
Biomolecular Composition of Sea Ice Microalgae and Its Influence on Marine Biogeochemical Cycling and Carbon Transfer through Polar Marine Food Webs


Abstract
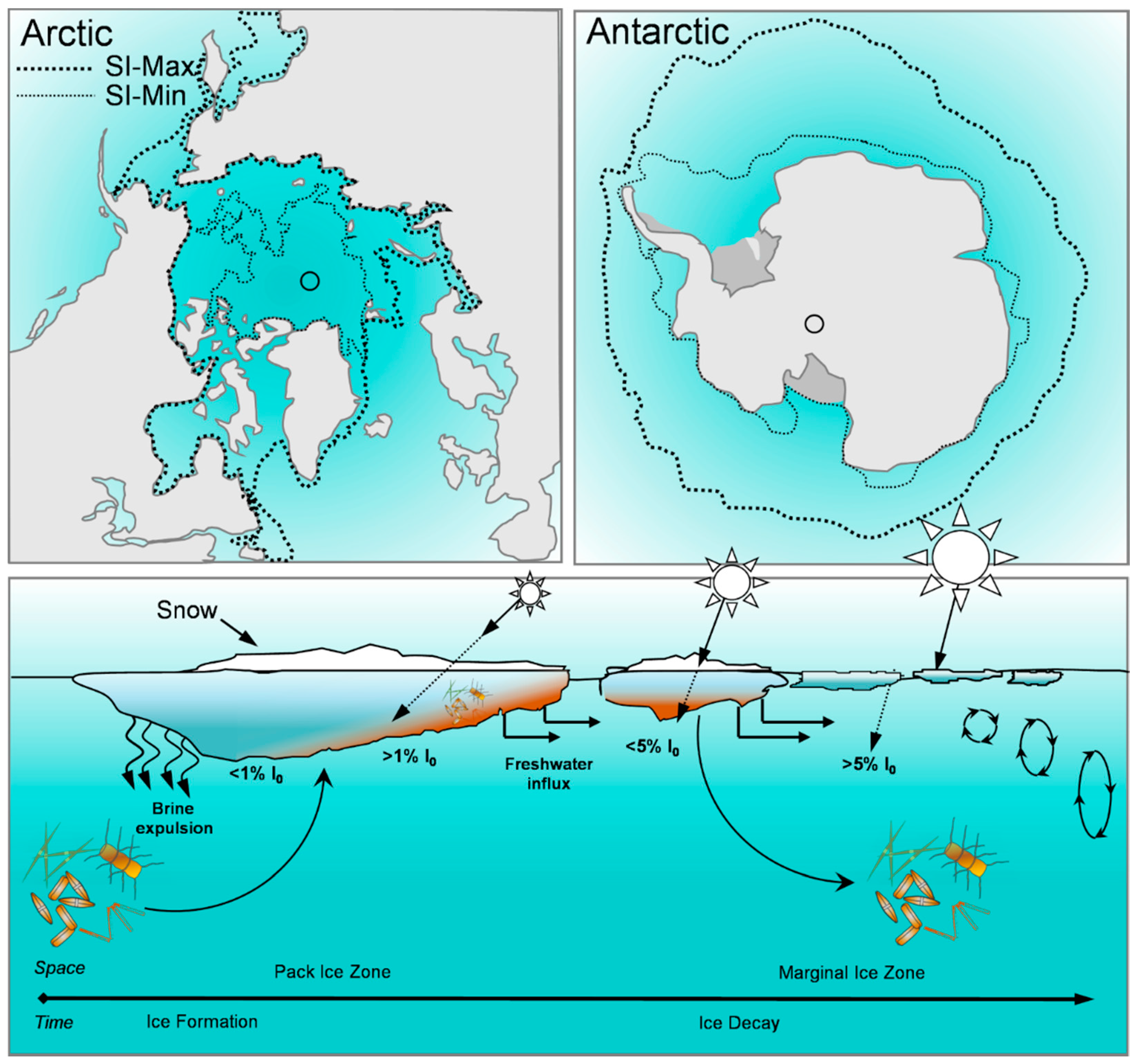
:1. Introduction
2. Biomolecular Composition of Sea Ice Algae from Polar Regions
3. Measuring Biochemical Composition in Microalgae
4. Environmental Factors That Influence Bioomolecular Composition
5. Sea Ice Microalgae Biochemistry and Carbon Transfer through the Polar Marine Food Web with Climate Change
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Parkinson, C.L. Global Sea Ice Coverage from Satellite Data: Annual Cycle and 35-Yr Trends. J. Clim. 2014, 27, 9377–9382. [Google Scholar] [CrossRef] [Green Version]
- Arrigo, K.R. Sea ice as a habitat for primary producers. In Sea Ice, 3rd ed.; Thomas, D., Ed.; Wiley-Blackwell: Oxford, UK, 2017; pp. 352–369. [Google Scholar] [CrossRef]
- Deming, J.W.; Eric Collins, R. Sea ice as a habitat for bacteria, archaea and viruses. In Sea Ice, 3rd ed.; Thomas, D., Ed.; Wiley-Blackwell: Oxford, UK, 2017; pp. 326–351. [Google Scholar] [CrossRef]
- Belt, S.T. Source-specific biomarkers as proxies for Arctic and Antarctic sea ice. Org. Geochem. 2018, 125, 277–298. [Google Scholar] [CrossRef] [Green Version]
- Kohlbach, D.; Hop, H.; Wold, A.; Schmidt, K.; Smik, L.; Belt, S.T.; Al-Habahbeh, A.K.; Woll, M.; Graeve, M.; Dąbrowska, A.M.; et al. Multiple trophic markers trace dietary carbon sources in barents sea zooplankton during late summer. Front. Mar. Sci. 2021, 7, 1216. [Google Scholar] [CrossRef]
- Rontani, J.-F.; Belt, S.T.; Brown, T.; Amiraux, R.; Gosselin, M.; Vaultier, F.; Mundy, C.J. Monitoring abiotic degradation in sinking versus suspended Arctic sea ice algae during a spring ice melt using specific lipid oxidation tracers. Org. Geochem. 2016, 98, 82–97. [Google Scholar] [CrossRef]
- Rontani, J.-F.; Amiraux, R.; Smik, L.; Wakeham, S.G.; Paulmier, A.; Vaultier, F.; Sun-Yong, H.; Jun-Oh, M.; Belt, S.T. Type II photosensitized oxidation in senescent microalgal cells at different latitudes: Does low under-ice irradiance in polar regions enhance efficiency? Sci. Total Environ. 2021, 779, 146363. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, S.; Xie, H.; Krotkov, N.; Larouche, P.; Vincent, W.F.; Babin, M. Photomineralization of terrigenous dissolved organic matter in Arctic coastal waters from 1979 to 2003: Interannual variability and implications of climate change. Glob. Biogeochem. Cycles 2006, 20, 1–13. [Google Scholar] [CrossRef]
- Massicotte, P.; Amon, R.M.W.; Antoine, D.; Archambault, P.; Balzano, S.; Bélanger, S.; Benner, R.; Boeuf, D.; Bricaud, A.; Bruyant, F.; et al. The MALINA oceanographic expedition: How do changes in ice cover, permafrost and UV radiation impact biodiversity and biogeochemical fluxes in the Arctic Ocean? Earth Syst. Sci. Data 2021, 13, 1561–1592. [Google Scholar] [CrossRef]
- Johnsen, G. Photoadaptation of sea-ice microalgae in the Barents Sea. Polar Biol. 1991, 11, 179–184. [Google Scholar] [CrossRef]
- Leu, E.; Mundy, C.; Assmy, P.; Campbell, K.; Gabrielsen, T.; Gosselin, M.; Juul-Pedersen, T.; Gradinger, R. Arctic spring awakening—Steering principles behind the phenology of vernal ice algal blooms. Prog. Oceanogr. 2015, 139, 151–170. [Google Scholar] [CrossRef]
- Cota, G.; Legendre, L.; Gosselin, M.; Ingram, R. Ecology of bottom ice algae: I. Environmental controls and variability. J. Mar. Syst. 1991, 2, 257–277. [Google Scholar] [CrossRef]
- Selz, V.; Laney, S.; Arnsten, A.E.; Lewis, K.M.; Lowry, K.E.; Joy-Warren, H.L.; Mills, M.M.; van Dijken, G.L.; Arrigo, K.R. Ice algal communities in the Chukchi and Beaufort Seas in spring and early summer: Composition, distribution, and coupling with phytoplankton assemblages. Limnol. Oceanogr. 2018, 63, 1109–1133. [Google Scholar] [CrossRef]
- Gosselin, M.; Levasseur, M.; Wheeler, P.A.; Horner, R.A.; Booth, B.C. New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 1997, 44, 1623–1644. [Google Scholar] [CrossRef]
- Legendre, L.; Ackley, S.F.; Dieckmann, G.S.; Horner, R.; Hoshiai, T.; Melnikov, I.A.; Reeburgh, W.S.; Spindler, M.; Sullivan, C.W. Ecology of sea ice biota. Polar Biol. 1992, 12, 429–444. [Google Scholar] [CrossRef]
- Gradinger, R. Sea-ice algae: Major contributors to primary production and algal biomass in the Chukchi and Beaufort Seas during May/June 2002. Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 1201–1212. [Google Scholar] [CrossRef]
- Fernández-Méndez, M.; Katlein, C.; Rabe, B.; Nicolaus, M.; Peeken, I.; Bakker, K.; Flores, H.; Boetius, A. Photosynthetic production in the central Arctic Ocean during the record sea-ice minimum in 2012. Biogeosciences 2015, 12, 3525–3549. [Google Scholar] [CrossRef] [Green Version]
- Rysgaard, S.; Kühl, M.; Glud, R.N.; Hansen, J.W. Biomass, production and horizontal patchiness of sea ice algae in a high-Arctic fjord (Young Sound, NE Greenland). Mar. Ecol. Prog. Ser. 2001, 223, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Whitledge, T.E.; Kang, S.-H. Spring time production of bottom ice algae in the landfast sea ice zone at Barrow, Alaska. J. Exp. Mar. Biol. Ecol. 2008, 367, 204–212. [Google Scholar] [CrossRef]
- Lizotte, M.P. The contributions of sea ice algae to Antarctic marine primary production1. Am. Zool. 2001, 41, 57–73. [Google Scholar] [CrossRef]
- Michel, C.; Legendre, L.; Ingram, R.G.; Gosselin, M.; Levasseur, M. Carbon budget of sea-ice algae in spring: Evidence of a significant transfer to zooplankton grazers. J. Geophys. Res. Earth Surf. 1996, 101, 18345–18360. [Google Scholar] [CrossRef]
- Riedel, A.; Michel, C.; Gosselin, M.; LeBlanc, B. Winter–spring dynamics in sea-ice carbon cycling in the coastal Arctic Ocean. J. Mar. Syst. 2008, 74, 918–932. [Google Scholar] [CrossRef]
- Arrigo, K.R.; Brown, Z.W.; Mills, M.M. Sea ice algal biomass and physiology in the Amundsen Sea, Antarctica. Elem. Sci. Anth. 2014, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Kohlbach, D.; Graeve, M.; Lange, B.A.; David, C.; Peeken, I.; Flores, H. The importance of ice algae-produced carbon in the central Arctic Ocean ecosystem: Food web relationships revealed by lipid and stable isotope analyses. Limnol. Oceanogr. 2016, 61, 2027–2044. [Google Scholar] [CrossRef]
- Søreide, J.E.; Leu, E.; Berge, J.; Graeve, M.; Falk-Petersen, S. Timing of blooms, algal food quality and Calanus glacialis reproduction and growth in a changing Arctic. Glob. Chang. Biol. 2010, 16, 3154–3163. [Google Scholar] [CrossRef] [Green Version]
- Leu, E.; Søreide, J.; Hessen, D.; Falk-Petersen, S.; Berge, J. Consequences of changing sea-ice cover for primary and secondary producers in the European Arctic shelf seas: Timing, quantity, and quality. Prog. Oceanogr. 2011, 90, 18–32. [Google Scholar] [CrossRef]
- Durbin, E.G.; Casas, M.C. Early reproduction by Calanus glacialis in the Northern Bering Sea: The role of ice algae as revealed by molecular analysis. J. Plankton Res. 2014, 36, 523–541. [Google Scholar] [CrossRef] [Green Version]
- Garrison, D.L.; Buck, K.R.; Fryxell, G.A. Algal assemblages in Antarctic pack ice and in ice-edge plankton 1. J. Phycol. 1987, 23, 564–572. [Google Scholar] [CrossRef]
- Michel, C.; Legendre, L.; Therriault, J.-C.; Demers, S.; Vandevelde, T. Springtime coupling between ice algal and phytoplankton assemblages in southeastern Hudson Bay, Canadian Arctic. Polar Biol. 1993, 13, 441–449. [Google Scholar] [CrossRef]
- Haecky, P.; Jonsson, S.; Andersson, A. Influence of sea ice on the composition of the spring phytoplankton bloom in the northern Baltic Sea. Polar Biol. 1998, 20, 1–8. [Google Scholar] [CrossRef]
- Jin, M.; Deal, C.; Wang, J.; Alexander, V.; Gradinger, R.; Saitoh, S.-I.; Iida, T.; Wan, Z.; Stabeno, P. Ice-associated phytoplankton blooms in the southeastern Bering Sea. Geophys. Res. Lett. 2007, 34, L006612. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, L.; Vichi, M.; Thomas, D.N. Process studies on the ecological coupling between sea ice algae and phytoplankton. Ecol. Model. 2012, 226, 120–138. [Google Scholar] [CrossRef]
- Szymanski, A.; Gradinger, R. The diversity, abundance and fate of ice algae and phytoplankton in the Bering Sea. Polar Biol. 2015, 39, 309–325. [Google Scholar] [CrossRef] [Green Version]
- Bhavya, P.S.; Kim, B.K.; Jo, N.; Kim, K.; Kang, J.J.; Lee, J.H.; Lee, D.; Lee, J.H.; Joo, H.; Ahn, S.H.; et al. A Review on the Macromolecular compositions of phytoplankton and the implications for aquatic biogeochemistry. Ocean Sci. J. 2018, 54, 1–14. [Google Scholar] [CrossRef]
- Bernard, K.S.; Gunther, L.A.; Mahaffey, S.H.; Qualls, K.M.; Sugla, M.; Saenz, B.T.; Cossio, A.M.; Walsh, J.; Reiss, C.S. The contribution of ice algae to the winter energy budget of juvenile Antarctic krill in years with contrasting sea ice conditions. ICES J. Mar. Sci. 2019, 76, 206–216. [Google Scholar] [CrossRef]
- Graeve, M.; Kattner, G.; Hagen, W. Diet-induced changes in the fatty acid composition of Arctic herbivorous copepods: Experimental evidence of trophic markers. J. Exp. Mar. Biol. Ecol. 1994, 182, 97–110. [Google Scholar] [CrossRef]
- Lee, R.; Hagen, W.; Kattner, G. Lipid storage in marine zooplankton. Mar. Ecol. Prog. Ser. 2006, 307, 273–306. [Google Scholar] [CrossRef]
- Hagen, W.; Auel, H. Seasonal adaptations and the role of lipids in oceanic zooplankton. Zoology 2001, 104, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Falk-Petersen, S.; Mayzaud, P.; Kattner, G.; Sargent, J.R. Lipids and life strategy of Arctic Calanus. Mar. Biol. Res. 2009, 5, 18–39. [Google Scholar] [CrossRef] [Green Version]
- Kohlbach, D.; Graeve, M.; Lange, B.A.; David, C.; Schaafsma, F.L.; Van Franeker, J.A.; Vortkamp, M.; Brandt, A.; Flores, H. Dependency of Antarctic zooplankton species on ice algae-produced carbon suggests a sea ice-driven pelagic ecosystem during winter. Glob. Chang. Biol. 2018, 24, 4667–4681. [Google Scholar] [CrossRef]
- Meredith, M.; Sommerkorn, M.; Cassotta, S.; Derksen, C.; Ekaykin, A.; Hollowed, A.; Kofinas, G.; Macintosh, A.; Melbourne-Thomas, J.; Muelbert, M.M.C.; et al. Polar Regions. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- Post, E.; Bhatt, U.S.; Bitz, C.M.; Brodie, J.F.; Fulton, T.L.; Hebblewhite, M.; Kerby, J.; Kutz, S.J.; Stirling, I.; Walker, D.A. Ecological consequences of sea-ice decline. Science 2013, 341, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Mallett, R.D.C.; Stroeve, J.C.; Tsamados, M.; Landy, J.C.; Willatt, R.; Nandan, V.; Liston, G.E. Faster decline and higher variability in the sea ice thickness of the marginal Arctic seas when accounting for dynamic snow cover. Cryosphere 2021, 15, 2429–2450. [Google Scholar] [CrossRef]
- Post, E.; Alley, R.B.; Christensen, T.R.; Macias-Fauria, M.; Forbes, B.C.; Gooseff, M.N.; Iler, A.; Kerby, J.T.; Laidre, K.L.; Mann, M.E.; et al. The polar regions in a 2 °C warmer world. Sci. Adv. 2019, 5, eaaw9883. [Google Scholar] [CrossRef] [Green Version]
- National Snow & Ice Data Center (NSIDC) Sea Ice Index. Available online: https://nsidc.org/data/seaice_index/ (accessed on 14 September 2021).
- Overland, J.E.; Wang, M. When will the summer Arctic be nearly sea ice free? Geophys. Res. Lett. 2013, 40, 2097–2101. [Google Scholar] [CrossRef]
- Haine, T.W.N.; Martin, T. The Arctic-Subarctic sea ice system is entering a seasonal regime: Implications for future Arctic amplification. Sci. Rep. 2017, 7, 4618. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, C.L. A 40-y record reveals gradual Antarctic sea ice increases followed by decreases at rates far exceeding the rates seen in the Arctic. Proc. Natl. Acad. Sci. USA 2019, 116, 14414–14423. [Google Scholar] [CrossRef] [Green Version]
- Stroeve, J.; Notz, D. Changing state of Arctic sea ice across all seasons. Environ. Res. Lett. 2018, 13, 103001. [Google Scholar] [CrossRef]
- Timmermans, M.; Marshall, J. Understanding Arctic Ocean circulation: A review of ocean dynamics in a changing climate. J. Geophys. Res. Oceans 2020, 125, 1–35. [Google Scholar] [CrossRef]
- Terhaar, J.; Kwiatkowski, L.; Bopp, L. Emergent constraint on Arctic Ocean acidification in the twenty-first century. Nature 2020, 582, 379–383. [Google Scholar] [CrossRef]
- Terhaar, J.; Orr, J.C.; Ethé, C.; Regnier, P.; Bopp, L. Simulated Arctic Ocean response to doubling of riverine carbon and nutrient delivery. Glob. Biogeochem. Cycles 2019, 33, 1048–1070. [Google Scholar] [CrossRef] [Green Version]
- Archer, S.; Leakey, R.; Burkill, P.; Sleigh, M.; Appleby, C. Microbial ecology of sea ice at a coastal Antarctic site: Community composition, biomass and temporal change. Mar. Ecol. Prog. Ser. 1996, 135, 179–195. [Google Scholar] [CrossRef]
- Becquevort, S.; Dumont, I.; Tison, J.-L.; Lannuzel, D.; Sauvée, M.-L.; Chou, L.; Schoemann, V. Biogeochemistry and microbial community composition in sea ice and underlying seawater off East Antarctica during early spring. Polar Biol. 2009, 32, 879–895. [Google Scholar] [CrossRef]
- Mock, T.; Kroon, B.M. Photosynthetic energy conversion under extreme conditions—II: The significance of lipids under light limited growth in Antarctic sea ice diatoms. Phytochemistry 2002, 61, 53–60. [Google Scholar] [CrossRef]
- Leu, E.; Wiktor, J.; Søreide, J.E.; Berge, J.; Falk-Petersen, S. Increased irradiance reduces food quality of sea ice algae. Mar. Ecol. Prog. Ser. 2010, 411, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Sackett, O.; Petrou, K.; Reedy, B.; De Grazia, A.; Hill, R.; Doblin, M.; Beardall, J.; Ralph, P.; Heraud, P. Phenotypic plasticity of southern ocean diatoms: Key to success in the sea ice habitat? PLoS ONE 2013, 8, e81185. [Google Scholar] [CrossRef]
- Sackett, O.; Petrou, K.; Reedy, B.; Hill, R.; Doblin, M.; Beardall, J.; Ralph, P.; Heraud, P. Snapshot prediction of carbon productivity, carbon and protein content in a Southern Ocean diatom using FTIR spectroscopy. ISME J. 2015, 10, 416–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogorzelec, N.; Mundy, C.; Findlay, C.; Campbell, K.; Diaz, A.; Ehn, J.; Rysgaard, S.; Gough, K. FTIR imaging analysis of cell content in sea-ice diatom taxa during a spring bloom in the lower northwest passage of the Canadian Arctic. Mar. Ecol. Prog. Ser. 2017, 569, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, C.; Nielsen, D.A.; Petrou, K. Macromolecular composition, productivity and dimethylsulfoniopropionate in Antarctic pelagic and sympagic microalgal communities. Mar. Ecol. Prog. Ser. 2020, 640, 45–61. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Follows, M.J.; Liefer, J.; Brown, C.M.; Benner, I.; Irwin, A.J. Phylogenetic Diversity in the Macromolecular Composition of Microalgae. PLoS ONE 2016, 11, e0155977. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.N.; Dieckmann, G.S. Antarctic sea ice—A habitat for extremophiles. Science 2002, 295, 641–644. [Google Scholar] [CrossRef] [Green Version]
- Morgan-Kiss, R.M.; Priscu, J.C.; Pocock, T.; Gudynaite-Savitch, L.; Huner, N.P.A. Adaptation and acclimation of photosynthetic microorganisms to permanently cold environments. Microbiol. Mol. Biol. Rev. 2006, 70, 222–252. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.V.; Follows, M.J.; Irwin, A.J. Size-scaling of macromolecules and chemical energy content in the eukaryotic microalgae. J. Plankton Res. 2016, 38, 1151–1162. [Google Scholar] [CrossRef] [Green Version]
- Van Oijen, T.; Van Leeuwe, M.A.; Granum, E.; Weissing, F.J.; Bellerby, R.G.J.; Gieskes, W.W.C.; De Baar, H.J.W. Light rather than iron controls photosynthate production and allocation in Southern Ocean phytoplankton populations during austral autumn. J. Plankton Res. 2004, 26, 885–900. [Google Scholar] [CrossRef] [Green Version]
- Ruess, L.; Müller-Navarra, D.C. Essential Biomolecules in Food Webs. Front. Ecol. Evol. 2019, 7, 269. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.M. Effect of growth rate of the food alga on the growth/ingestion efficiency of a marine herbivore. J. Mar. Biol. Assoc. UK 1980, 60, 681–702. [Google Scholar] [CrossRef]
- Lindqvist, K.; Lignell, R. Intracellular partitioning of 14c02 in phytoplankton during a growth season in the northern Baltic. Mar. Ecol. Prog. Ser. 1997, 152, 41–50. [Google Scholar] [CrossRef]
- Swalethorp, R.; Kjellerup, S.; Dünweber, M.; Nielsen, T.G.; Møller, E.F.; Rysgaard, S.; Hansen, B. Grazing, egg production, and biochemical evidence of differences in the life strategies of Calanus finmarchicus, C. glacialis and C. hyperboreus in Disko Bay, western Greenland. Mar. Ecol. Prog. Ser. 2011, 429, 125–144. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.K.; Lee, J.H.; Yun, M.S.; Joo, H.; Song, H.J.; Yang, E.J.; Chung, K.H.; Kang, S.-H.; Lee, S.H. High lipid composition of particulate organic matter in the northern Chukchi Sea, 2011. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 120, 72–81. [Google Scholar] [CrossRef]
- Yun, M.S.; Lee, D.B.; Kim, B.K.; Kang, J.J.; Lee, J.H.; Yang, E.J.; Park, W.G.; Chung, K.H.; Lee, S.H. Comparison of phytoplankton macromolecular compositions and zooplankton proximate compositions in the northern Chukchi Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 120, 82–90. [Google Scholar] [CrossRef]
- Smith, W.O.; Nelson, D.M.; DiTullio, G.R.; Leventer, A. Temporal and spatial patterns in the Ross Sea: Phytoplankton biomass, elemental composition, productivity and growth rates. J. Geophys. Res. Space Phys. 1996, 101, 18455–18465. [Google Scholar] [CrossRef]
- Fabiano, M.; Danovaro, R.; Povero, P. Vertical distribution and biochemical composition of pico- and microparticulate organic matter in the Ross Sea (Antarctica). In Oceanography of the Ross Sea Antarctica; Springer: Milano, Italy, 1999; pp. 233–246. [Google Scholar] [CrossRef]
- Kim, B.K.; Lee, J.H.; Joo, H.; Song, H.J.; Yang, E.J.; Lee, S.H. Macromolecular compositions of phytoplankton in the Amundsen Sea, Antarctica. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 123, 42–49. [Google Scholar] [CrossRef]
- Nelson, D.M.; Smith, W.O. Phytoplankton bloom dynamics of the western Ross Sea ice edge—II. Mesoscale cycling of nitrogen and silicon. Deep Sea Res. Part A Oceanogr. Res. Pap. 1986, 33, 1389–1412. [Google Scholar] [CrossRef]
- Pollard, R.; Tréguer, P.; Read, J. Quantifying nutrient supply to the Southern Ocean. J. Geophys. Res. Space Phys. 2006, 111, C0501. [Google Scholar] [CrossRef]
- Kim, B.K.; Lee, S.; Ha, S.-Y.; Jung, J.; Kim, T.W.; Yang, E.J.; Jo, N.; Lim, Y.J.; Park, J. Vertical distributions of macromolecular composition of particulate organic matter in the water column of the amundsen sea polynya during the summer in 2014. J. Geophys. Res. Oceans 2018, 123, 1393–1405. [Google Scholar] [CrossRef]
- Alderkamp, A.-C.; Buma, A.G.J.; van Rijssel, M. The carbohydrates of Phaeocystis and their degradation in the microbial food web. Biogeochemistry 2007, 83, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Hitchcock, G.L. A comparative study of the size-dependent organic composition of marine diatoms and dinoflagellates. J. Plankton Res. 1982, 4, 363–377. [Google Scholar] [CrossRef]
- Stehfest, K.; Toepel, J.; Wilhelm, C. The application of micro-FTIR spectroscopy to analyze nutrient stress-related change in biomass composition of phytoplankton algae. Plant Physiol. Biochem. 2005, 43, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R.J.; Nielsen, D.A.; Sheehan, C.E.; Deppeler, S.; Hancock, A.M.; Schulz, K.G.; Davidson, A.T.; Petrou, K. Ocean acidification alters the nutritional value of Antarctic diatoms. New Phytol. 2022, in press. [Google Scholar] [CrossRef]
- Gleitz, M.; Kirst, G. Photosynthesis-irradiance relationships and carbon metabolism of different ice algal assemblages collected from Weddell Sea pack ice during austral spring (EPOS 1). Polar Biol. 1991, 11, 385–392. [Google Scholar] [CrossRef]
- Smith, R.; Cavaletto, J.; Eadie, B.; Gardner, W. Growth and lipid composition of high Arctic ice algae during the spring bloom at Resolute, Northwest Territories, Canada. Mar. Ecol. Prog. Ser. 1993, 97, 19–29. [Google Scholar] [CrossRef]
- Henderson, R.J.; Hegseth, E.N.; Park, M.T. Seasonal variation in lipid and fatty acid composition of ice algae from the Barents Sea. Polar Biol. 1998, 20, 48–55. [Google Scholar] [CrossRef]
- Mock, T.; Kroon, B.M. Photosynthetic energy conversion under extreme conditions—I: Important role of lipids as structural modulators and energy sink under N-limited growth in Antarctic sea ice diatoms. Phytochemistry 2002, 61, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Teoh, M.-L.; Chu, W.-L.; Marchant, H.; Phang, S.-M. Influence of culture temperature on the growth, biochemical composition and fatty acid profiles of six Antarctic microalgae. J. Appl. Phycol. 2004, 16, 421–430. [Google Scholar] [CrossRef]
- Leu, E.; Falk-Petersen, S.; Kwaśniewski, S.; Wulff, A.; Edvardsen, K.; Hessen, D.O. Fatty acid dynamics during the spring bloom in a High Arctic fjord: Importance of abiotic factors versus community changes. Can. J. Fish. Aquat. Sci. 2006, 63, 2760–2779. [Google Scholar] [CrossRef]
- An, M.; Mou, S.; Zhang, X.; Ye, N.; Zheng, Z.; Cao, S.; Xu, D.; Fan, X.; Wang, Y.; Miao, J. Temperature regulates fatty acid desaturases at a transcriptional level and modulates the fatty acid profile in the Antarctic microalga Chlamydomonas sp. ICE-L. Bioresour. Technol. 2013, 134, 151–157. [Google Scholar] [CrossRef]
- Torstensson, A.; Hedblom, M.; Andersson, J.; Andersson, M.X.; Wulff, A. Synergism between elevated pCO2 and temperature on the Antarctic sea ice diatom Nitzschia lecointei. Biogeosciences 2013, 10, 6391–6401. [Google Scholar] [CrossRef]
- Torstensson, A.; Jiménez, C.; Nilsson, A.K.; Wulff, A. Elevated temperature and decreased salinity both affect the biochemical composition of the Antarctic sea-ice diatom Nitzschia lecointei, but not increased pCO2. Polar Biol. 2019, 42, 2149–2164. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Wang, Y.; Fan, X.; Wang, D.; Ye, N.; Zhang, X.; Mou, S.; Guan, Z.; Zhuang, Z. Long-Term Experiment on Physiological Responses to Synergetic Effects of Ocean Acidification and Photoperiod in the Antarctic Sea Ice Algae Chlamydomonas sp. ICE-L. Environ. Sci. Technol. 2014, 48, 7738–7746. [Google Scholar] [CrossRef] [PubMed]
- Lund-Hansen, L.; Hawes, I.; Hancke, K.; Salmansen, N.; Nielsen, J.; Balslev, L.; Sorrell, B. Effects of increased irradiance on biomass, photobiology, nutritional quality, and pigment composition of Arctic sea ice algae. Mar. Ecol. Prog. Ser. 2020, 648, 95–110. [Google Scholar] [CrossRef]
- Sargent, J.R.; Eilertsen, H.C.; Falk-Petersen, S.; Taasen, J.P. Carbon assimilation and lipid production in phytoplankton in northern Norwegian fjords. Mar. Biol. 1985, 85, 109–116. [Google Scholar] [CrossRef]
- Jonasdottir, S.; Visser, A.; Jespersen, C. Assessing the role of food quality in the production and hatching of Temora longicornis eggs. Mar. Ecol. Prog. Ser. 2009, 382, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Johns, L.; Wraige, E.; Belt, S.; Lewis, C.; Massé, G.; Robert, J.-M.; Rowland, S. Identification of a C25 highly branched isoprenoid (HBI) diene in Antarctic sediments, Antarctic sea-ice diatoms and cultured diatoms. Org. Geochem. 1999, 30, 1471–1475. [Google Scholar] [CrossRef]
- Brown, T.; Belt, S.T.; Tatarek, A.; Mundy, C.J. Source identification of the Arctic sea ice proxy IP25. Nat. Commun. 2014, 5, 4197. [Google Scholar] [CrossRef]
- Belt, S.T.; Brown, T.; Smik, L.; Tatarek, A.; Wiktor, J.; Stowasser, G.; Assmy, P.; Allen, C.S.; Husum, K. Identification of C25 highly branched isoprenoid (HBI) alkenes in diatoms of the genus Rhizosolenia in polar and sub-polar marine phytoplankton. Org. Geochem. 2017, 110, 65–72. [Google Scholar] [CrossRef]
- Palmisano, A.C.; Sullivan, C.W. Pathways of photosynthetic carbon assimilation in sea-ice microalgae from McMurdo Sound, Antarctica. Limnol. Oceanogr. 1985, 30, 674–678. [Google Scholar] [CrossRef]
- Smith, R.E.H.; Clement, P.; Cota, G.F.; Li, W.K.W. Intracellular photosynthate allocation and the control of arctic marine ice algal production1. J. Phycol. 2007, 23, 124–132. [Google Scholar] [CrossRef]
- Smith, R.; Herman, A. In situ patterns of intracellular photosynthate allocation by sea ice algae in the Canadian high arctic. Polar Biol. 1992, 12, 545–551. [Google Scholar] [CrossRef]
- Mock, T.; Gradinger, R. Changes in photosynthetic carbon allocation in algal assemblages of Arctic sea ice with decreasing nutrient concentrations and irradiance. Mar. Ecol. Prog. Ser. 2000, 202, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Li, W.K.W.; Glover, H.E.; Morris, I. Physiology of carbon photoassimilation by Oscillatoria thiebautii in the Caribbean Sea. Limnol. Oceanogr. 1980, 25, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Nichols, P.D.; Palmisano, A.C.; Rayner, M.S.; Smith, G.A.; White, D.C. Changes in the lipid composition of Antarctic sea-ice diatom communities during a spring bloom: An indication of community physiological status. Antarct. Sci. 1989, 1, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Leu, E.; Falk-Petersen, S.; Hessen, D.O. Ultraviolet radiation negatively affects growth but not food quality of arctic diatoms. Limnol. Oceanogr. 2007, 52, 787–797. [Google Scholar] [CrossRef]
- McConville, M.J.; Mitchell, C.; Wetherbee, R. Patterns of carbon assimilation in a microalgal community from annual sea ice, east Antarctica. Polar Biol. 1985, 4, 135–141. [Google Scholar] [CrossRef]
- Smith, R.E.H.; Clement, P.; Head, E. Biosynthesis and photosynthate allocation patterns of arctic ice algae. Limnol. Oceanogr. 1989, 34, 591–605. [Google Scholar] [CrossRef]
- Cade-Menun, B.J.; Paytan, A. Nutrient temperature and light stress alter phosphorus and carbon forms in culture-grown algae. Mar. Chem. 2010, 121, 27–36. [Google Scholar] [CrossRef]
- Heraud, P.; Wood, B.R.; Tobin, M.J.; Beardall, J.; McNaughton, D. Mapping of nutrient-induced biochemical changes in living algal cells using synchrotron infrared microspectroscopy. FEMS Microbiol. Lett. 2005, 249, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Heraud, P.; Wood, B.R.; Beardall, J.; McNaughton, D. Probing the Influence of the Environment on Microalgae Using Infrared and Raman Spectroscopy. New Approaches Biomed. Spectrosc. 2007, 963, 85–106. [Google Scholar] [CrossRef]
- Dean, A.P.; Nicholson, J.M.; Sigee, D.C. Changing patterns of carbon allocation in lake phytoplankton: An FTIR analysis. Hydrobiologia 2011, 684, 109–127. [Google Scholar] [CrossRef]
- Fanesi, A.; Wagner, H.; Birarda, G.; Vaccari, L.; Wilhelm, C. Quantitative macromolecular patterns in phytoplankton communities resolved at the taxonomical level by single-cell Synchrotron FTIR-spectroscopy. BMC Plant Biol. 2019, 19, 142. [Google Scholar] [CrossRef] [PubMed]
- Reffner, J.A.; Martoglio, P.A.; Williams, G.P. Fourier transform infrared microscopical analysis with synchrotron radiation: The microscope optics and system performance (invited). Rev. Sci. Instrum. 1995, 66, 1298–1302. [Google Scholar] [CrossRef]
- Giordano, M.; Kansiz, M.; Heraud, P.; Beardall, J.; Wood, B.; McNaughton, D. Fourier transform infrared spectroscopy as a novel tool to investigate changes in intracellular macromolecular pools in the marine microalga Chaetoceros muellerii (bacillariophyceae). J. Phycol. 2001, 37, 271–279. [Google Scholar] [CrossRef]
- Wagner, H.; Liu, Z.; Langner, U.; Stehfest, K.; Wilhelm, C. The use of FTIR spectroscopy to assess quantitative changes in the biochemical composition of microalgae. J. Biophotonics 2010, 3, 557–566. [Google Scholar] [CrossRef]
- Sigee, D.C.; Bahrami, F.; Estrada, B.; Webster, R.E.; Dean, A.P.; Elizabeth, R.; Peter Dean, A.; Sigee, D.C.; Bahrami, F.; Estrada, B.; et al. The influence of phosphorus availability on carbon allocation and P quota in Scenedesmus subspicatus: A synchrotron-based FTIR analysis. Phycologia 2007, 46, 583–592. [Google Scholar] [CrossRef]
- Findlay, C.; Morrison, J.; Mundy, C.J.; Sedlmair, J.; Hirschmugl, C.J.; Gough, K.M. Thermal source Fourier transform infrared microtomography applied to Arctic sea ice diatoms. Analyst 2017, 142, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Sackett, O.; Armand, L.; Beardall, J.; Hill, R.; Doblin, M.; Connelly, C.; Howes, J.; Stuart, B.; Ralph, P.; Heraud, P. Taxon-specific responses of Southern Ocean diatoms to Fe enrichment revealed by synchrotron radiation FTIR microspectroscopy. Biogeosciences 2014, 11, 5795–5808. [Google Scholar] [CrossRef] [Green Version]
- Boetius, A.; Anesio, A.M.; Deming, J.M.; Mikucki, J.A.; Rapp, J.Z. Microbial ecology of the cryosphere: Sea ice and glacial habitats. Nat. Rev. 2015, 13, 677–690. [Google Scholar] [CrossRef]
- Gleitz, M.; Thomas, D.N. Variation in phytoplankton standing stock, chemical composition and physiology during sea-ice formation in the southeastern Weddell Sea, Antarctica. J. Exp. Mar. Biol. Ecol. 1993, 173, 211–230. [Google Scholar] [CrossRef]
- Thomas, D.N.; Dieckmann, G.S. Biogeochemistry of antarctic sea ice. In Oceanography and Marine Biology, an Annual Review; Gibson, R.N., Barnes, M., Atkinson, A., Taylor, R.J., Eds.; CRC Press: Boca Raton, FL, USA, 2002; Volume 40, pp. 151–156. ISBN 9780429217722. [Google Scholar]
- Jacobs, S.S.; Giulivi, C.F.; Mele, P.A. Freshening of the ross sea during the late 20th century. Science 2002, 297, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.A.; Holding, J.M.; Carmack, E.C. Understanding regional and seasonal variability is key to gaining a pan-arctic perspective on arctic ocean freshening. Front. Mar. Sci. 2020, 7, 606. [Google Scholar] [CrossRef]
- Lee, S.H.; Whitledge, T.E.; Kang, S.-H. Carbon uptake rates of sea ice algae and phytoplankton under different light intensities in a landfast sea ice zone, Barrow, Alaska. Arctic 2009, 61, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Leu, E.; Wängberg, S.-Å.; Wulff, A.; Falk-Petersen, S.; Ørbæk, J.B.; Hessen, D.O. Effects of changes in ambient PAR and UV radiation on the nutritional quality of an Arctic diatom (Thalassiosira antarctica var. borealis). J. Exp. Mar. Biol. Ecol. 2006, 337, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Budge, S.M.; Wooler, M.J.; Springer, A.M.; Iverson, S.J.; McRoy, C.P.; Divoky, G.J. Tracing carbon flow in an arctic marine food web using fatty acid-stable isotope analysis. Oecologia 2008, 157, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, D.; Denman, K.; Michel, C. Modeling ice algal growth and decline in a seasonally ice-covered region of the Arctic (Resolute Passage, Canadian Archipelago). J. Geophys. Res. Space Phys. 2005, 110, 1–17. [Google Scholar] [CrossRef]
- Rozanska, M.; Gosselin, M.; Poulin, M.; Wiktor, J.; Michel, C. Influence of environmental factors on the development of bottom ice protist communities during the winter–spring transition. Mar. Ecol. Prog. Ser. 2009, 386, 43–59. [Google Scholar] [CrossRef] [Green Version]
- Vancoppenolle, M.; Bopp, L.; Madec, G.; Dunne, J.; Ilyina, T.; Halloran, P.R.; Steiner, N. Future Arctic Ocean primary productivity from CMIP5 simulations: Uncertain outcome, but consistent mechanisms. Glob. Biogeochem. Cycles 2013, 27, 605–619. [Google Scholar] [CrossRef] [Green Version]
- Slagstad, D.; Wassmann, P.F.J.; Ellingsen, I. Physical constrains and productivity in the future Arctic Ocean. Front. Mar. Sci. 2015, 2, 85. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Yoshida, T.; Quigg, A. Photosynthetic performance, lipid production and biomass composition in response to nitrogen limitation in marine microalgae. Plant Physiol. Biochem. 2012, 54, 70–77. [Google Scholar] [CrossRef]
- Chen, J.-J.; Li, Y.-R.; Lai, W.-L. Application of experimental design methodology for optimization of biofuel production from microalgae. Biomass Bioenergy 2014, 64, 11–19. [Google Scholar] [CrossRef]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Nitrogen starvation-induced cellular crosstalk of ROS-scavenging antioxidants and phytohormone enhanced the biofuel potential of green microalga Acutodesmus dimorphus. Biotechnol. Biofuels 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, Z.U.; Anal, A.K. Enhanced lipid and starch productivity of microalga (Chlorococcum sp. TISTR 8583) with nitrogen limitation following effective pretreatments for biofuel production. Biotechnol. Rep. 2019, 21, e00298. [Google Scholar] [CrossRef]
- Dean, A.P.; Sigee, D.C.; Estrada, B.; Pittman, J.K. Using FTIR spectroscopy for rapid determination of lipid accumulation in response to nitrogen limitation in freshwater microalgae. Bioresour. Technol. 2010, 101, 4499–4507. [Google Scholar] [CrossRef] [PubMed]
- Berges, J.A.; Charlebois, D.O.; Mauzerall, D.C.; Falkowski, P.G. Differential Effects of Nitrogen Limitation on Photosynthetic Efficiency of Photosystems I and II in Microalgae. Plant Physiol. 1996, 110, 689–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berges, J.A.; Falkowski, P.G. Physiological stress and cell death in marine phytoplankton: Induction of proteases in response to nitrogen or light limitation. Limnol. Oceanogr. 1998, 43, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Peltomaa, E.T.; Aalto, S.L.; Vuorio, K.M.; Taipale, S.J. The Importance of Phytoplankton Biomolecule Availability for Secondary Production. Front. Ecol. Evol. 2017, 5, 128. [Google Scholar] [CrossRef] [Green Version]
- Hancock, A.M.; King, C.K.; Stark, J.S.; McMinn, A.; Davidson, A.T. Effects of ocean acidification on Antarctic marine organisms: A meta-analysis. Ecol. Evol. 2020, 10, 4495–4514. [Google Scholar] [CrossRef] [Green Version]
- Petrou, K.; Kranz, S.A.; Trimborn, S.; Hassler, C.S.; Blanco-Ameijeiras, S.; Sackett, O.; Ralph, P.J.; Davidson, A.T. Southern Ocean phytoplankton physiology in a changing climate. J. Plant Physiol. 2016, 203, 135–150. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 411–484. [Google Scholar]
- Bestion, E.; Haegeman, B.; Codesal, S.A.; Garreau, A.; Huet, M.; Barton, S.; Montoya, J.M. Phytoplankton biodiversity is more important for ecosystem functioning in highly variable thermal environments. Proc. Natl. Acad. Sci. USA 2021, 118, e2019591118. [Google Scholar] [CrossRef] [PubMed]
- Hancock, A.M.; Davidson, A.T.; McKinlay, J.; McMinn, A.; Schulz, K.G.; Enden, R.L.V.D. Ocean acidification changes the structure of an Antarctic coastal protistan community. Biogeosciences 2018, 15, 2393–2410. [Google Scholar] [CrossRef] [Green Version]
- Sugie, K.; Fujiwara, A.; Nishino, S.; Kameyama, S.; Harada, N. Impacts of temperature, CO2, and salinity on phytoplankton community composition in the Western Arctic Ocean. Front. Mar. Sci. 2020, 6, 821. [Google Scholar] [CrossRef] [Green Version]
- Moline, M.A.; Claustre, H.; Frazer, T.K.; Schofield, O.; Vernet, M. Alteration of the food web along the Antarctic Peninsula in response to a regional warming trend. Glob. Chang. Biol. 2004, 10, 1973–1980. [Google Scholar] [CrossRef]
- Li, W.K.W.; McLaughlin, F.A.; Lovejoy, C.; Carmack, E.C. Smallest Algae Thrive As the Arctic Ocean Freshens. Science 2009, 326, 539. [Google Scholar] [CrossRef] [Green Version]
- Montes-Hugo, M.; Doney, S.C.; Ducklow, H.W.; Fraser, W.; Martinson, D.; Stammerjohn, S.E.; Schofield, O. Recent changes in phytoplankton communities associated with rapid regional climate change along the Western Antarctic Peninsula. Science 2009, 323, 1470–1473. [Google Scholar] [CrossRef] [PubMed]
- Neeley, A.R.; Harris, L.A.; Frey, K.E. Unraveling phytoplankton community dynamics in the Northern Chukchi sea under sea-ice-covered and sea-ice-free conditions. Geophys. Res. Lett. 2018, 45, 7663–7671. [Google Scholar] [CrossRef]
- Boyd, C.M.; Heyraud, M.; Boyd, C.N. Feeding of the Antarctic Krill Euphausia superba. J. Crustac. Biol. 1984, 4, 123–141. [Google Scholar] [CrossRef]
- Quetin, L.B.; Ross, R.M. Feeding by Antarctic Krill, Euphausia superba: Does size matter? In Antarctic Nutrient Cycles and Food Webs; Springer: Berlin/Heidelberg, Germany, 1985; pp. 372–377. [Google Scholar] [CrossRef]
- Moline, M.A.; Claustre, H.; Frazerj, T.K.; Grzymskj, J.O.E.; Schofielly, O.; Verner, M. Changes in phytoplankton assemblages along the Antarctic Peninsula and potential implications for the antarctic food web. In Antarctic Ecosystems: Models for Wider Ecological Understanding; Davidson, W., Howard-Williams, C., Broady, P., Eds.; New Zealand Natural Sciences: Christchurch, New Zealand, 2001; pp. 263–271. [Google Scholar]
- Campbell, K.; Mundy, C.J.; Belzile, C.; Delaforge, A.; Rysgaard, S. Seasonal dynamics of algal and bacterial communities in Arctic sea ice under variable snow cover. Polar Biol. 2018, 41, 41–58. [Google Scholar] [CrossRef]
- Saggiomo, M.; Escalera, L.; Saggiomo, V.; Bolinesi, F.; Mangoni, O. Phytoplankton blooms below the antarctic landfast ice during the melt season between late spring and early summer. J. Phycol. 2020, 57, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Hygum, B.H.; Rey, C.; Hansen, B.W.; Tande, K. Importance of food quantity to structural growth rate and neutral lipid reserves accumulated in Calanus finmarchicus. Mar. Biol. 2000, 136, 1057–1073. [Google Scholar] [CrossRef]
- Thingstad, T.F.; Bellerby, R.G.J.; Bratbak, G.; Børsheim, K.Y.; Egge, J.K.; Heldal, M.; Larsen, A.; Neill, C.; Nejstgaard, J.; Norland, S.; et al. Counterintuitive carbon-to-nutrient coupling in an Arctic pelagic ecosystem. Nat. Cell Biol. 2008, 455, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Krause, J.W.; Duarte, C.M.; Marquez, I.A.; Assmy, P.; Fernández-Méndez, M.; Wiedmann, I.; Wassmann, P.; Kristiansen, S.; Agustí, S. Biogenic silica production and diatom dynamics in the Svalbard region during spring. Biogeosciences 2018, 15, 6503–6517. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhao, J.; Su, J.; Li, C.; Cheng, B.; Hui, F.; Yang, Q.; Shi, L. Spatial and Temporal variations in the extent and thickness of Arctic landfast ice. Remote Sens. 2019, 12, 64. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.; Phillips, T.; Marshall, G.J.; Hosking, J.S.; Pope, J.O.; Bracegirdle, T.J.; Deb, P. Unprecedented springtime retreat of Antarctic sea ice in 2016. Geophys. Res. Lett. 2017, 44, 6868–6875. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Saenz, B.; Scanniello, J.; Daly, K.; Ainley, D. Local climatology of fast ice in McMurdo Sound, Antarctica. Antarct. Sci. 2018, 30, 125–142. [Google Scholar] [CrossRef]
- Palmer, M.; Saenz, B.T.; Arrigo, K.R. Impacts of sea ice retreat, thinning, and melt-pond proliferation on the summer phytoplankton bloom in the Chukchi Sea, Arctic Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 105, 85–104. [Google Scholar] [CrossRef]
- Arrigo, K.R.; Van Dijken, G. Continued increases in Arctic Ocean primary production. Prog. Oceanogr. 2015, 136, 60–70. [Google Scholar] [CrossRef]
- Stammerjohn, S.E.; Massom, R.; Rind, D.; Martinson, D.G. Regions of rapid sea ice change: An inter-hemispheric seasonal comparison. Geophys. Res. Lett. 2012, 39, 6501. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Raymond, B.; Bracegirdle, T.J.; Chadès, I.; Fuller, R.; Shaw, J.; Terauds, A. Climate change drives expansion of Antarctic ice-free habitat. Nature 2017, 547, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; van Dijken, G.L.; Arrigo, K.R. Changes in phytoplankton concentration now drive increased Arctic Ocean primary production. Science 2020, 369, 198–202. [Google Scholar] [CrossRef]
- Kahru, M.; Brotas, V.; Manzano-Sarabia, M.; Mitchell, B.G. Are phytoplankton blooms occurring earlier in the Arctic? Glob. Chang. Biol. 2010, 17, 1733–1739. [Google Scholar] [CrossRef]
- McConnell, B.; Gradinger, R.; Iken, K.; Bluhm, B.A. Growth rates of arctic juvenile Scolelepis squamata (Polychaeta: Spionidae) isolated from Chukchi Sea fast ice. Polar Biol. 2012, 35, 1487–1494. [Google Scholar] [CrossRef]
- Koeller, P.; Fuentes-Yaco, C.; Platt, T.; Sathyendranath, S.; Richards, A.; Ouellet, P.; Orr, D.; Skúladóttir, U.; Wieland, K.; Savard, L.; et al. Basin-scale coherence in phenology of shrimps and phytoplankton in the North Atlantic Ocean. Science 2009, 324, 791–793. [Google Scholar] [CrossRef]
- Dünweber, M.; Swalethorp, R.; Kjellerup, S.; Nielsen, T.; Arendt, K.; Hjorth, M.; Tönnesson, K.; Møller, E.F. Succession and fate of the spring diatom bloom in Disko Bay, western Greenland. Mar. Ecol. Prog. Ser. 2010, 419, 11–29. [Google Scholar] [CrossRef]
- Henley, S.F.; Schofield, O.; Hendry, K.; Schloss, I.R.; Steinberg, D.K.; Moffat, C.; Peck, L.S.; Costa, D.; Bakker, D.; Hughes, C.; et al. Variability and change in the west Antarctic Peninsula marine system: Research priorities and opportunities. Prog. Oceanogr. 2019, 173, 208–237. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Study | Taxa | Location | Latitude, Longitude | Sampling Date | Biomolecules Investigated | |
|---|---|---|---|---|---|---|
| Antarctica | An et al., 2013 | Chlamydomonas sp. ICE-L | Zhongshan Research Station | 69° S, 77° E | N/A | Fatty acids |
| Cade-Menun & Paytan 2010 | Fragilariopsis curta, Fragilariopsis cylindrus, Nitzschia subcurvata, Phaeocystis Antarctica, Thalassiosira weissflogii, Dunaliella tertiolecta, Synechoccus sp. | Culture | N/A | N/A | Lipid, protein, carbohydrate | |
| Gleitz & Kirst 1991 | Diatom-dominated mixed community, primarily Nitzschia sp., Chaetoceros sp., Navicula sp., Corethron sp., Rhizosolenia sp., Amphiprora sp., Dactyliosolen sp., Synedropsis sp., Tropidoneis and Phaeocystis pouchetii | Weddell Sea | 58–63° S, 55–45° W | 1988/1989 | Lipid, amino acid, carbohydrate | |
| Mock & Kroon 2002a | Fragilariopsis curta, Navicula gelida var.-antarctica, Nitzschia medioconstricta | Weddell Sea | 70°02′ S, 06°00′ W | March–May 1999 | Lipid, protein | |
| Mock & Kroon 2002b | Fragilariopsis curta, Navicula gelida var.-antarctica, Nitzschia medioconstricta | Weddell Sea | 70°02′ S, 06°00′ W | March–May 1999 | Lipid, protein | |
| Palmisano & Sullivan 1985 | Diatom-dominated mixed community, primarily Pleurosigma sp., Nitzschia stellata, Berkeleya sp., Amphiprora kuferathii, Phaeocystis sp. and small centrics. | McMurdo Sound | 77° S, 166° W | November–December 1983 | Lipid, protein, polysaccharide | |
| Teoh et al., 2004 | Chlamydomonas sp. and Navicula sp. | Windmill Islands | 66°17′ S, 110°29′ E | N/A | Lipid, protein, carbohydrate, fatty acids | |
| Sackett et al., 2013 | Fragilariopsis cylindrus, Chaetoceros simplex and Pseudo-nitzschia subcurvata | Southern Ocean and Prydz Bay | 66° S, 147° E, 68° S, 73° E | N/A | Lipid, protein, carbohydrate, fatty acids, amino acids | |
| Xu et al., 2014 | Chlamydomonas sp. ICE-L | Zhongshan Research Station | 69° S, 77° E | N/A | Lipid, fatty acids | |
| Arctic | Lee et al., 2008a | Mixed community dominated by large chain-forming diatoms | Barrow, Alaska | 71°20′ N, 156°39′ W | April–June 2003 | Lipid, protein, polysaccharide |
| Lee et al., 2008b | Mixed community dominated by large chain-forming diatoms | Barrow, Alaska | 71°20′ N, 156°39′ W | February–June 2003 | Lipid, protein, polysaccharide | |
| Leu et al., 2006b | Thalassiosira antarctica var. borealis | Ny-Ålesund, Svalbard | 78°55′ N, 11°56′ E | May–June 2004 | Fatty acids | |
| Leu et al., 2010 | Diatom-dominated mixed community, primarily Nitzschia frigida, Navicula septentrionalis and Fragilariopsis cylindrus. | Ripfjorden, Svalbard |
80° N, 22° E | March–July 2007 | Fatty acids | |
| Lund-Hansen et al., 2020 | Mixed diatom-dominated community. Primarily Nitzschia frigida, Nitzschia longissima and Thalassiosira sp. | Kangerlussuaq West Greenland | 66°57′ N, 50°57′ W | March 2016 | Fatty acids | |
| Mock & Gradinger 2000 | Mixed community dominated by Nitzschia sp., Fragilariopsis sp. and Chaetoceros sp. | Barents Sea |
77°10′ N, 34°04′ E | May–June 1997 | Lipid, protein, polysaccharides | |
| Pogorzelec et al., 2017 | Nitzschia frigida, pennate ribbon colonies and Attheya sp. | Dease Strait, Nunavut, Canada |
69°1′ N, 105°19′ W | March–May 2014 | Lipid, protein | |
| Smith et al., 1987 | Mixed community | Resolute Passage, Canada |
74°41′ N, 95°50′ W | April–June 1985 | Lipid, protein, polysaccharides | |
| Smith et al., 1989 | Diatom-dominated mixed community, primarily Nitzschia frigida and Nitzschia grunowii | Central Canadian Arctic |
74°40′ N, 94°54′ W | April–May 1985; 1986 | Lipid, protein, amino acid, polysaccharide | |
| Smith et al., 1993 | Diatom-dominated mixed community | Resolute Passage, Canada |
74°41′ N, 95°50′ W | March–June 1989 | Lipid | |
| Smith & Herman 1992 | Diatom-dominated mixed community | Resolute Passage, Canada |
74°41′ N, 95°50′ W | May 1987, May–June 1988 | Lipid, protein, polysaccharide | |
| Søreide et al., 2010 | Diatom-dominated mixed community | Ripfjorden, Svalbard |
80°27′ N, 22°29′ E | March–July 2007 | Fatty acids | |
| Torstensson et al., 2013 | Nitzschia lecointei | Amundsen Sea | N/A | January 2011 | Fatty acids | |
| Torstensson et al., 2019 | Nitzschia lecointei | Amundsen Sea | N/A | N/A | Lipid, protein carbohydrate, fatty acids |
| Method | Biomolecules Investigated | Advantages | Disadvantages | Example Studies with Sea Ice Algae |
|---|---|---|---|---|
| High Performance/Thin Layer Chromatography (HPTLC/TLC) | Lipid Fatty acid Amino Acid Carbohydrate | Rapid and easy to run multiple samples in parallel. Useful for complex lipids (therefore most marine lipids). Can be used for small quantity of sample. | Reproducibility can be unreliable. Temperature gradients can exist across the plate resulting in partial distillation of the sample. The silica plate is not reusable. | Gleitz & Kirst 1991 Smith et al., 1993 Henderson et al., 1998 Mock & Kroon, 2002a,b |
| Gas Chromatography (GC) | Lipid Fatty acid Protein Carbohydrate | Highly sensitive Accurate and reproducible. Easy to couple with detection and quantification techniques. Can analyze all biomolecular classes at once. Effective with a very small amount of sample. | Requires sample to be volatile and therefore lipids need to be derivatized into Fatty Acid Methyl Esters (FAMEs). | Nichols et al., 1989 [103] Teoh et al., 2004 Leu et al., 2006a,b; 2007 [104]; 2010 Søreide et al., 2010 An et al., 2013 Xu et al., 2014 Lund-Hansen et al., 2020 Torstensson et al., 2013; 2019 |
| Mass Spectrometry (MS) | Lipid Protein Polysaccharide | Sensitive Accurate and reproducible. | Less sensitive than GC. Requires coupling with another technique, e.g., HPLC or radioisotope labelling | Lee et al., 2008a,b |
| Radioisotope Labelling | Lipid Protein Amino Acid Polysaccharide | Highly sensitive Accurate and reproducible. Rapid and easy to run multiple samples in parallel. | Requires additional measurements to determine absolute concentrations. Requires extraction of biomolecular classes. Requires correction for quenching. Requires training and precautions due to radioactive materials | McConville et al., 1985 [105] Palmisano & Sullivan 1985 Smith et al., 1987 Palmisano et al., 1988 Smith & Herman 1992 Mock & Gradinger 2000 |
| Fluorescent Dye (BIODIPY 505/515) | Lipid | Rapid Inexpensive Performed in vivo Has lipid specificity, only binding to lipid bodies and chloroplasts and no other cytoplasmic compartments. | Does not stain all microalgae successfully. Can be issues associated with fading (i.e., fluorescence extinction) | Xu et al., 2014 |
| Sulpho-phospho-vanillin (SPV) reaction | Lipid | Rapid High throughput Relatively easy to implement. Relatively cheap Requires a small amount of sample. | Requires a reference standard Color intensity varies between different lipids Requires a two-step reaction | Smith et al., 1989 |
| Lowry and Smith Assays | Protein | Highly sensitive Produces a linear response curve. Low protein-to-protein variation meaning higher accuracy in unknown protein samples. Widely used and well characterized. | Susceptible to interference by some common chemicals present in samples. Time sensitive during analysis. Lowry is more complicated with more steps than the Smith Assay. Destructive to proteins. | Smith et al., 1989 Mock & Kroon 2002a Torstensson et al., 2019 |
| Phenol-Sulfuric Acid Method | Carbohydrate | Rapid Relatively easy to implement. Accurate and reproducible. Widely used and well characterized. |
Phenol is a toxic compound posing health risks Non-stoichiometric method, meaning a calibration curve using a series of standards must be generated, limiting the analysis of more complex carbohydrates. | Smith et al., 1989 Torstensson et al., 2019 |
| Carbon-13 Nuclear Magnetic Resonance (13C NMR) Spectroscopy | Lipid Protein Carbohydrate | Can analyze all biomolecular classes at once. Provides intramolecular detail. | Low sensitivity Long duration of analysis. | Cade-Menun & Paytan 2010 |
| FTIR-microspectroscopy | Lipid Fatty acid Protein Amino acid Carbohydrate | Highly sensitive Can be used to analyze single cells giving species-specific results. Nondestructive. Ability to obtain data at multiple wavelengths simultaneously. Can analyze all biomolecular classes at once. Effective with a very small amount of sample. | Small size mounting chamber. Synchrotron light source is not readily available and is expensive. | Sackett et al., 2013 Pogorzelec et al., 2017 Sheehan et al., 2020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duncan, R.J.; Petrou, K. Biomolecular Composition of Sea Ice Microalgae and Its Influence on Marine Biogeochemical Cycling and Carbon Transfer through Polar Marine Food Webs. Geosciences 2022, 12, 38. https://doi.org/10.3390/geosciences12010038
Duncan RJ, Petrou K. Biomolecular Composition of Sea Ice Microalgae and Its Influence on Marine Biogeochemical Cycling and Carbon Transfer through Polar Marine Food Webs. Geosciences. 2022; 12(1):38. https://doi.org/10.3390/geosciences12010038
Chicago/Turabian StyleDuncan, Rebecca Julianne, and Katherina Petrou. 2022. "Biomolecular Composition of Sea Ice Microalgae and Its Influence on Marine Biogeochemical Cycling and Carbon Transfer through Polar Marine Food Webs" Geosciences 12, no. 1: 38. https://doi.org/10.3390/geosciences12010038
APA StyleDuncan, R. J., & Petrou, K. (2022). Biomolecular Composition of Sea Ice Microalgae and Its Influence on Marine Biogeochemical Cycling and Carbon Transfer through Polar Marine Food Webs. Geosciences, 12(1), 38. https://doi.org/10.3390/geosciences12010038