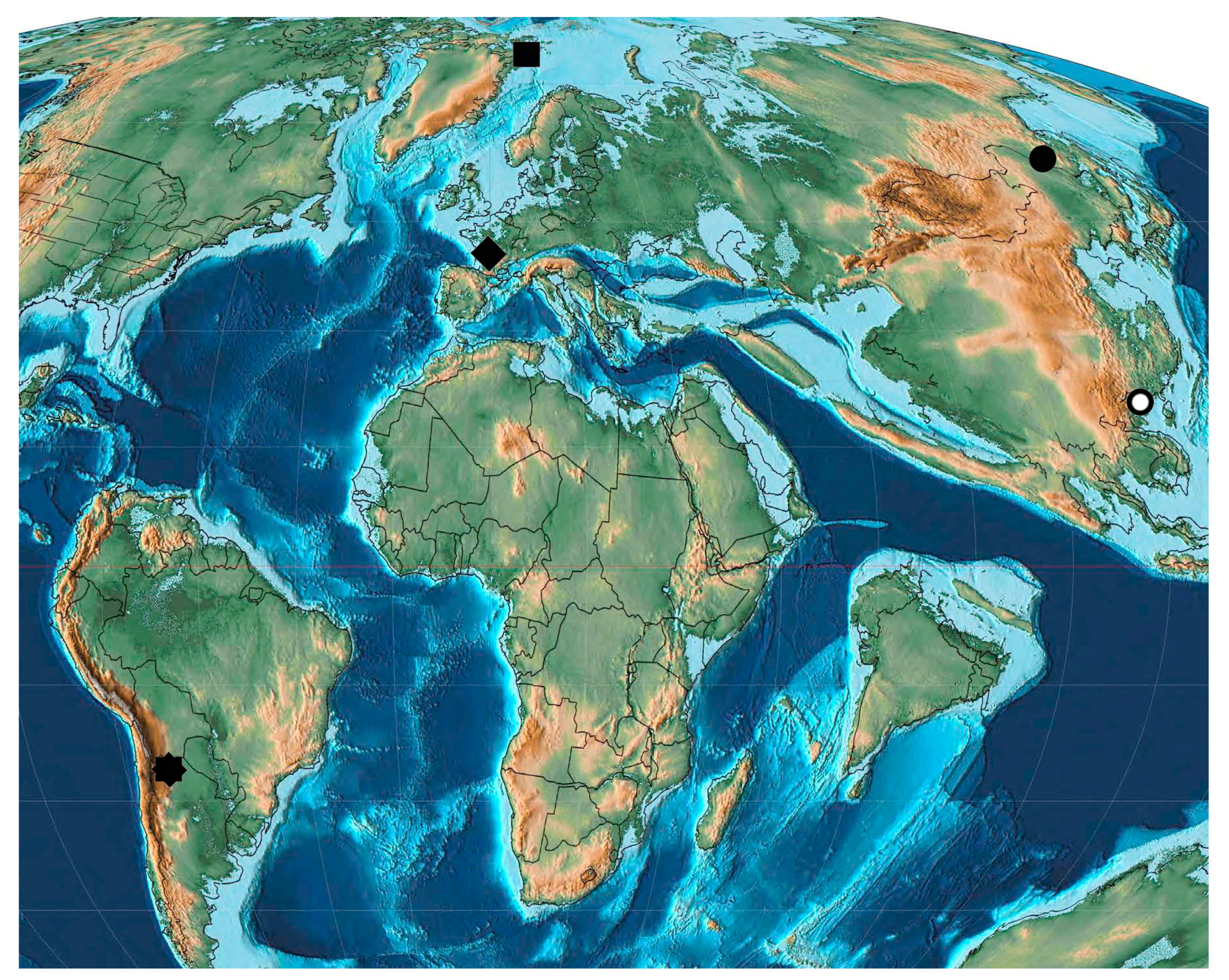
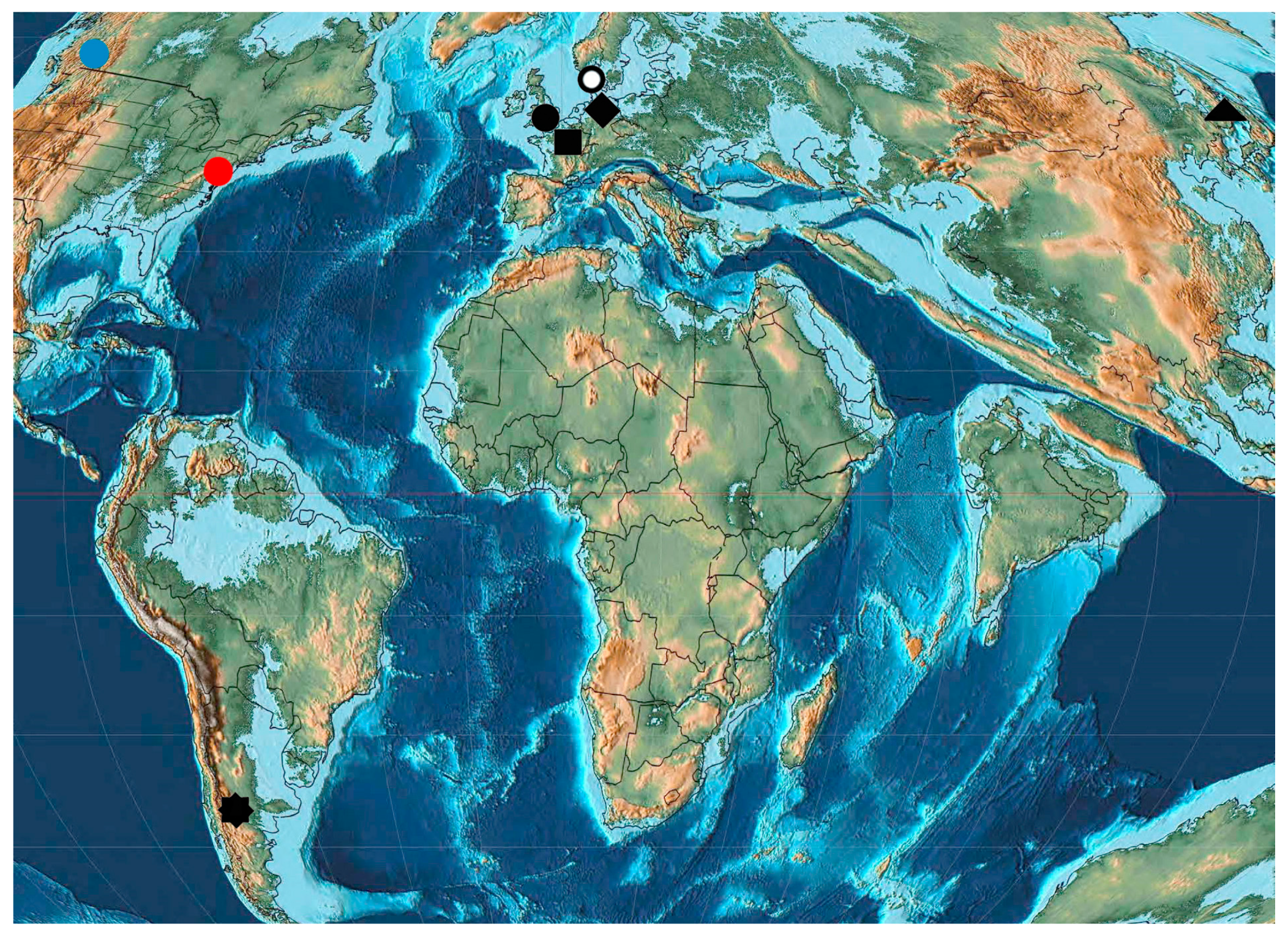
| Name | Abbreviation | Location | Age |
|---|---|---|---|
| Arkhara | Arkh (P1d) | Russia: Amurskaya Region, Arkharinskii District, quarry at Arkhara Railway Station | Lower Paleocene, Danian, Cagayan |
| Starostin | Star (P1d) | Norway: Spitsbergen, Svalbard and Jan Mayen, Firkanten Formation | Lower Paleocene, Danian, 63 ± 2 Ma |
| Sunchal | Sunc (P1d) | Argentina: northern Argentina, Jujuy Province, La Mendieta, Maiz Gordo Formation | Lower Paleocene, Danian, 66.0−55.8 Ma |
| Menat | Mena (P1sl-t) | France: Puy-de-Dome | Middle-Upper Paleocene, Selandian-Thanetian, 61.0–59.0 Ma |
| Mirs Bay | Mirs (P1) | China: Hong Kong, Peng Chau Island, gray sandy shales, Ping Chau Formation | Paleocene |
| Mors | Mors (P2i) | Denmark, Fur Formation | Lower Eocene, Ypresian, 54.0 Ma |
| London Clay | LonC (P2i) | United Kingdom: England: Sussex, Bognor Regis | Lower Eocene, Ypresian, 54.0–50.0 Ma |
| Roan Mountain | RoaM (P2i-l) | United States: Colorado, Green River Formation | Lower-Middle Eocene, Bridgerian, 53.5–48.5 Ma |
| Green River | GreR (P2i-l) | United States: Colorado, Wyoming, Utah States, 3–4 km west of railway crossing of Green River, Green-River Formation | Lower-Middle Eocene, Bridgerian, 53.5–48.5 Ma |
| Oise amber | OisJ(P2i) | France: Paris basin, Creil, Oise | Lowermost Eocene, Ypresian, 53.0 Ma |
| Huitrera Formation | Argentina: Arroyo Chacay, Río Negro, near Estancia Don Hipólito, W, about 60 km east from San Carlos de Bariloche, Huitrera Formation | Lower Eocene, Ypresian, 54.24 ± 0.45 Ma | |
| Peckham | Peck (P2i) | England: South London, Reding Beds | Lower Eocene, Ypresian |
| Havighorst | Havi (P2i) | Germany: east of Hamburg, Schleswig Holstein | Lower Eocene, Ypresian |
| Quilchena | Canada: British Columbia, 3 km south of Nicola Lake, lacustrine shale | Lower Eocene, Ypresian, 51.5 ± 0.4 Ma | |
| Republic | Repu (P2i) | United States: Northeast Washington State, Klondike Mountain Formation | Lower Eocene, Ypresian, 49.0–48.0Ma |
| Tadushi | Tadu (P2i) | Russia: Russian Far East, Primorsky Krai, Kavalerovsky District, Pestrushka River near mouth and Ugol’nyi Creek, tributary of Zerkal’nyi (Tadushi) River, near village of Suvorovo, Tadushi Formation | Lower Eocene, Ypresian |
| Messel | Mess (P2l) | Germany: near Frankfurt, oil shales | Middle Eocene, Upper Ypresian Lower Lutetian, 48.27 ± 0.22–47.0 Ma |
| Corfe | Corf (P2l) | United Kingdom: England: Dorset, Corfe Clay, Lower Bagshot Beds | Middle Eocene, Lutetian, |
| Bournemouth | Bour (P2l) | United Kingdom: England, Dorset, Bagshot Series | Middle Eocene, Lutetian, 50.0–42.0 Ma |
| Geiseltal | Geis (P2l) | Germany: near Halle | Middle Eocene, Lutetian, 47.5–42.5 Ma |
| Eckfelder Maar | Maar (P2l) | Germany: Rheinland-Pfalz, Rhine Palatine | Middle Eocene, Lutetian, 44.3 Ma |
| Baltic amber (Kaliningrad region) | BalJ(P2b) | Russia: Kaliningrad Region, Baltic Sea coast and Amber Jantarnyi quarry near Kaliningrad, Prussian Formation | Middle Eocene, Bartonian, 48.0–33.0 Ma |
| Polish amber | PolJ(P2b) | Poland: Gdansk city area, at the Wisla River Estuary, Baltic amber, Prussian Formation; | Middle Eocene, Bartonian, 48.0–33.0 Ma |
| Scandinavian amber | ScanJ(P2b) | Denmark, amber deposits on the Danish coast, Baltic amber, Prussian Formation | Middle Eocene, Bartonian, 48.0–33.0 Ma |
| Rovno amber | RovJ (P2b) | Ukraine: Rovno Region, Klesov and Dubrovice quarries | Middle Eocene, Bartonian, 48.0–33.0 Ma |
| Bitterfeld amber | Germany: Saxony-Anhalt, Goitzsche near Bitterfeld | Middle Eocene, Bartonian, 48.0–33.0 Ma | |
| Romanian amber | Romania: Alunisului Hill, Valea Sibiciului, Colti | Middle Eocene, Bartonian, 48.0–33.0 Ma | |
| Kutschlin | Kuts (P2b) | Czech Republic, northwest Bohemia, near Bilina | Middle Eocene, Bartonian |
| Celas | Cela (P2p) | France: Gard Department, railway Uze’s–Saint-Julien-de-Casig-nac, Fumades, Corents, Bassein Ales | Upper Eocene, Priabonian |
| Ales-Monteils | Ales (P2p) | France: Gard | Upper Eocene, Priabonian |
| Florissant | Flor (P2p) | United States: Colorado, Rocky Mountains near Pike’s Peak, Florissant Formation | Latest Eocene, Priabonian, 34.07 ± 0.10 Ma |
| Biamo | Biam (P2p) | Russia: Russian Far East, Primorsky Region, Pozharskii District, source of Barachek Creek, right tributary of Bol’shaya Svetlovodnaya (formerly Biamo) River | Upper Eocene, Priabonian |
| Fonseca | Fons (P2p) | Brazil: Minas Gerais State, near Fonseca municipality, Whitish shales, under bituminous shales, Fonseca Basin, Fonseca Formation | Oligocene, Priabonian |
| White River Badlands | WhiR (P3r) | United States: South Dakota | Lower Oligocene, Rupelian, 34.0–31.2 Ma |
| Brunnstatt | Brun (P3r) | France: Alsace, Haut-Rhine Dept., 5 km southwest of Mulhouse | Lower Oligocene, Rupelian, 33.9 Ma |
| Corent | Core (P3r) | France: South France, Puy-de-Dom Department, Gergovia Plateau, south of Clermon-Ferran | Lower Oligocene, Rupelian |
| Cereste | France: Alpes-de-Haute Province, Basses Alp Department | Lower Oligocene, Rupelian | |
| Seifhennersdorf | Germany: Saxony, Oberlausitz | Lower Oligocene, Rupelian, 30.5–30.2 Ma | |
| Ahuehuetes | Mexico: Puebla, 4.5 km NNE from Tepexi de Rodrigues, Los Ahuehuetes, Coatzingo Formation | Lower Oligocene, Rupelian | |
| Kundratice | Czech Republic: near Litomioice and Seifhennersdorf | Lower Oligocene, Rupelian | |
| Sieblos | Sieb (P3) | Germany: Hessen, Rhon | Middle Oligocene |
| Kleinkembs | Klei (P3) | Germany: Baden-Wurtemberg, 10 km northwest of Lorrach, Pays de Bade, Salt Formation | Middle Oligocene |
| Gaube | Gaub (P3) | France: Puy-de-Dome, ravin de la Gaube | Lower Oligocene, Rupelian |
| Puy-Saint-Jean | PuStJ (P3) | France: Puy-de-St. Jean | Middle Oligocene |
| Luzice | Luzi (P3h) | Czech Republic: northwest Bohemia | Upper Oligocene, Chattian |
| Enspel | Germany: Rheinland-Pfalz, Westerwald, Bad Marienberg | Upper Oligocene, Chattian, 24.79 − 24.56 Ma | |
| Ashutas | Kazakhstan: East Kazakhstan Region, Kurshim District, right bank of River Cherny Irtysh | Upper Oligocene, Chattian | |
| Perekishkyul’ | Azerbaijan: middle course of Sumgait River, village of Perekeshkul, Maikop Formation | Upper Oligocene, Chattian | |
| Rott | Rott (P3h) | Germany: North Rhine-Westfalia, Siebengebirge, near Bonn, Rott Formation | Latest Oligocene, Upper Chattian, 24.0–23.0 Ma |
| Aix-en-Provance | Aix (P3h) | France: Bouches-du-Rhane, Aix-en-Provence Formation | Latest Oligocene, Upper Chattian |
| Kap Dalton | KapD (P3) | Denmark: Greenland | Oligocene |
| Kenderlyk II | Kazakhstan: East Kazakhstan Region, Zaisan district, left bank of Kenderlyk River, 6 km from village of Kenderlyk | Oligocene |
| No. | Taxon | Locality | Age | Sources |
|---|---|---|---|---|
| Nemonychidae Bedel, 1882 | ||||
| Cretonemonychinae Gratshev et Legalov, 2009 | ||||
| Eocaenonemonychini Legalov, 2013 | ||||
| 1 | Eocaenonemonyx kuscheli Legalov, 2013 | GreR | P2i-l | 72 |
| 2 | “Sitones” grandaevus Scudder, 1876 | GreR | P2i-l | 73 |
| 3 | “Eugnamptus” decemsatus Scudder, 1878 | KapD | P3 | 74 |
| Cimberidinae Gozis, 1882 | ||||
| Kuschelomacerini Riedel, 2010 | ||||
| 4 | Kuschelomacer kerneggeri Riedel, 2010 | BalJ | P2b | 75 |
| Anthribidae Billberg, 1820 | ||||
| Anthribinae Billberg, 1820 | ||||
| Cratoparini LeConte, 1876 | ||||
| 5 | “Euparius” elusus (Scudder, 1878) | GreR | P2i-l | 74 |
| 6 | “E.” repertus (Scudder, 1878) | Flor | P2p | 74 |
| 7 | “E.” adumbratus (Wickham, 1911) | Flor | P2p | 76 |
| 8 | ”E.” arcessitus (Scudder, 1893) | Flor | P2p | 77 |
| Anthribini Billberg, 1820 | ||||
| 9 | “Ormiscus“ partitus (Scudder, 1890) | GreR | P2i-l | 78 |
| 10 | “Trigonorhinus“ pristinus (Scudder, 1876) | GreR | P2i-l | 73 |
| 11 | “T.“ sordidus (Scudder, 1893) | Flor | P2p | 77 |
| Ecelonerini Lacordaire, 1865 | ||||
| 12 | Pseudochirotenon eocaenicus Legalov, 2018 | GreR | P2i-l | 69 |
| 13 | Saperdirhynchus prisctitillator Scudder, 1893 | Flor | P2p | 77 |
| Mecocerini Lacordaire, 1865 | ||||
| 14 | Pseudoecocerus aleksevi Legalov, 2020 | BalJ | P2b | 71 |
| Tropiderini Lacordaire, 1865 | ||||
| 15 | “Tropideres“ remotus Scudder, 1893 | GreR | P2i-l | 77 |
| 16 | “T.“ vastats Scudder, 1893 | Flor | P2p | 77 |
| Allandrini Pierce, 1930 | ||||
| 17 | Pseudomecorhis orlovi Zherikhin, 1971 | BalJ | P2b | 79 |
| 18 | P. simulator Voss, 1953 | BalJ | P2b | 80 |
| 19 | Allandroides vossi Legalov, 2015 | BalJ | P2b | 30 |
| Oiserhinini Legalov, Kirejtshuk et Nel, 2019 | ||||
| 20 | Oiserhinus insolitus Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| Zygaenodini Lacordaire, 1865 | ||||
| 21 | Glaesotropis (Pseudoglaesotropis) balticus Legalov, 2020 | BalJ | P2b | 71 |
| 22 | G. (P.) martynovi (Legalov, 2012) | BalJ | P2b | 82 |
| 23 | G. (Glaesotropis) diadiasashai Gratshev et Perkovsky, 2008 | RovJ | P2b | 83 |
| 24 | G. (G.) gusakovi Legalov, 2015 | BalJ | P2b | 30 |
| 25 | G. (G.) minor Gratshev et Zherikhin, 1995 | BalJ | P2b | 84 |
| 26 | G. (G.) succiniferus Legalov, 2015 | BalJ | P2b | 30 |
| 27 | G. (G.) weitschati Gratshev et Zherikhin, 1995 | BalJ | P2b | 84 |
| 28 | G. (Electranthribus) alleni Legalov, 2015 | BalJ | P2b | 30 |
| 29 | G. (E.) gratshevi Legalov, 2015 | BalJ | P2b | 30 |
| 30 | G. (E.) zherikhini (Legalov, 2013) | BalJ | P2b | 47 |
| Tribe incertae sedis | ||||
| 31 | Stiraderes conradi Scudder, 1893 | Flor | P2p | 77 |
| Choraginae W. Kirby, 1819 | ||||
| Choragini W. Kirby, 1819 | ||||
| 32 | “Choragus“ fictilis Scudder, 1890 | GreR | P2i-l | 78 |
| 33 | “Ch.“ tertiarius Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Valenfriesiini Alonso-Zarazaga et Lyal, 1999 | ||||
| 34 | Eduardoxenus unicus Legalov, Nazarenko et Perkovsky, 2018 | RovJ | P2b | 86 |
| Urodontinae C.G Thomson, 1859 | ||||
| 35 | “Bruchela“ cincta (Foerster, 1891) | Brun | P3r | 87 |
| 36 | “B.“ priscus (C. Heyden, 1862) | Rott | P3h | 88 |
| 37 | “B.“ multipunctata (Schlechtendal, 1894) | Rott | P3h | 89 |
| Ithyceridae Schoenherr, 1823 | ||||
| Chilecarinae Legalov, 2009 | ||||
| Chilecarini Legalov, 2009 | ||||
| 38 | Petropsis rostratus Legalov, Kirejtshuk et Nel, 2017 | Mena | P1sl-t | 50 |
| Ithycerinae Schoenherr, 1823 | ||||
| 39 | Eoceneithycerus carpenteri Legalov, 2013 | Repu | P2i | 47 |
| 40 | Ithyceroides klondikensis Legalov, 2015 | Repu | P2i | 30 |
| Belidae Schoenherr, 1826 | ||||
| Oxycoryninae Schoenherr, 1840 | ||||
| Oxycraspedini Marvaldi et Oberprieler, 2006 | ||||
| 41 | Oxycraspedus poinari Legalov, 2016 | BalJ | P2b | 90 |
| Metrioxenini Voss, 1953 | ||||
| Metrioxenina Voss, 1953 | ||||
| 42 | Archimetrioxena electrica Voss, 1953 | BalJ | P2b | 80 |
| 43 | A. zherikhini (Legalov, 2012) | BalJ | P2b | 82 |
| Zherichinixenina Legalov, 2009 | ||||
| 44 | Paltorhynchus bisculcatus Scudder, 1893 | RoaM | P2i-l | 77 |
| 45 | P. narwhal Scudder, 1893 | Flor | P2p | 77 |
| 46 | Succinometrioxena attenuata Legalov et Poinar, 2020 | BalJ | P2b | 91 |
| 47 | S. bachofeni Legalov, 2013 | BalJ | P2b | 47 |
| 48 | S. poinari Legalov, 2012 | BalJ | P2b | 92 |
| Allocorynitae Sharp, 1890 | ||||
| Palaeorhopalotriini Legalov, 2013 | ||||
| 49 | Palaeorhopalotria neli Legalov, 2013 | Ales | P2p | 47 |
| Rhynchitidae Gistel, 1856 | ||||
| Sayrevilleinae Legalov, 2003 | ||||
| Sanyrevilleini Legalov, 2003 | ||||
| 50 | Baltocar convexus Legalov, 2015 | BalJ | P2b | 30 |
| 51 | B. groehni Riedel, 2012 | BalJ | P2b | 93 |
| 52 | B. hoffeinsorum Riedel, 2012 | BalJ | P2b | 93 |
| 53 | B. subnudus Riedel, 2012 | BalJ | P2b | 93 |
| 54 | B. succinicus (Voss, 1953) | BalJ | P2b | 80 |
| Vossicartini Legalov, 2003 | ||||
| 55 | Germanocartus heydeni (Schlechtendal, 1894) | Rott | P3h | 89 |
| Rhynchitinae Gistel, 1856 | ||||
| Auletini Desbrochers des Loges, 1908 | ||||
| Auletina Desbrochers des Loges, 1908 | ||||
| 56 | Electrauletes unicus Legalov, 2015 | BalJ | P2b | 30 |
| Pseudauletina Voss, 1933 | ||||
| 57 | Eoropseudauletes plucinskii Kania et Legalov, 2019 | BalJ | P2b | 94 |
| Pseudomesauletina Legalov, 2003 | ||||
| 58 | Pseudomesauletes groehni Bukejs et Legalov, 2019 | RovJ | P2b | 95 |
| 59 | P. culex (Scudder, 1893) | Flor | P2p | 77 |
| 60 | P. ibis (Wickham, 1912) | Flor | P2p | 96 |
| 61 | P. obliquus (Wickham, 1913) | Flor | P2p | 97 |
| 62 | P. striaticeps (Wickham, 1911) | Flor | P2p | 76 |
| Subtribe incertae sedis | ||||
| 63 | “Teretrum” quiescitum Scudder, 1893 | GreR | P2i-l | 77 |
| 64 | T. primulum Scudder, 1893 | Flor | P2p | 77 |
| 65 | Docirhynchus terebrans Scudder, 1893 | Flor | P2p | 77 |
| 66 | “Trypanorhynchus” depratus Scudder, 1893 | Flor | P2p | 77 |
| 67 | “Paltorhynchus” rectirostris Scudder, 1893 | Flor | P2p | 77 |
| 68 | “Trypanorhynchus” sedatus Scudder, 1893 | Flor | P2p | 77 |
| Rhynchitini Gistel, 1856 | ||||
| Temnocerina Legalov, 2003 | ||||
| 69 | Eocenorhynchites vossi Legalov, 2012 | BalJ | P2b | 74 |
| Perrhynchitina Legalov, 2003 | ||||
| 70 | Succinorhynchites alberti Legalov, 2013 | BalJ | P2b | 47 |
| 71 | Tatianaerhynchites goergesi (Zherikhin, 1992) | Rott | P3h | 98 |
| Rhynchitina Gistel, 1856 | ||||
| 72 | Cartorhynchites struvei Zherikhin, 1992 | Rott | P3h | 98 |
| 73 | Opacoinvolvulus rottensis (Zherikhin, 1992) | Rott | P3h | 98 |
| 74 | O. zherichini Legalov, 2003 | Rott | P3h | 55 |
| Subtribe incertae sedis | ||||
| 75 | “Rhysosternum” punctatolineatum Piton, 1940 | Mena | P1sl-t | 99 |
| 76 | Isothea alleni Scudder, 1893 | Flor | P2p | 77 |
| =Trypanorhynchus corruptivus Scudder, 1893 | 77 | |||
| 77 | Prodeporaus curiosum (Scudder, 1893) | Flor | P2p | 77 |
| 78 | P. exanimale (Scudder, 1893) | Flor | P2p | 77 |
| 79 | P. exilis (Wickham, 1913) | Flor | P2p | 97 |
| 80 | P. minutissimus (Wickham, 1913) | Flor | P2p | 97 |
| 81 | P. smithii (Scudder, 1893) | Flor | P2p | 77 |
| 82 | “Prodeporaides“ laminarum (Wickham, 1916) | Flor | P2p | 100 |
| 83 | “P.“ subterraneus (Scudder, 1893) | Flor | P2p | 77 |
| 84 | “P.“ vulcan (Wickham, 1916) | Flor | P2p | 100 |
| 85 | P. wymani (Scudder, 1893) | Flor | P2p | 77 |
| 86 | “Masteutes” saxifer Scudder, 1893 | Flor | P2p | 77 |
| 87 | “Rhynchites” hageni Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Eugnamptini Voss, 1930 | ||||
| 88 | “Eugnamptidea” florissantensis Wickham, 1913 | Flor | P2p | 97 |
| 89 | E. robusta Wickham, 1916 | Flor | P2p | 100 |
| 90 | E. tertiaria Wickham, 1912 | Flor | P2p | 101 |
| Attelabidae Billberg, 1820 | ||||
| Attelabinae Billberg, 1820 | ||||
| Attelabitae Billberg, 1820 | ||||
| Euscelini Voss, 1925 | ||||
| Clinolabina Legalov, 2003 | ||||
| 91 | Paleoclinolabus dormitus (Scudder, 1893) | Flor | P2p | 77 |
| Attelabini Billberg, 1820 | ||||
| 92 | “Phytonomus” punctatus Piton, 1940 | Mena | P1sl-t | 99 |
| 93 | Palaeoalatorostrum schaali Rheinheimer, 2007 | Mess | P2l | 102 |
| Brentidae Billberg, 1820 | ||||
| Apioninae Schoenherr, 1823 | ||||
| Tanaitae Schoenherr, 1839 | ||||
| Tanaini Schoenherr, 1839 | ||||
| 94 | Cretotanaos bontsaganensis Legalov, 2014 | Tadu | P2i | 103 |
| Rhadinocybitae Alonso-Zarazaga, 1992 | ||||
| Rhadinocybini Alonso-Zarazaga, 1992 | ||||
| 95 | Baltocyba electrinus Legalov, 2018 | PolJ | P2b | 104 |
| Notapionini Zimmerman, 1994 | ||||
| 96 | Archinvolvulus liquidus Voss, 1972 | ScanJ | P2b | 105 |
| Palaeotanaitae Kirejtshuk, Legalov et Nel, 2015 | ||||
| Palaeotanaini Kirejtshuk, Legalov et Nel, 2015 | ||||
| 97 | Palaeotanaos oisensis Kirejtshuk, Legalov et Nel, 2015 | OisJ | P2i | 8 |
| Aspidapiitae Alonso-Zarazaga, 1990 | ||||
| Aspidapiini Alonso-Zarazaga, 1990 | ||||
| 98 | Pseudaspidapion khnzoriani (Zherikhin, 1971) | BalJ | P2b | 79 |
| 99 | Baltoapion gusakovi (Legalov, 2015) | BalJ | P2b | 30 |
| 100 | B. subdiscedens (Voss, 1953) | BalJ | P2b | 80 |
| Kalcapiini Alonso-Zarazaga, 1990 | ||||
| 101 | Melanapion (Melanapionoides) poinari Legalov, 2015 | BalJ | P2b | 30 |
| 102 | M. (M.) wanati Legalov, 2012 | BalJ | P2b | 67 |
| 103 | Succinapion telnovi Legalov et Bukejs, 2014 | BalJ | P2b | 106 |
| Apionitae Schoenherr, 1823 | ||||
| Piezotrachelini Voss, 1959 | ||||
| 104 | Conapium alleni Legalov, 2012 | BalJ | P2b | 67 |
| 105 | Baltoconapium anderseni (Voss, 1972) | ScanJ | P2b | 105 |
| 106 | Electrapion kuntzeni (Wagner, 1924) | BalJ | P2b | 107 |
| Aplemonini Kissinger, 1968 | ||||
| 107 | Perapion menatensis Legalov, Kirejtshuk et Nel, 2017 | Mena | P1sl-t | 50 |
| 108 | P. rasnitsyni Legalov, 2018 | GreR | P2i-l | 69 |
| 109 | “P.“ profundum (Schlechtendal, 1894) | Rott | P3h | 89 |
| Apionini Schoenherr, 1823 | ||||
| Toxorhynchina Scudder, 1893 | ||||
| 110 | Apionion evestigatum (Scudder, 1893) | RoaM | P2i-l | 77 |
| 111 | Toxorhynchus europeoeocenicus Bukejs et Legalov, 2020 | RovJ | P2b | 108 |
| 112 | T. michalskii Legalov, in print. | PolJ | P2b | 109 |
| 113 | “T.“ arctus (Scudder, 1893) | Flor | P2p | 77 |
| 114 | “T.“ confectum (Scudder, 1893) | Flor | P2p | 77 |
| 115 | “T.“ corruptus (Scudder, 1893) | Flor | P2p | 77 |
| 116 | T. florissantensis (Wickham, 1916) | Flor | P2p | 100 |
| 117 | T. minusculus (Scudder, 1893 | Flor | P2p | 77 |
| 118 | T. oculatus (Scudder, 1893 | Flor | P2p | 77 |
| 119 | “T.” pumilum (Scudder, 1893) | Flor | P2p | 77 |
| 120 | “T.” refrenatum (Scudder, 1893) | Flor | P2p | 77 |
| 121 | “T.” reventus (Scudder, 1893) | Flor | P2p | 77 |
| 122 | T. scudderianum (Wickham, 1916) | Flor | P2p | 100 |
| Tribe incertae sedis | ||||
| 123 | “Apion” cockerelli Wickham, 1911 | Flor | P2p | 76 |
| 124 | “A.” obtusus (Scudder, 1893) | Flor | P2p | 77 |
| 125 | “A.” levirostre Foerster, 1891 | Brun | P3r | 87 |
| 126 | “A.” parvum Foerster, 1891 | Brun | P3r | 87 |
| 127 | “A.” sulcatum Foerster, 1891 | Brun | P3r | 87 |
| 128 | “Rhynchites” orcinus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| 129 | “Apion” primordiale Heyden et Heyden, 1866 | Rott | P3h | 85 |
| 130 | “A.” profundum Schlechtendal, 1894 | Rott | P3h | 89 |
| 131 | “Coeliodes” primigenius Oustalet, 1874 | Aix | P3h | 110 |
| Nanophyinae Gistel, 1856 | ||||
| Nanophyini Gistel, 1856 | ||||
| 132 | Palaeonanophyes zherikhini Legalov, 2015 | GreR | P2i-l | 111 |
| 133 | Baltonanophyes crassirostre Legalov, 2018 | PolJ | P2b | 104 |
| 134 | Zherikhiniellus rarus Legalov, in print. | BalJ | P2b | 109 |
| 135 | “Nanophyes” japetus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Corimaliini Alonso-Zarazaga, 1989 | ||||
| 136 | “Corimalia” cycloptera Theobald, 1937 | Cela | P2p | 112 |
| Brentinae Billberg, 1820 | ||||
| Trachelizini Lacordaire, 1865 | ||||
| Stereodermina Sharp, 1895 | ||||
| 137 | Cerobates (Cerobates) eocenicus Legalov et Wappler, in print. | Maar | P2l | 113 |
| Trachelizina Lacordaire, 1865 | ||||
| 138 | Eckfelderolispa petrefacta Legalov et Wappler, in print. | Maar | P2l | 113 |
| 139 | E. perita Legalov et Wappler, in print. | Maar | P2l | 113 |
| 140 | E. manderschieta Legalov et Wappler, in print. | Maar | P2l | 113 |
| Curculionidae Latreille, 1802 | ||||
| Erirhininae Schoenherr, 1825 | ||||
| Erirhinini Schoenherr, 1825 | ||||
| Erirhinina Schoenherr, 1825 | ||||
| 141 | Erirhinites bognorensis Britton, 1960 | LonC | P2i | 114 |
| 142 | “Procas” vinculatus Scudder 1893 | RoaM, | P2i-l, | 77 |
| WhiR | P3r | |||
| 143 | “P.” verberatus Scudder, 1893 | Flor | P2p | 77 |
| 144 | “Erycus” brevicollis Scudder, 1893 | Flor | P2p | 77 |
| Dorytomini Bedel, 1886 | ||||
| 145 | Dorytomus bukejsi Legalov, 2020 | BalJ | P2b | 71 |
| 146 | D. electrinus Legalov, 2016 | BalJ | P2b | 90 |
| 147 | D. groehni Bukejs et Legalov, 2019 | BalJ | P2b | 116 |
| 148 | D. korotyaevi Legalov, 2020 | BalJ | P2b | 71 |
| 149 | D. nudus Legalov, 2016 | BalJ | P2b | 115 |
| 150 | D. vlaskini Legalov, Nazarenko et Perkovsky, 2019 | RovJ | P2b | 117 |
| 151 | “D.” coercitus Scudder, 1893 | Flor | P2p | 77 |
| 152 | “D.” vulcanicus Wickham, 1912 | Flor | P2p | 101 |
| 153 | “D.” williamsi Scudder, 1893 | Flor | P2p | 77 |
| Bagoini C.G. Thomson, 1859 | ||||
| 154 | “Bagous” palintonus Foerster, 1891 | Brun | P3r | 87 |
| Molytinae Schoenherr, 1823 | ||||
| Molytini Schoenherr, 1823 | ||||
| Hylobiina W. Kirby, 1837 | ||||
| 155 | Archaralites zherichini Legalov, 2010 | Arkh | P1d | 118 |
| 156 | Furhylobius troesteri Legalov, 2015 | Mors | P2i | 30 |
| 157 | Archaeoheilus packardii (Scudder, 1893) | GreR | P2i-l | 77 |
| 158 | A. provectus (Scudder, 1876) | GreR | P2i-l | 73 |
| 159 | A. scudderi Legalov, 2018 | GreR | P2i-l | 69 |
| 160 | A. ovalis Legalov, 2018 | GreR | P2i-l | 69 |
| 161 | A. deleticius (Scudder, 1893) | WhiR | P3r | 77 |
| 162 | A. lacoei (Scudder, 1893) | Flor | P2p | 77 |
| 163 | “Anisorhynchus” offosus Oustalet, 1870 | Core | P3r | 119 |
| 164 | “Hylobius” antiquus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Plinthini Lacordaire, 1863 | ||||
| Leiosomatina Reitter, 1913 | ||||
| 165 | Leiosoma klebsi Legalov, 2016 | BalJ | P2b | 115 |
| Pissodini Gistel, 1856 | ||||
| 166 | Lithopissodes luschitzensis Beier, 1952 | Luzi | P3h | 120 |
| Acicnemidini Lacordaire, 1865 | ||||
| 167 | Electrotribus henningseni (Voss, 1972) | ScanJ, PolJ | P2b | 105 |
| BalJ, PolJ | ||||
| 168 | E. theryi Hustache, 1942 | P2b | 121 | |
| =Paleopissodes weigangae Ulke, 1947 | 122 | |||
| =Anchorthorrhinus incertus Voss, 1953 | 80 | |||
| =Isalcidodes macellus Voss, 1953 | BalJ | 80 | ||
| 169 | E. wolfschwenningerae (Rheinheimer, 2007) | BalJ | P2b | 102 |
| 170 | E. rarus Legalov, 2020 | P2b | 71 | |
| Magdalini Pascoe, 1870 | ||||
| 171 | “Magdalis“ sedimentorum Scudder, 1893 | Flor | P2p | 77 |
| 172 | “M.“ moesta Schlechtendal, 1894 | Rott | P3h | 89 |
| 173 | “M.“ deucalionis (Heyden et Heyden, 1866) | Rott | P3h | 85 |
| 174 | “M.“ protogenius (Heyden et Heyden, 1866) | Rott | P3h | 85 |
| Cleogonini Gistel, 1856 | ||||
| 175 | Rhysosternum aeternabile Scudder, 1893 | Flor | P2p | 77 |
| 176 | Rh. longirostre Scudder, 1893 | Flor | P2p | 77 |
| 177 | “Conotrachelus” florissantensis Wickham, 1912 | Flor | P2p | 102 |
| Sciabregmini Legalov, Kirejtshuk et Nel, 2019 | ||||
| 178 | Sciabregma rugosa Scudder, 1893 | RoaM | P2i-l | 77 |
| 179 | S. squamosa Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| 180 | S. tenuicornis Cockerell, 1921 | GreR | P2i-l | 123 |
| Camptorhinini Lacordaire, 1865 | ||||
| 181 | Camptorrhinites orarius Britton, 1960 | LonC | P2i | 114 |
| 182 | Korystina gracilis Britton, 1960 | LonC | P2i | 114 |
| Aedemonini Faust, 1898 | ||||
| 183 | Electrorhinus friedhelmi Legalov, 2020 | BalJ | P2b | 71 |
| Cryptorhynchini Schoenherr, 1825 | ||||
| Cryptorhynchina Schoenherr, 1825 | ||||
| 184 | Taylorius litoralis Britton, 1960 | LonC | P2i | 114 |
| 185 | “Cryptorhynchus“ annosus Scudder, 1876 | GreR | P2i-l | 73 |
| 186 | “C.” coloradensis Wickham, 1912 | Flor | P2p | 102 |
| 187 | “C.” evinctus Scudder, 1893 | Flor | P2p | 77 |
| 188 | “C.” falli Wickham, 1912 | Flor | P2p | 102 |
| 189 | “C.” kerri Scudder, 1893 | Flor | P2p | 77 |
| 190 | “C.” profusus Scudder, 1893 | Flor | P2p | 77 |
| 191 | Oisecalles latosquamosus Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| 192 | Cryptorrhynchites sculpturatus Haupt, 1950 | Geis | P2l | 124 |
| 193 | Succinacalles uniqus Zherikhin, 1971 | BalJ | P2b | 79 |
| Tylodina Lacordaire, 1865 | ||||
| 194 | Baltacalles triumurbium Bukejs, Alekseev et Legalov, 2020 | BalJ | P2b | 125 |
| 195 | “Acalles” icarus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Tribe incertae sedis | ||||
| 196 | Lutago fetosus Britton, 1960 | LonC | P2i | 114 |
| 197 | L. nanus Britton, 1960 | LonC | P2i | 114 |
| 198 | Pissodites argillosus Britton, 1960 | LonC | P2i | 114 |
| 199 | “Chalcodermus” kirschi Deichmueller, 1881 | Kuts | P2b | 126 |
| 200 | Laccopygus nilesii Scudder, 1893 | Flor | P2p | 77 |
| 201 | L. rhenanus Meunier, 1924 | Rott | P3h | 127 |
| 202 | “Acalles” exhumatus Wickham, 1913 | Flor | P2p | 97 |
| 203 | “Pissodes” planatus Foerster, 1891 | Brun | P3r | 87 |
| 204 | “Hylobius” deletus Oustalet, 1870 | Core | P3r | 119 |
| 205 | “Plinthus” redivivus Oustalet, 1870 | Core | P3r | 119 |
| 206 | “Hylobius” morosus Oustalet, 1874 | Core | P3r | 110 |
| 207 | “Molytes” hassencampi Heyden, 1858 | Sieb | P3 | 128 |
| 208 | “Pissodes” effossus Heyden, 185 | Sieb | P3 | 128 |
| 209 | 8 “Hylobius” carbo Oustalet, 1874 | Aix | P3h | 110 |
| 210 | “Plinthus” heerii Oustalet, 1874 | Aix | P3h | 110 |
| Lixinae Schoenherr, 1823 | ||||
| Lixini Schoenherr, 1823 | ||||
| 212 | Lixus ligniticus Piton, 1940 | Mena | P1sl-t | 99 |
| 213 | “Larinus“ longirostris Foerster, 1891 | Brun | P3r | 87 |
| 214 | “L.“ bronni Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Cleonini Schoenherr, 1826 | ||||
| 215 | Eocleonus subjectus Scudder, 1893 | Flor | P2p | 77 |
| 216 | “Cleonus” degenerates Scudder, 1893 | Flor | P2p | 77 |
| 217 | “C.” estriatus Wickham, 1912 | Flor | P2p | 102 |
| 218 | “C.” exterraneus Scudder, 1893 | Flor | P2p | 77 |
| 219 | “C.” foersteri Scudder, 1893 | Flor | P2p | 77 |
| 220 | “C.” primoris Scudder, 1893 | Flor | P2p | 77 |
| 221 | “C.” rohweri Wickham, 1911 | Flor | P2p | 76 |
| 222 | “C.” arvernensis Oustalet, 1870 | Core | P3r | 119 |
| 223 | “C.” fouilhouxii Oustalet, 1870 | Core | P3r | 119 |
| Dryophthorinae Schoenherr, 1825 | ||||
| Stromboscerini Lacordaire, 1865 | ||||
| 224 | Palaeodexipeus kirejtshuki Legalov, 2016 | BalJ | P2b | 90 |
| 225 | Rovnoslonik damzeni Legalov, Nazarenko et Perkovsky, 2019 | RovJ | P2b | 117 |
| 226 | Stenommatomorphus hexarthrus Nazarenko, 2009 | RovJ | P2b | 129 |
| Dryophthorini Schoenherr, 1825 | ||||
| 227 | Rhinoporkus gratiosus Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| 228 | Lithophthorus rugosicollis Scudder, 1893 | Flor | P2p | 77 |
| 229 | Spodotribus terrulentus Scudder, 1893 | Flor | P2p | 77 |
| 230 | Dryophthorus incertus (Theobald, 1935) | Aix | P3h | 130 |
| Sphenophorini Lacordaire, 1865 | ||||
| Sphenophorina Lacordaire, 1865 | ||||
| 231 | “Scyphophorus“ fossionis Scudder, 1893 | Flor | P2p | 77 |
| 232 | S. laevis Scudder, 1893 | Flor | P2p | 77 |
| 233 | “S.” tertiarius Wickham, 1911 | Flor | P2p | 76 |
| 234 | Oryctorhinus tenuirostris Scudder, 1893 | Flor | P2p | 77 |
| 235 | “Sphenophorus“ proluviosus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Tribe incertae sedis | ||||
| 236 | Hipporhinops sternbergi Cockerell, 1926 | Flor | P2p | 131 |
| Cossoninae Schoenherr, 1825 | ||||
| Dryotribini LeConte, 1876 | ||||
| 237 | Ampharthropelma decipiens Voss, 1972 | ScanJ | P2b | 105 |
| 238 | Caulophilus martynovae Legalov, Nazarenko et Perkovsky, 2019 | RovJ | P2b | 117 |
| 239 | C. rarus Legalov, 2016 | BalJ | P2b | 115 |
| 240 | C. squamosus Legalov, 2016 | BalJ | P2b | 115 |
| 241 | C. sucinopunctatus (Kuska, 1992) | BalJ | P2b | 132 |
| 242 | C. zherikhini Nazarenko, Legalov et Perkovsky, 2011 | RovJ | P2b | 133 |
| 243 | Necrodryophthorus inquilinus Voss, 1953 | BalJ | P2b | 80 |
| 244 | Synommatodes patruelis (Voss, 1953) | BalJ | P2b | 80 |
| 245 | Electrocossonus kirejtshuki Legalov, 2020 | BalJ | P2b | 71 |
| Rhyncolini Gistel, 1856 | ||||
| Rhyncolina Gistel, 1856 | ||||
| 246 | Rhyncolus sitonifrons Zherikhin, 1992 | Rott | P3h | 98 |
| Cossonini Schoenherr, 1825 | ||||
| 247 | “Cossonus” devoratus Cockerell, 1925 | Sunc | P1d | 134 |
| 248 | “C.” rutus Scudder, 1893 | RoaM | P2i-l | 77 |
| 249 | “C.” gabbii Scudder, 1893 | Flor | P2p | 77 |
| 250 | C. robustus Meunier, 1916 | Aix | P3h | 135 |
| Conoderinae Schoenherr, 1833 | ||||
| Bariditae Schoenherr, 1836 | ||||
| Apostasimerini Schoenherr, 1844 | ||||
| Coelonertina Casey, 1922 | ||||
| 251 | Geraeus diruptus (Scudder, 1893) | GreR | P2i-l | 77 |
| 252 | G. anvilis Legalov, 2018 | GreR | P2i-l | 70 |
| 253 | G. fossilis Legalov, 2018 | GreR | P2i-l | 70 |
| 254 | G. antediluviana (Wickham, 1916) | Flor | P2p | 100 |
| 255 | G. hoveyi (Wickham, 1912) | Flor | P2p | 101 |
| 256 | G. hypogaeus (Wickham, 1916) | Flor | P2p | 100 |
| 257 | G. matura (Scudder, 1893) | Flor | P2p | 77 |
| 258 | G. obnuptus (Scudder, 1893) | Flor | P2p | 77 |
| 259 | G. schucherti (Wickham, 1912) | Flor | P2p | 101 |
| 260 | G. vulcanicus (Wickham, 1913) | Flor | P2p | 97 |
| 261 | Lithogeraeus greenriverensis Legalov, 2018 | GreR | P2i-l | 70 |
| 262 | L. circumscripta (Scudder, 1893) | RoaM | P2i-l | 77 |
| 263 | L. ancilla (Scudder, 1893) | RoaM | P2i-l | 77 |
| 264 | L. comminute (Scudder, 1893) | WhiR | P3r | 77 |
| 265 | L. damnata (Scudder, 1893) | Flor | P2p | 77 |
| 266 | L. cremastorhynchoides (Wickham, 1913) | Flor | P2p | 97 |
| 267 | L. florissantensis (Wickham, 1913) | Flor | P2p | 97 |
| 268 | L. nearctica (Wickham, 1916) | Flor | P2p | 100 |
| 269 | L. primalis (Wickham, 1917) | Flor | P2p | 136 |
| 270 | L. renovata (Wickham, 1916) | Flor | P2p | 100 |
| 271 | Nicentrus curvirostris Legalov, 2018 | GreR | P2i-l | 70 |
| 272 | Steganus barrandei Scudder, 1893 | RoaM | P2i-l | 77 |
| 273 | Miogeraeus recurrens Wickham, 1916 | Flor | P2p | 100 |
| 274 | “Pachybaris” rudis Wickham, 1912 | Flor | P2p | 96 |
| Baridini Schoenherr, 1836 | ||||
| Baridina Schoenherr, 1836 | ||||
| 275 | “Baris” palaeophilus Cockerell, 1920 | Bour | P2l | 137 |
| 276 | “B.” divisa Scudder, 1893 | Flor | P2p | 77 |
| 277 | “B.” harlani Scudder, 1893 | Flor | P2p | 77 |
| 278 | “B.” imperfecta Scudder, 1893 | Flor | P2p | 77 |
| 279 | “B.” naviculare (Foerster, 1891) | Brun | P3r | 87 |
| 280 | Catobaris coenosa Scudder, 1893 | Flor | P2p | 77 |
| Eurhinina Lacordaire, 1865 | ||||
| 281 | Eurhinus occultus Scudder, 1876 | Flor | P2p | 77 |
| Tribe incertae sedis | ||||
| 282 | “Ceutorhynchus” clausus Scudder, 1893 | Flor | P2p | 77 |
| Conoderintae Schoenherr, 1833 | ||||
| Conoderini Schoenherr, 1833 | ||||
| 283 | Jantarhinus compressus Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| Palaeomallerini Legalov, 2018 | ||||
| 284 | Palaeomallerus longirostris Legalov, 2018 | GreR | P2i-l | 70 |
| Ceutorhynchitae Gistel, 1848 | ||||
| Ceutorhynchini Gistel, 1848 | ||||
| 285 | “Ceutorhynchus” degravatus Scudder, 1893 | RoaM | P2i-l | 77 |
| 286 | “C.” eocenicus Cockerell, 1920 | Peck | P2i | 137 |
| 287 | C. alekseevi Legalov, 2016 | BalJ | P2b | 115 |
| 288 | C. electrinus Legalov, 2016 | BalJ | P2b | 115 |
| 289 | C. succinus Legalov, 2013 | BalJ | P2b | 47 |
| 290 | “C.” blaisdelli Wickham, 1916 | Flor | P2p | 100 |
| 291 | “C.” compactus Scudder, 1893 | Flor | P2p | 77 |
| 292 | “C.” duratus Scudder, 1893 | Flor | P2p | 77 |
| 293 | “C.” irvingi (Scudder, 1893) | Flor | P2p | 77 |
| 294 | “C.” fischeri Foerster, 1891 | Brun | P3r | 87 |
| 295 | “C.” crassirostris Foerster, 1891 | Brun | P3r | 87 |
| 296 | “C.” obliquus Foerster, 1891 | Brun | P3r | 87 |
| 297 | “C.” funeratus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| 298 | Baltocoeliodes sontagae Legalov et Bukejs, 2018 | BalJ | P2b | 138 |
| Cnemogonini Colonnelli, 1979 | ||||
| 299 | “Coeliodes” primotinus Scudder, 1893 | Flor | P2p | 77 |
| Phytobiini Gistel, 1848 | ||||
| 300 | “Ceuthorrhynchus”miegi Theobald, 1937 | Klei | P3 | 112 |
| Curculioninae Latreille, 1802 | ||||
| Acalyptini C.G. Thomson, 1859 | ||||
| 301 | Jantaronosik nebulosus Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| Ellescini C.G. Thomson, 1859 | ||||
| 302 | Succinostyphlus mroczkowskii Kuska, 1996 | BalJ | P2b | 139 |
| =Electrotribus erectosquamata Rheinheimer, 2007 | 122 | |||
| 303 | Pachytychius eocenicus Legalov, 2016 | BalJ | P2b | 115 |
| Palaeoanoplini Legalov, in print. | ||||
| 304 | Palaeoanoplus horridus Legalov, in print. | BalJ | P2b | 140 |
| Smicronychini Seidlitz, 1891 | ||||
| 305 | “Smicronyx” antiquus Foerster, 1891 | Brun | P3r | 87 |
| Curculionini Latreille, 1802 | ||||
| Erganiina Pelsue et O’Brien, 2011 | ||||
| 306 | Pseudoergania perkovskyi Legalov, 2019 | BalJ | P2b | 141 |
| Timolina Pelsue et O’Brien, 2011 | ||||
| 307 | Baltocurculio manukyani Legalov, 2020 | BalJ | P2b | 71 |
| Curculionina Latreille, 1802 | ||||
| 308 | Menatorhis elegans (Piton, 1940) | Mena | P1sl- | 99 |
| 309 | Curculio havighorstensis Zherikhin, 1995 | Havi | tP2i | 142 |
| 310 | C. anicularis (Scudder, 1893) | Flor | P2p | 77 |
| 311 | C. curvirostris (Scudder, 1893) | Flor | P2p | 77 |
| 312 | C. duttoni (Scudder, 1893) | Flor | P2p | 77 |
| 313 | C. extinctus (Wickham, 1912) | Flor | P2p | 102 |
| 314 | C. femoratus (Scudder, 1893) | Flor | P2p | 77 |
| 315 | C. flexirostris (Scudder, 1893) | Flor | P2p | 77 |
| 316 | C. florissantensis (Wickham, 1913) | Flor | P2p | 97 |
| 317 | C. minusculoides (Wickham, 1911) | Flor | P2p | 76 |
| 318 | C. minusculus (Scudder, 1893) | Flor | P2p | 77 |
| 319 | C. restrictus (Scudder, 1893) | Flor | P2p | 77 |
| 320 | “C.” beeklyi (Cockerell, 1918) | Flor | P2p | 143 |
| 321 | Numitor claviger Scudder, 1893 | Flor | P2p | 77 |
| Anthonomini C.G. Thomson, 1859 | ||||
| 322 | “Anthonomus” sunchalensis Cockerell, 1925 | Sunc | P1d | 134 |
| 323 | “A.” soporus Scudder, 1890 | GreR, RoaM, WhiR | P2i-l | 78 |
| GreR | P3r | |||
| Flor | ||||
| 324 | “A.” revictus Scudder, 1893 | Flor | P2i-l | 77 |
| 325 | “A.” concussus Scudder, 1893 | Flor | P2p | 77 |
| 326 | “A.” debilatus Scudder, 1893 | Flor | P2p | 77 |
| 327 | “A.” defossus Scudder, 1893 | Flor | P2p | 77 |
| 328 | “A.” evigilatus Scudder, 1893 | Flor | P2p | 77 |
| 329 | “A.” primordius Scudder, 1893 | Flor | P2p | 77 |
| 330 | “A.” rohweri Wickham, 1912 | Flor | P2p | 96 |
| 331 | Coccotorus principalis Scudder, 1893 | Flor | P2p | 77 |
| 332 | “C.” requiescens Scudder, 1893 | Flor | P2p | 77 |
| 333 | Cremastorhynchus stabilis Scudder, 1893 | P2p | 77 | |
| 334 | Smicrorhynchus macgeei Scudder, 1893 | P2p | 77 | |
| Eugnomini Lacordaire, 1863 | ||||
| 335 | Archaeoeugnomus balticus Legalov, 2016 | BalJ | P2b | 115 |
| 336 | Anthonoeugnomus barsevskisi Legalov, 2020 | BalJ | P2b | 71 |
| 337 | Mazurieugnomus pilosus Legalov, 2020 | BalJ | P2b | 71 |
| 338 | Groehnius electrum Bukejs et Legalov, 2019 | BalJ | P2b | 144 |
| 339 | G. parvum Legalov, 2020 | BalJ | P2b | 71 |
| Rhamphini Rafinesque, 1815 | ||||
| Palaeorhamphina Legalov, 2016 | ||||
| 340 | Palaeorhamphus damzeni Legalov, 2020 | BalJ | P2b | 71 |
| 341 | P. eichmanni Legalov, 2020 | BalJ | P2b | 71 |
| 342 | P. primitivus Legalov, 2016 | BalJ | P2b | 115 |
| Rhamphina Rafinesque, 1815 | ||||
| 343 | Orchestes tatjanae Legalov, 2016 | BalJ | P2b | 115 |
| 344 | “O.” languidulus Scudder, 1893 | Flor | P2p | 77 |
| 345 | Tachyerges hyperoche Legalov et Poinar, 2020 | PolJ | P2b | 91 |
| Tychiini C.G. Thomson, 1859 | ||||
| Tychiina C.G. Thomson, 1859 | ||||
| 346 | Eocenesibinia prussica Legalov, 2016 | BalJ | P2b | 115 |
| 347 | “Sibinia” whitneyi (Scudder, 1893) | Flor | P2p | 77 |
| 348 | “S.” melancholicus (Oustalet, 1874) | Aix | P3h | 110 |
| 349 | “Tychius” evolatus Scudder, 1893 | Flor | P2p | 77 |
| 350 | “T.” ferox Wickham, 1917 | Flor | P2p | 136 |
| 351 | “T.” secretus Scudder, 1893 | Flor | P2p | 77 |
| 352 | “T.” latus Foerster, 1891 | Brun | P3r | 87 |
| 353 | “T.” manderstjernai Heyden et Heyden, 1866 | Rott | P3h | 85 |
| 354 | Macrorhoptus intutus Scudder, 1893 | Flor | P2p | 77 |
| Camarotini Schoenherr, 1833 | ||||
| Prionomerina Lacordaire, 1863 | ||||
| 355 | Paleodontopus smirnovae Legalov, 2020 | BalJ | P2b | 71 |
| 356 | Masteutes rupis Scudder, 1893 | Flor | P2p | 77 |
| Curculioninae incertae sedis | ||||
| 357 | “Centrinus” longipes Piton, 1940 | Mena | P1sl-t | 99 |
| 358 | “Gymnetron” lecontei Scudder, 1878 | GreR | P2i-l | 74 |
| 359 | “G.” antecurrens Scudder, 1893 | Flor RovJ | P2p | 77 |
| 360 | “Protoceletes” hirtus Nazarenko et Perkovsky, 2016 | P2b | 145 | |
| Cyclominae Schoenherr, 1826 | ||||
| Listroderini LeConte, 1876 | ||||
| Palaechthina Brinck, 1948 | ||||
| 361 | “Listronotus” muratus Scudder, 1890 | GreR | P2i-l | 78 |
| Listroderina LeConte, 1876 | ||||
| 362 | “Listroderes” differens (Wickham, 1912) | Flor | P2p | 102 |
| 363 | “L.” eviscerates (Scudder, 1893) | Flor | P2p | 77 |
| Entiminae Schoenherr, 1823 | ||||
| Entimintae Schoenherr, 1823 | ||||
| Tropiphorini Marseul, 1863 | ||||
| 364 | Primocentron wickhami Legalov, 2018 | GreR | P2i-l | 69 |
| 365 | Limalophus poinari Legalov, 2020 | PolJ | P2b | 71 |
| 366 | Scuccinalophus attenboroughi Legalov, 2016 | BalJ | P2b | 90 |
| 367 | Limalophus compositus Scudder, 1893 | GreR, WhiR | P2i-l, | 77 |
| GreR | P3r | |||
| 368 | L. contractus Scudder, 1893 | WhiR | P2i-l | 77 |
| 369 | “L.” receptus Scudder, 1893 | WhiR | P3r | 77 |
| 370 | “Coniatus” refractus Scudder, 1893 | RoaM | P3r | 77 |
| 371 | “Cryptorhynchus” durus Scudder, 1893 | GreR | P2i-l | 77 |
| 372 | “Lepyrus” evictus Scudder, 1893 | GreR, RoaM | P2 i-l | 77 |
| 373 | “Sitona” paginarum Scudder, 1893 | RoaM | P2 i-l | 77 |
| 374 | RoaM GreR | |||
| 375 | “Otiorhynchus” subteractus Scudder, 1893 | RoaM | P2 i-l | 77 |
| 376 | “Ophryastes” grandis Scudder, 1893 | Flor | P2 i-l | 77 |
| 377 | “O.” compactus Scudder, 1878 | Flor | P2 i-l | 77 |
| 378 | “O.” petrarum Scudder, 1893 | Flor | P2 i-l | 77 |
| 379 | “O.” championi Wickham, 1912 | Flor | P2p | 102 |
| 380 | Geralophus antiquarius Scudder, 1893 | Flor | P2p | 77 |
| 381 | G. fossicius Scudder, 1893 | Flor | P2p | 77 |
| 382 | G. lassatus Scudder, 1893 | Flor | P2p | 77 |
| 383 | G. occultus Scudder, 1893 | Flor | P2p | 77 |
| 384 | G. pumiceus Scudder, 1893 | Flor | P2p | 77 |
| G. repositus Scudder, 1893 | Flor | P2p | 77 | |
| 385 | G. retritus Scudder, 1893 | Flor | P2p | 77 |
| 386 | G. saxuosus Scudder, 1893 | Flor | P2p | 77 |
| 387 | G. scudderi Wickham, 1911 | Aix | P2p | 76 |
| 388 | Centron moricollis Scudder, 1893 | P2p | 77 | |
| 389 | Tenillus firmus Scudder, 1893 | P2p | 77 | |
| 390 | Rhytideres sexsulcatus (Heer, 1856) | Aix | P3h | 146 |
| =Phytonomus annosus Oustalet, 1874 | 110 | |||
| =Hipporhinus reynesii Oustalet, 1874 | 110 | |||
| 391 | “Rh.” heeri (Germar, 1849) | P3h | 147 | |
| =Hipporhinus matheroni Nicolas, 1891 | 148 | |||
| =Hipporhinus similis Nicolas, 1891 | 148 | |||
| =Hipporhinus intermedius Nicolas, 1891 | Aix | 148 | ||
| =Hipporhinus marioni Nicolas, 1891 | Aix | 148 | ||
| =Hipporhinus pertonii Nicolas, 1891 | 148 | |||
| 392 | “Rh.” schaumi (Heer, 1856) | P3h | 146 | |
| 393 | “Hipporhinus” brevis Giebel, 1856 | P3h | 149 | |
| Entimini Schoenherr, 1823 | ||||
| Entimina Schoenherr, 1823 | ||||
| 394 | Entimus primordialis Scudder, 1876 | GreR | P2i-l | 73 |
| Eudiagogini LeConte, 1874 | ||||
| 395 | Eudiagogus vossi Legalov, 2018 | GreR | P2i-l | 69 |
| 396 | Tolstonosik oisensis Legalov, Kirejtshuk et Nel, 2019 | OisJ | P2i | 81 |
| 397 | Oligocryptus sectus (Scudder, 1893) | Flor | P2p | 77 |
| 398 | Eudomus pinguis Scudder, 1893 | Flor | P2p | 77 |
| 399 | E. robustus Scudder, 1893 | Flor | P2p | 77 |
| Hyperitae Lacordaire, 1863 | ||||
| Hyperini Lacordaire, 1863 | ||||
| Cepurina Capiomont, 1867 | ||||
| 400 | Palaeophelypera kuscheli Legalov, 2013 | BalJ | P2b | 47 |
| 401 | “Geralophus” discessus Scudder, 1893 | Flor | P2p | 77 |
| 402 | Hyperites nadezhkini Zherikhin, 1989 | Biam | P2p | 150 |
| Otiorhynchitae Schoenherr, 1826 | ||||
| Hormorini Horn, 1876 | ||||
| 403 | “Hormorus” saxorum Scudder, 1893 | Flor | P2p | 77 |
| Cyphiceritae Lacordaire, 1863 | ||||
| Sciaphilini Sharp, 1891 | ||||
| 404 | “Mitostylus” obdurefactus (Scudder, 1893) | RoaM | P2i-l | 77 |
| 405 | “M.” seculorum (Scudder, 1890) | GreR | P2i-l | 78 |
| 406 | “M.” abacus (Scudder, 1893) | WhiR | P3r | 77 |
| Trachyphloeini Lacordaire, 1863 | ||||
| Pseudocneorrhinina Kono, 1930 | ||||
| 407 | Archaeocallirhopalus alekseevi Legalov et Bukejs, 2015 | BalJ | P2b | 151 |
| 408 | A. larssoni Legalov, 2013 | BalJ | P2b | 47 |
| Polydrusitae Schoenherr, 1823 | ||||
| Sitonini Gistel, 1856 | ||||
| 409 | Sitonitellus egregius (Haupt, 1956) | Geis | P2l | 152 |
| 410 | Sitona lata Theobald, 1937 | Klei | P3 | 112 |
| 411 | “S.” venustulus Heyden et Heyden, 1866 | Rott | P3h | 85 |
| Anypotactini Champion, 1911 | ||||
| 412 | Paonaupactus gracilis Legalov, Nazarenko et Perkovsky, 2019 | RovJ | P2b | 117 |
| 413 | P. katyae Legalov, Nazarenko and Perkovsky, 2019 | RovJ | P2b | 117 |
| 414 | P. microphthalmus (Zherikhin, 1971) | BalJ | P2b | 79 |
| 415 | P. sitonitoides Voss, 1953 | BalJ, | P2b | 80 |
| =Polydrosus scheelei Voss, 1953 | ScanJ, | 80 | ||
| =Pyllobius cephalotes Voss, 1972 | PolJ | 105 | ||
| =Otiorhynchus pellucidipes Voss, 1972 | 105 | |||
| 416 | P. sobrinus (Voss, 1972) | ScanJ, | P2b | 105 |
| Balj | ||||
| 417 | P. viridis (Wanat et Borowiec, 1986) | BalJ, PolJ | P2b | 153 |
| Naupactini Gistel, 1856 | ||||
| 418 | “Hipporhinus” ventricosus (Piton, 1940) | Mena | P1sl-t | 99 |
| 419 | Arostropsis groehni Yunakov et Kirejtshuk, 2011 | BalJ | P2b | 154 |
| 420 | A. gusakovi Legalov, 2020 | BalJ | P2b | 71 |
| 421 | A. perkovskyi Bukejs et Legalov, 2019 | RovJ | P2b | 155 |
| 422 | “Cyrtomon” subterraneus Wickham, 1911 | Flor | P2p | 76 |
| 423 | “C.” florissantensis Wickham, 1914 | Flor | P2p | 156 |
| Geonemini Gistel, 1856 | ||||
| 424 | “Epicaerus” dilapsus (Scudder, 1893) | GreR | P2i-l | 77 |
| 425 | “E.” effossus (Scudder, 1876) | GreR | P2i-l | 73 |
| 426 | “E.” eradicatus (Scudder, 1893) | WhiR | P2i-l | 77 |
| 427 | “E.” exanimis (Scudder, 1876) | GreR | P2i-l | 73 |
| 428 | “E.” excissus (Scudder, 1893) | RoaM | P2i-l | 77 |
| 429 | “E.” fodinarum (Scudder, 1893) | GreR | P2i-l | 77 |
| 430 | “E.” saxatilis (Scudder, 1876) | GreR | P2i-l | 73 |
| 431 | “E.” subterraneus (Scudder, 1893) | GreR | P2i-l | 77 |
| 432 | “E.” terrosus (Scudder, 1878) | RoaM, WhiR | P2i-l, P3r | 74 |
| 433 | “E.” evigoratus (Scudder, 1893) | WhiR | P3r | 77 |
| 434 | Evopes veneratus Scudder, 1893 | Flor | P2p | 77 |
| 435 | E. occubatus Scudder, 1893 | Flor | P2p | 77 |
| 436 | “Lachnopus” recuperatus Scudder, 1893 | Flor | P2p | 77 |
| 437 | “Omileus” evanidus Scudder, 1893 | Flor | P2p | 77 |
| Psallidiini Lacordaire, 1863 | ||||
| 438 | Trigonoscuta inventa Scudder, 1893 | Flor | P2p | 77 |
| Eustylini Lacordaire, 1863 | ||||
| 439 | Pseudophaops perditus (Scudder, 1876) | GreR | P2i-l | 73 |
| Polydrusini Schoenherr, 1823 | ||||
| 440 | Polydrusus archetypus Zherikhin, 1971 | BalJ | P2b | 79 |
| 441 | P. zherikhini Legalov, 2020 | BalJ | P2b | 71 |
| 442 | Archaeosciaphilus marshalli Legalov, 2012 | BalJ | P2b | 82 |
| Brachyderini Schoenherr, 1826 | ||||
| 443 | Palaeocrassirhinus messelensis Rheinheimer, 2007 | Mess | P2l | 102 |
| 444 | P. rugosithorax Rheinheimer, 2007 | Mess | P2l | 102 |
| 445 | Palaeocneorhinus messelensis Rheinheimer, 2007 | Mess | P2l | 102 |
| 446 | “Brachyderes” rugosus (Deichmueller, 1881) | Kuts | P2b | 126 |
| 447 | Brachymycterus curculionoides Heyden et Heyden, 1866 | Rott | P3h | 85 |
| =Eurychirus induratus Heyden et Heyden, 1866 | 85 | |||
| =Rhinocyllus improbus Heyden et Heyden, 1866 | 85 | |||
| =Varus ignotus Schlechtendal, 1894 | 89 | |||
| Tanymecini Lacordaire, 1863 | ||||
| Tainophthalmina Desbrochers des Loges, 1873 | ||||
| 448 | Protainophthalmus asperulus (Heer, 1856) | Aix | P3h | 146 |
| =Brachyderes aquisextanus Oustalet, 1874 | 110 | |||
| =Brachyderes longipes Oustalet, 1874 | 110 | |||
| = Cleonus marcelli Oustalet, 1874 | 110 | |||
| 449 | P. margarum (Germar, 1849) | Aix | P3h | 147 |
| =Sitona antiqua Giebel, 1856 | 149 | |||
| 450 | P. punctulatus (Nicolas, 1891) | Aix | P3h | 148 |
| 451 | P. regularis (Nicolas, 1891) | Aix | P3h | 148 |
| 452 | P. thaisi (Nicolas, 1891) | Aix | P3h | 148 |
| 453 | P. tuberculatus (Nicolas, 1891) | Aix | P3h | 148 |
| Pandeleteina Pierce, 1913 | ||||
| 454 | Pandeleteinus nudus Wickham, 1917 | Flor | P2p | 138 |
| Tribe incertae sedis | ||||
| 455 | “Sitona” exitiorum Scudder, 1893 | Flor | P3r | 77 |
| 456 | “Strophosomus” marcelini Theobald, 1937 | Cela | P2p | 112 |
| 457 | “Sciaphilus” nigrescens Theobald, 1937 | Cela | P2p | 112 |
| 458 | “?Argoptochus” incertus Theobard, 1937 | Cela | P2p | 112 |
| 459 | “Bagous” atavus Oustalet, 1870 | Core | P3r | 119 |
| 460 | “Brachycerus” exilis Germar, 1837 | Aix | P3h | 157 |
| 461 | “Phytonomus” firmus Heer, 1856 | Aix | P3h | 146 |
| Curculionidae incertae sedis | ||||
| 462 | Otiorhynchites williamsi Cockerell, 1943 | Mirs | P1 | 158 |
| 463 | O. aterrimus Cockerell, 1925 | Sunc | P1d | 134 |
| 464 | O. crassus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 465 | O. densepunctatus Haupt, 1956 | Geis | P2l | 152 |
| 466 | O. fossilis Scudder, 1893 | GreR | P2i-l | 77 |
| 467 | O. commutatus Scudder, 1893 | RoaM | P2i-l | 77 |
| 468 | O. tysoni Scudder, 1893 | RoaM | P2i-l | 77 |
| 469 | O. absentivus Scudder, 1893 | Flor | P2p | 77 |
| 470 | O. florissantensis Wickham, 1911 | Flor | P2p | 76 |
| 471 | O. wilcoxianus Wickham, 1929 | Flor | P2p | 160 |
| 472 | Curculionites taxodii Heer, 1870 | Star | P1d | 161 |
| 473 | C. angustior Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 474 | C. epistictus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 475 | C. eustictus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 476 | C. harringtoni Cockerell, 1925 | Sunc | P1d | 134 |
| 477 | C. jujuensis Cockerell, 1925 | Sunc | P1d | 134 |
| 478 | C. latiusculus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 479 | C. magdalinus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 480 | C. megastictus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 481 | C. microstictus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 482 | C. parastictus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 483 | C. stebingeri Cockerell, 1926 | Sunc | P1d | 131 |
| 484 | C. sunchalicus Cockerell et Wagner, 1936 | Sunc | P1d | 159 |
| 485 | C. wielandi Cockerell, 1925 | Sunc | P1d | 134 |
| 486 | C. marginatus Giebel, 1856 | Corf | P2l | 149 |
| 487 | C. punctillatus Haupt, 1950 | Geis | P2l | 124 |
| 488 | C. bartonicus Cockerell, 1920 | Bour | P2l | 162 |
| 489 | C. brenthiformis Cockerell, 1920 | Bour | P2l | 162 |
| 490 | C. optimus Cockerell, 1920 | Bour | P2l | 162 |
| 491 | C. ovatus Oustalet, 1870 | Core | P3r | 119 |
| 492 | C. morosus Heer, 1856 | Aix | P3h | 146 |
| 493 | Calandrites defessus Scudder, 1893 | RoaM | P2i-l | 77 |
| 494 | C. cineratius Scudder, 1893 | GreR,RoaM | P2i-l | 77 |
| 495 | C. hindsi Cockerell, 1917 | Flor | P2p | |
| 496 | C. ursorum Cockerell, 1918 | Flor | P2p | |
| 497 | Ophryastites gardneri Cockerell, 1920 | Bour | P2l | |
| 498 | O. absconsus Scudder, 1893 | Flor | P2p | 77 |
| 499 | O. cinereus Scudder, 1893 | Flor | P2p | 77 |
| 500 | O. digressus Scudder, 1893 | Flor | P2p | 77 |
| 501 | O. hendersoni Cockerell, 1917 | Flor | P2p | |
| 502 | O. miocenus Wickham, 1912 | Flor | P2p | |
| 503 | O. dispertitus Scudder, 1893 | WhiR | P3r | 77 |
| 504 | O. heribaudi Piton, 1936 | PuStJ | P3 | |
| 505 | Mononychites punctipennis Haupt, 1956 | Geis | P2l | |
| 506 | M. rotundatus Haupt, 1956 | Geis | P2l | |
| 507 | Syntomostylus rudis Scudder, 1893 | RoaM | P2i-l | |
| 508 | “S.” fortis Cockerell, 1909 | GreR | P2i-l | |
| 509 | “Otiorhynchus” flaccus Scudder, 1893 | RoaM | P2i-l | 77 |
| 510 | “Otiorhynchus” tumbae Scudder, 1890 | GreR | P2i-l | |
| 511 | “Pachylobius” depraedatus Scudder, 1893 | RoaM | P2i-l | 77 |
| 512 | “Pachylobius” compressus Scudder, 1893 | GreR,RoaM | P2i-l | 77 |
| 513 | “Phyllobius” antecessor Scudder, 1893 | RoaM | P2i-l | 77 |
| 514 | “Rhyssomatus” tabescens Scudder, 1893 | WhiR | P3r | 77 |
| 515 | “Scythropus” somniculosus Scudder, 1893 | WhiR | P3r | 77 |
| 516 | “Phyllobius” avus Scudder, 1893 | RoaM | P2i-l | 77 |
| 517 | “Phyllobius” carcerarius Scudder, 1893 | RoaM | P2i-l | 77 |
| 518 | “Pachylobius” yungi Piton, 1936 | Gaub | P3 | 164 |
| 519 | “Tanymecus” gautieri Piton, 1936 | Gaub | P3 | 164 |
| 520 | “Pachylobius” martyi Piton et Theobald, 1937 | PuStJ | P3 | 166 |
| 521 | “Lachnopus” dilatatus Theobald, 1937 | Klei | P3 | 166 |
| 522 | “Cleonus” foersteri Theobald, 1937 | Klei | P3 | 112 |
| 523 | “Lachnopus” humatus Scudder, 1893 | Flor | P2p | 77 |
| 524 | “Sphenophorus” elegans Theobald, 1935 | Aix | P3h | 130 |
| Platypodidae Shuckard, 1840 | ||||
| Tesserocerinae Strohmeyer, 1914 | ||||
| Tesserocerini Strohmeyer, 1914 | ||||
| 525 | Eoplatypus jordali Peris, Solórzano Kraemer et Cognato, 2017 | BalJ | P2b | 167 |
| 526 | Cenocephalus aniskini Legalov, 2020 | BalJ | P2b | 71 |
| Scolytidae Latreille, 1804 | ||||
| Hylesininae Erichson, 1836 | ||||
| HylastiniLeConte, 1876 | ||||
| 527 | Hylastes aterites Schedl, 1947 | BalJ | P2b | 168 |
| 528 | “H.” americanus Wickham, 1913 | Flor | P2b | 97 |
| 529 | Hylurgops corpulentus Schedl, 1947 | BalJ | P2b | 168 |
| 530 | H. dubius (Hagedorn, 1906) | BalJ | P2b | 169 |
| 531 | H. electrinus (Germar, 1813) | BalJ | P2b | 170 |
| 532 | H. granulatus (Schedl, 1947) | BalJ | P2b | 168 |
| 533 | H. pilosellus Schedl, 1947 | BalJ | P2b | 168 |
| 534 | H. schellwieni (Hagedorn, 1906) | BalJ | P2b | 168 |
| 535 | H. tuberculatus Schedl, 1947 | BalJ | P2b | 168 |
| 536 | H. piger Wickham, 1913 | Flor | P2p | 97 |
| HylesininiErichson, 1836 | ||||
| 537 | Hylesinus extractus Scudder, 1893 | Flor | P2p | 77 |
| 538 | H. hydropicus (Wickham, 1916) | Flor | P2p | 101 |
| 539 | H. neli Petrov et Zherikhin, 2004 | Aix | P3h | 171 |
| 540 | “H.” facilis Heer, 1856 | Aix | P3h | 146 |
| Hylurgini Gistel, 1848 | ||||
| 541 | Xylechinus mozolevskae Petrov et Perkovsky, 2008 | RovJ | P2b | 172 |
| 542 | Klesovia pubescens Petrov et Perkovsky, 2018 | RovJ | P2b | 173 |
| 543 | Xylechinites anceps Hagedorn, 1906 | BalJ | P2b | 169 |
| Phloeosinini Nuesslin, 1912 | ||||
| 544 | Phloeosinus assimilis (Schedl, 1947) | BalJ | P2b | 168 |
| 545 | Ph. brunni (Hagedorn, 1906) | BalJ | P2b | 169 |
| 546 | Ph. regimontanus (Hagedorn, 1906) | BalJ | P2b | 169 |
| 547 | Ph. rehi (Hagedorn, 1906) | BalJ | P2b | 169 |
| 548 | Ph. robustus (Schedl, 1947) | BalJ | P2b | 168 |
| 549 | Ph. sexspinosus (Schedl, 1947) | BalJ | P2b | 168 |
| 550 | Ph. tuberculifer (Schedl, 1947) | BalJ | P2b | 168 |
| 551 | Ph. wolffi (Schedl, 1947) | BalJ | P2b | 168 |
| Phloeotribini Chapuis, 1869 | ||||
| 552 | Phloeotribus zimmermani Wickham, 1916 | Flor | P2p | 101 |
| Polygraphini Chapuis, 1869 | ||||
| 553 | Carphoborus keilbachi (Schedl, 1947) | BalJ | P2b | 168 |
| 554 | C. posticus (Schedl, 1947) | BalJ | P2b | 168 |
| Scolytinae Latreille, 1804 | ||||
| DryocoetiniLindemann, 1877 | ||||
| 555 | Dryocoetes diluvialis (Wickham, 1916) | GreR BalJ | P2i-l P2b | 101 |
| 556 | Taphramites gnathotrichus Schedl, 1947 | RovJ | P2b | 168 |
| 557 | T. rovnoensis Petrov et Perkovsky, 2008 | BalJ | P2b | 172 |
| 558 | Taphrorychus immaturatus Schedl, 1947 | 168 | ||
| Incertae sedis | ||||
| 559 | “Dryocoetes” carbonarius Scudder, 1878 | GreR | P2i-l | 74 |
| 560 | “Trypodendron” impressus Scudder, 1876 | GreR | P2i-l | 73 |
| 561 | “Polygraphus” wortheni Scudder, 1893 | RoaM | P2i-l | 77 |
| 562 | Xyleborites longipennis Wickham, 1913 | Flor | P2p | 97 |
| 563 | Duartia pulchella Martins-Neto, 2001 | Fons | P2p | 174 |
| Curculionoidea incertae sedis | ||||
| 564 | Thryogenosoma cariniger (Motschulsky, 1857) | BalJ | P2b | 175 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legalov, A.A. Fossil History of Curculionoidea (Coleoptera) from the Paleogene. Geosciences 2020, 10, 358. https://doi.org/10.3390/geosciences10090358
Legalov AA. Fossil History of Curculionoidea (Coleoptera) from the Paleogene. Geosciences. 2020; 10(9):358. https://doi.org/10.3390/geosciences10090358
Chicago/Turabian StyleLegalov, Andrei A. 2020. "Fossil History of Curculionoidea (Coleoptera) from the Paleogene" Geosciences 10, no. 9: 358. https://doi.org/10.3390/geosciences10090358
APA StyleLegalov, A. A. (2020). Fossil History of Curculionoidea (Coleoptera) from the Paleogene. Geosciences, 10(9), 358. https://doi.org/10.3390/geosciences10090358