
Benthic Foraminifera as Environmental Indicators in Mediterranean Marine Caves: A Review




Abstract
:1. Introduction
1.1. General Characteristics of Marine Caves
1.2. Benthic Foraminifera as Ecological and Paleoecological Indicators
2. Reconstructing Paleoenvironments in the Present and Past Marine Caves
3. Characterizing Present Environments in Marine Caves
3.1. General Overview on Recent Benthic Foraminifera from Worldwide Marine Caves
3.2. The Mediterranean Caves
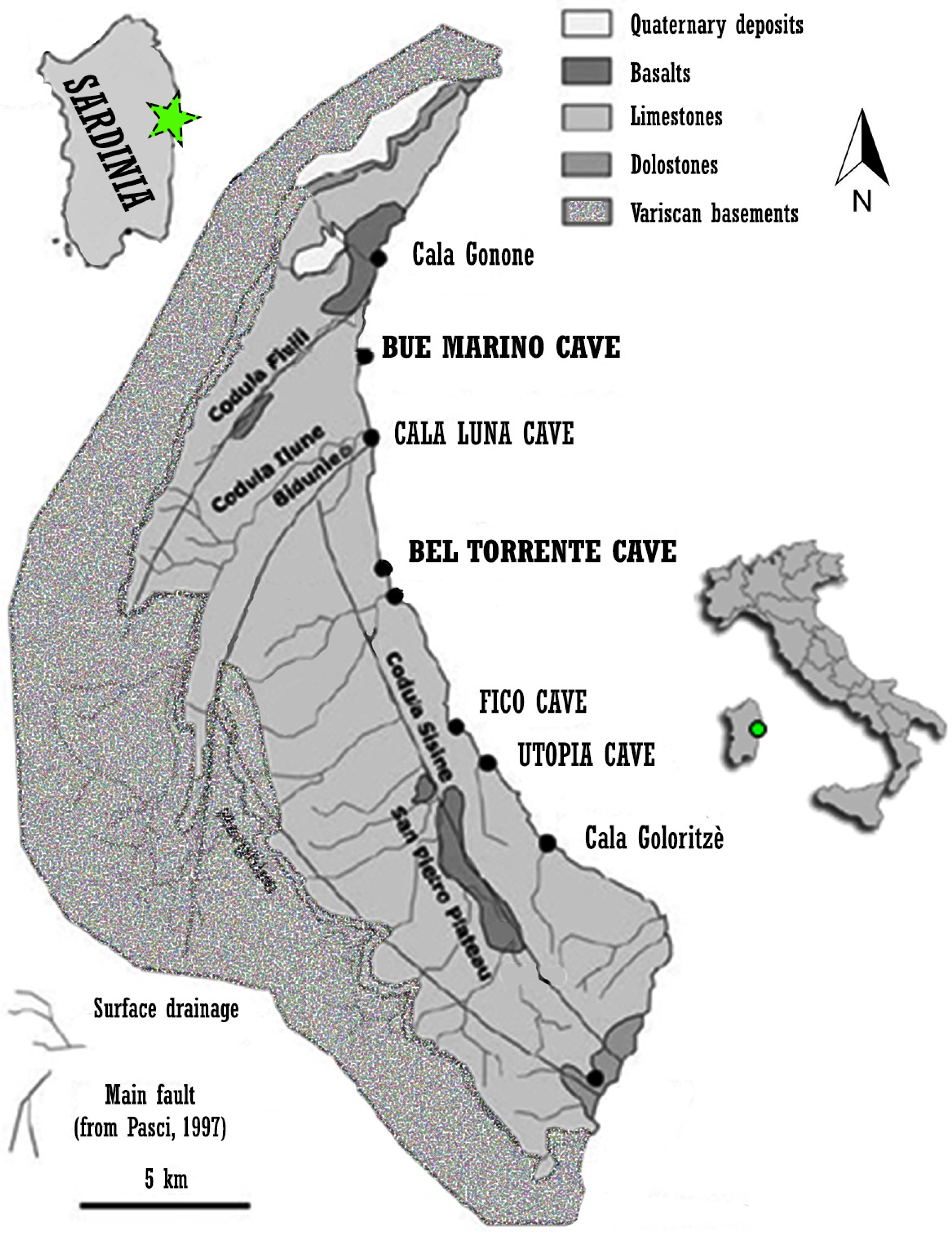
3.2.1. Studies Carried out in the Gulf of Orosei (Italy)
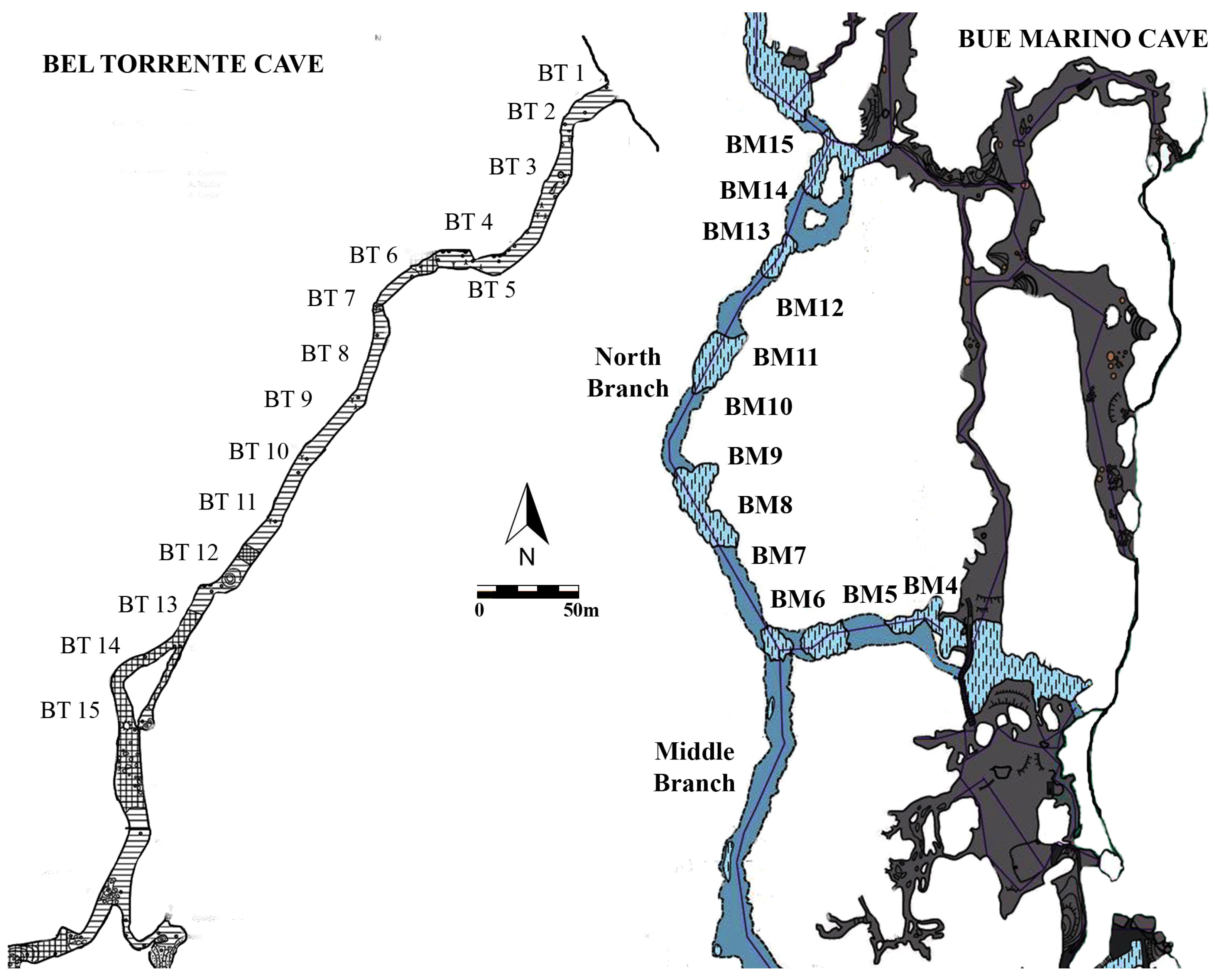
- A marine ecozone, located immediately close to the entrance, was exclusively characterized by taxa with carbonate test (porcelaneous or hyaline). Typical shallow-water symbiont-bearing species, such as Peneroplis pertusus, P. planatus, and Elphidium crispum, prevailed in the assemblage and were exclusive to this ecozone. Because these species are common in the shallow-water assemblages of the Sardinian coast, this assemblage indicates environmental conditions very similar to those that are normally marine.
- The entrance ecozone started within the first 90 m, in correspondence of BT4 and BM4. Hyaline taxa generally prevailed, but the agglutinated ones were also rather abundant. The dominant species, the hyaline Gavelinopsis praegeri and Rosalina bradyi, are epifaunal taxa that may be attached to phytal substrates or mineral grains. The first one is possibly living in a wide bathymetric range of the continental shelf. The hyaline infaunal B. variabilis accompanied them, and the agglutinated Reophax dentaliniformis was also abundant. It is an infaunal species common in the Baltic Sea and Norwegian fjords, recognized as a successful opportunistic species in re-colonizing disturbed habitats.
- The transitional ecozone started at 120 and 300 m from the entrance in correspondence of BT6 and BM11, respectively. The innermost foraminiferal assemblage characterized it, mainly constituted by agglutinated taxa. Eggerelloides advenus, Lepidodeuterammina ochracea, and R. dentaliniformis were the most common species. Eggerelloides advenus is an infaunal species typical of high latitude basins which tolerates low-salinity conditions, and is considered to be a stress-tolerant opportunistic species. Lepidodeuterammina ochracea is an epifaunal clinging species that is typical of cold waters and adapted to live on the coarse sand of high-energy cold waters. Station BM6, although included in this ecozone due to the prevalence of E. advenus, showed peculiar features in terms of the low species diversity and high dominance of the main species. In this case, E. advenus showed an opportunistic behavior due to the abundant vegetal debris present in the sediment. This species is known to be opportunistic, being able to benefit from the abundant refractory organic material.
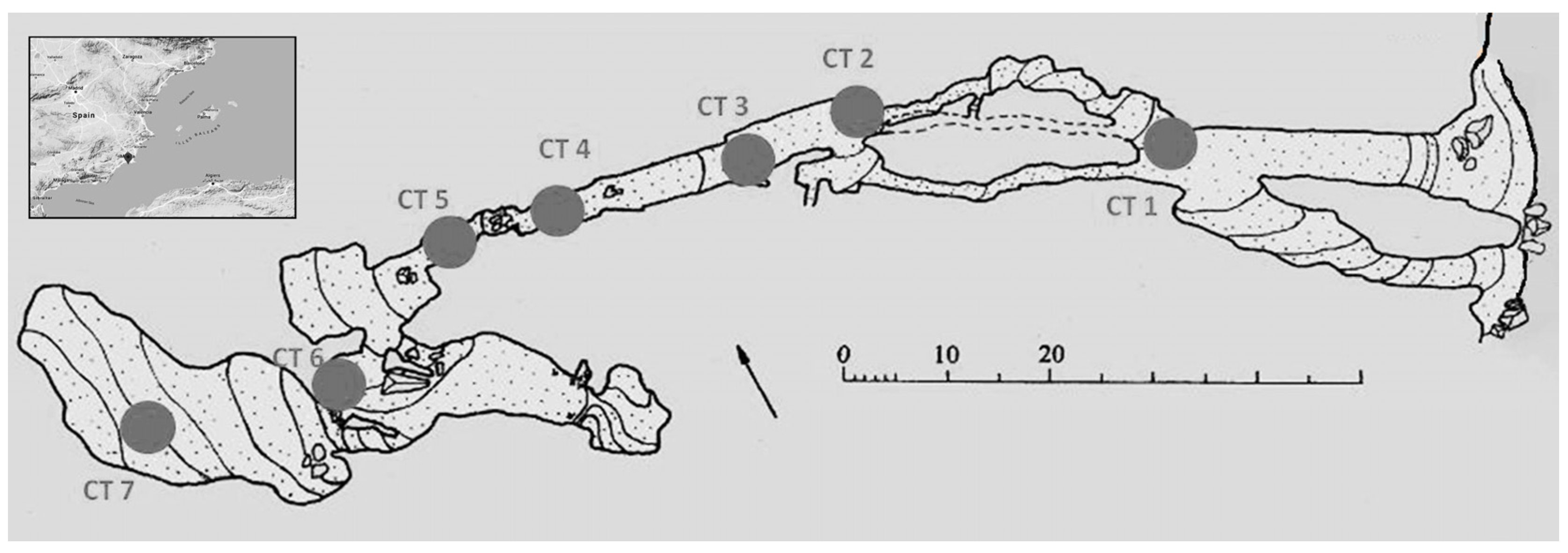
3.2.2. Studies Carried out in the Murcia Region (Spain)
4. Lessons Learned from Foraminiferal Studies in Marine Caves
Author Contributions
Funding
Conflicts of Interest
References
- Bakalowicz, M. Karst groundwater: A challenge for new resources. Hydrogeol. J. 2005, 13, 148–160. [Google Scholar] [CrossRef]
- Bakalowicz, M. Karst and karst groundwater resources in the Mediterranean. Environ. Earth Sci. 2015, 74, 5–14. [Google Scholar] [CrossRef]
- Goldscheider, N.; Chen, Z.; Auler, A.S.; Bakalowicz, M.; Broda, S.; Drew, D.; Hartmann, J.; Jiang, G.; Moosdorf, N.; Stevanovic, Z.; et al. Global distribution of carbonate rocks and karst water resources. J. Hydrogeol. 2020, 28, 1661–1677. [Google Scholar] [CrossRef] [Green Version]
- Stevanovic, Z. Global distribution and use of water from karst aquifers. In Advances in Karst Research: Theory, Fieldwork and Applications; Parise, M., Gabrovsek, F., Kaufmann, G., Ravbar, N., Eds.; Geological Society of London: London, UK, 2018; pp. 217–236. [Google Scholar]
- Stevanovic, Z. Karst waters in potable water supply: A global scale overview. Environ. Earth Sci. 2019, 78, 662. [Google Scholar] [CrossRef]
- Fleury, P.; Bakalowicz, M.; de Marsily, G. Submarine springs and coastal karst aquifers: A review. J. Hydrol. 2007, 339, 79–92. [Google Scholar] [CrossRef]
- Mijatovic, B. The groundwater discharge in the Mediterranean karst coastal zones and freshwater tapping: Set problems and adopted solutions. Case studies. Environ. Geol. 2007, 51, 737–742. [Google Scholar] [CrossRef]
- Parise, M.; Ravbar, N.; Živanovic, V.; Mikszewski, A.; Kresic, N.; Mádl-Szonyi, J.; Kukuric, N. Hazards in Karst and Managing Water Resources Quality. In Karst Aquifers—Characterization and Engineering; Professional Practice in Earth Sciences; Stevanovic, Z., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 601–687. [Google Scholar]
- Parise, M.; Gabrovsek, F.; Kaufmann, G.; Ravbar, N. Recent advances in karst research: From theory to fieldwork and applications. In Advances in Karst Research: Theory, Fieldwork and Applications; Parise, M., Gabrovsek, F., Kaufmann, G., Ravbar, N., Eds.; Geological Society of London: London, UK, 2018; pp. 1–24. [Google Scholar]
- Pinault, J.L.; Doerfliger, N.; Ladouche, B.; Bakalowicz, M. Characterizing a coastal karst aquifer using an inverse modeling approach: The saline springs of Thau, southern France. Water Resour. Res. 2004, 40. [Google Scholar] [CrossRef]
- Olarinoye, T.; Gleeson, T.; Marx, V.; Seeger, S.; Adinehvand, R.; Allocca, V.; Andreo, B.; Apaéstegui, J.; Apolit, C.; Arfib, B.; et al. Global karst springs hydrograph dataset for research and management of the world’s fastest-flowing groundwater. Sci. Data 2020, 7, 59. [Google Scholar] [CrossRef] [Green Version]
- Ford, D.C.; Williams, P. Karst Hydrogeology and Geomorphology; Wiley: Chichester, UK, 2007. [Google Scholar]
- Palmer, A.N. Cave Geology; Cave Books: Dayton, OH, USA, 2007. [Google Scholar]
- Harris, J.; Mylroie, J.; Carew, J. Banana holes: Unique karst features of the Bahamas. Carbonates Evaporites 1995, 10, 215–224. [Google Scholar] [CrossRef]
- Del Prete, S.; Iovine, G.; Parise, M.; Santo, A. Origin and distribution of different types of sinkholes in the plain areas of Southern Italy. Geodin. Acta 2010, 23, 113–127. [Google Scholar] [CrossRef]
- Parise, M. Hazards in karst. Sustainability of the karst environment. Dinaric karst and other karst regions. In Proceedings of the International Interdisciplinary Scientific Conference, Plitvice Lakes, Croatia, 23–26 September 2009; Bonacci, O., Ed.; Series on Groundwater no. 2. IHP-UNESCO: Paris, France, 2010; pp. 155–162. [Google Scholar]
- Parise, M. Sinkholes. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2019; pp. 934–942. [Google Scholar]
- Gutiérrez, F.; Parise, M.; De Waele, J.; Jourde, H. A review on natural and human-induced geohazards and impacts in karst. Earth-Sci. Rev. 2014, 138, 61–88. [Google Scholar] [CrossRef]
- Viles, H.A. Recent advances in field and experimental studies of rock weathering. Z. Geomorphol. 2000, 120, 343–368. [Google Scholar]
- Nicholson, D.T. Pore properties as indicator of breakdown mechanism in experimentally weathered limestones. Earth Surf. Process. Landf. 2001, 26, 819–838. [Google Scholar] [CrossRef]
- Gomez-Pujol, L.; Fornós, J.J.; Swantesson, J.O.H. Rock surface millimeter-scale roughness and weathering of supratidal Mallorcan carbonate coasts (Balearic Islands). Earth Surf. Process. Landf. 2006, 31, 1792–1801. [Google Scholar] [CrossRef]
- Calcaterra, D.; Parise, M. Weathering as a Predisposing Factor to Slope Movements; sp. publ. 23; Calcaterra, D., Parise, M., Eds.; Geological Society of London: London, UK, 2010; p. 233. [Google Scholar]
- Naylor, L.A.; Coombes, M.A.; Viles, H.A. Reconceptualising the role of organisms in the erosion of rock coasts: A new model. Geomorphology 2012, 157-158, 17–30. [Google Scholar] [CrossRef]
- Moses, C.; Robinson, D.; Barlow, J. Methods for measuring rock surface weathering and erosion: A critical review. Earth Sci. Rev. 2014, 135, 141–161. [Google Scholar] [CrossRef]
- Waelbroeck, C.; Labeyrie, L.; Michel, E.; Duplessy, J.C.; McManus, J.F.; Lambeck, K.; Balbon, E.; Labracherie, M. Sea-level and deep-water temperature changes derived from benthic foraminifera isotopic records. Quat. Sci. Rev. 2002, 21, 295–305. [Google Scholar] [CrossRef]
- Palmer, A.N. Origin and morphology of limestone caves. Geol. Soc. Am. Bull. 1991, 103, 1–21. [Google Scholar] [CrossRef]
- Klimchouk, A.B.; Ford, D.C.; Palmer, A.N.; Dreybrodt, W. Speleogenesis: Evolution of Karst Aquifers; National Speleological Society: Huntsville, AL, USA, 2000. [Google Scholar]
- Stock, J.H.; Iliffe, T.M.; Williams, D. The concept “anchialine” reconsidered. Stygologia 1986, 2, 90–92. [Google Scholar]
- van Hengstum, P.J.; Scott, D.B. Ecology of foraminifera and habitat variability in an underwater cave: Distinguishing anchialine versus submarine cave environments. J. Foraminifer. Res. 2011, 41, 201–229. [Google Scholar] [CrossRef] [Green Version]
- van Hengstum, P.J.; Scott, D.B.; Grocke, D.R.; Charette, M.A. Sea level controls sedimentation and environments in coastal caves and sinkholes. Mar. Geol. 2011, 286, 35–50. [Google Scholar] [CrossRef]
- Choquette, P.W.; Pray, L.C. Geologic nomenclature and classification of porosity in sedimentary carbonates. Amer. Ass. Petrol. Geol. Bull. 1970, 54, 207–250. [Google Scholar]
- Mylroie, J.E.; Mylroie, J.R. Void development on carbonate coasts: Creation of anchialine habitats. Hydrobiologia 2011, 677, 15–32. [Google Scholar] [CrossRef]
- Jenson, J.W.; Keel, T.M.; Mylroie, J.R.; Mylroie, J.E.; Stafford, K.W.; Taborosi, D.; Wexel, C. Karst of the Mariana Islands: The interaction of tectonics, glacioeustasy and freshwater/sea-water mixing in island carbonates. Geol. Soc. Am. Spec. Pap. 2006, 404, 129–138. [Google Scholar]
- Mylroie, J.R.; Mylroie, J.E. Development of the carbonate island karst model. J. Cave Karst Stud. 2007, 69, 59–75. [Google Scholar]
- Arfib, B.; de Marsily, G.; Ganoulis, J. Locating the zone of saline intrusion in a coastal karst aquifer using spring flow data. Ground Water 2007, 45, 28–35. [Google Scholar] [CrossRef]
- Beddows, P.A.; Smart, P.L.; Whitaker, F.F.; Smith, S.L. Decoupled fresh-saline circulation of a coastal carbonate aquifer: Spatial patterns of temperature and specific electrical conductivity. J. Hydrol. 2007, 16, 18–32. [Google Scholar] [CrossRef]
- Back, W.; Hanshaw, B.B.; Herman, J.S.; Van Driel, J.N. Differential dissolution of a Pleistocene reef in the groundwater mixing zone of coastal Yucatan, Mexico. Geology 1986, 14, 137–140. [Google Scholar] [CrossRef]
- Raeisi, E.; Mylroie, J.E. Hydrodynamic behavior of caves formed in the freshwater lens of carbonate islands. Carbonates Evaporites 1995, 10, 207–214. [Google Scholar] [CrossRef]
- Mylroie, J.E.; Carew, J.L. The flank margin model for dissolution cave development in carbonate platforms. Earth Surf. Process. Landf. 1990, 15, 413–424. [Google Scholar] [CrossRef]
- Carew, J.L.; Mylroie, J.E. Quaternary tectonic stability of the Bahamian Archipelago: Evidence from fossil coral reefs and flank margin caves. Quat. Sci. Rev. 1995, 14, 144–153. [Google Scholar] [CrossRef]
- Labourdette, R.; Lascu, I.; Mylroie, J.; Roth, M. Process-like modeling of flank margin caves: From genesis to burial evolution. J. Sediment. Res. 2007, 77, 965–979. [Google Scholar] [CrossRef]
- Waterstrat, W.J.; Mylroie, J.E.; Owen, A.M.; Mylroie, J.R. Coastal caves in Bahamian eolian calcarenites: Differentiating between sea caves and flank margin caves using quantitative morphology. J. Caves Karst Stud. 2010, 72, 61–74. [Google Scholar] [CrossRef]
- Mylroie, J.E.; Carew, J.J. Solution conduits as indicators of late Quaternary Sea level position. Quat. Sci. Rev. 1988, 7, 55–64. [Google Scholar] [CrossRef]
- Dipova, N.; Okudan, E.S. Sea caves, flank margin caves and tufa caves observed on Antalya coastal cliffs. J. Coastal Res. 2011, 61, 382–386. [Google Scholar] [CrossRef]
- Gulley, J.D.; Martin, J.B.; Moore, P.J.; Brown, A.; Spellman, P.D.; Ezell, J. Heterogeneous distributions of CO2 may be more important for dissolution of karstification in coastal eogenetic limestone than mixing dissolution. Earth Surf. Process. Landf. 2015, 40, 1057–1071. [Google Scholar] [CrossRef]
- Gulley, J.D.; Martin, J.B.; Brown, A. Organic carbon inputs, common ions and degassing: Rethinking mixing dissolution in coastal eogenetic carbonate aquifers. Earth Surf. Process. Landf. 2016, 41, 2098–2110. [Google Scholar] [CrossRef]
- Otoničar, B.; Buzjak, N.; Mylroie, J.; Mylroie, J. Flank margin cave development in carbonate talus breccia facies: An example from Cres Island, Croatia. Acta Carsologica 2010, 39, 79–91. [Google Scholar] [CrossRef] [Green Version]
- White, S.; Webb, J.A. The influence of tectonics on flank margin cave formation on a passive continental margin: Naracoorte, Southeastern Australia. Geomorphology 2015, 229, 58–72. [Google Scholar] [CrossRef]
- Suric, M.; Juracic, M.; Horvatincic, N.; Bronic, I.K. Late-Pleistocene–Holocene sea level rise and the pattern of coastal karst inundation: Records from submerged speleothems along the Eastern Adriatic Coast (Croatia). Mar. Geol. 2005, 214, 163–175. [Google Scholar] [CrossRef]
- Mylroie, J.E.; Mylroie, J.R.; Nelson, C.S. Flank margin cave development in telogenetic limestones of New Zealand. Acta Carsologica 2008, 37, 15–40. [Google Scholar] [CrossRef]
- Arriolabengoa, M.; D’Angeli, I.M.; De Waele, J.; Parise, M.; Ruggieri, R.; Sanna, L.; Madonia, G.; Vattano, M. Flank Margin Caves in Telogenetic Limestones in Italy. In Proceedings of the 17th International Congress of Speleology, Sydney, Australia, 23–29 July 2017; Moore, K., White, S., Eds.; Australian Speleological Federation Inc.: Sydney, Australia, 2017; Volume 2, pp. 289–292. [Google Scholar]
- Bonacci, O.; Pipan, T.; Culver, D.C. A framework for karst ecohydrology. Environ. Geol. 2009, 56, 891–900. [Google Scholar] [CrossRef]
- Northup, D.E.; Lavoie, K.H. Geomicrobiology of caves: A review. Geomicrobiol. J. 2001, 18, 199–222. [Google Scholar]
- Barton, H.A.; Northup, D.E. Geomicrobiology in cave environments: Past, current and future perspectives. J. Cave Karst Stud. 2007, 69, 163–178. [Google Scholar]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Viles, H.A. Microbial geomorphology: A neglected link between life and landscape. Geomorphology 2012, 157-158, 6–16. [Google Scholar] [CrossRef]
- Phillips, J.D. Biogeomorphology and contingent ecosystem engineering in karst landscapes. Prog. Phys. Geogr. 2016, 40, 503–526. [Google Scholar] [CrossRef]
- Mejía-Ortíz, L.M.; Pipan, T.; Culver, D.C.; Sprouse, P. The blurred line between photic and aphotic environments: A large Mexican cave with almost no dark zone. Int. J. Speleol. 2018, 47, 69–80. [Google Scholar] [CrossRef]
- D’Angeli, I.M.; Naylor, L.A.; Lee, M.; Miller, A.Z.; Mylroie, J.; De Waele, J. Light attenuation as a control for microbiogeomorphic features: Implications for coastal cave speleogenesis. Geomorphology 2020, 354, 107054. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Bianchi, N. Mediterranean marine caves: A synthesis of current knowledge. In Oceanography and Marine Biology: An Annual Review; Hawkins, S.J., Lemasson, A.J., Allcock, A.L., Bates, A.E., Byrne, M., Evans, A.J., Firth, L.B., Marzinelli, E.M., et al., Eds.; Taylor and Francis: London, UK, 2021; Volume 59, pp. 1–88. [Google Scholar]
- Cavalier-Smith, T. Kingdom Chromista and its eight phyla: A new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma 2018, 255, 297–357. [Google Scholar] [CrossRef] [Green Version]
- Hayward, B.W.; Le Coze, F.; Vachard, D.; Gross, O. World Foraminifera Database. Available online: http://www.marinespecies.org/foraminifera (accessed on 21 September 2021).
- Murray, J.W. Biodiversity of living benthic foraminifera: How many species are there? Mar. Micropal. 2007, 64, 163–176. [Google Scholar] [CrossRef]
- Dias, B.B.; Hart, M.B.; Smart, C.W.; Hall-Spencer, J.M. Modern sea-water acidification: The response of foraminifera to high-CO2 conditions in the Mediterranean Sea. J. Geol. Soc. Lond. 2010, 167, 843–846. [Google Scholar] [CrossRef] [Green Version]
- Alve, E. Colonization of new habitats by benthic foraminifera: A review. Earth Sci. Rev. 1999, 46, 167–185. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Palaeoecology of Benthic Foraminifera; Longman Scientific & Technical: London, UK, 1991; pp. 1–397. [Google Scholar]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: New York, NY, USA, 2006; pp. 1–426. [Google Scholar]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S. Members of the FOBIMO group, 2012. The FOBIMO (Foraminiferal Bio-Monitoring) initiative—Towards a standardized protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropaleontol. 2012, 94-95, 1–13. [Google Scholar] [CrossRef]
- Bergamin, L.; Di Bella, L.; Ferraro, L.; Frezza, V.; Pierfranceschi, G.; Romano, E. Benthic foraminifera in a coastal marine area of the eastern Ligurian Sea (Italy): Response to environmental stress. Ecol. Indic. 2019, 96, 16–31. [Google Scholar] [CrossRef]
- de Nooijer, L.J.; Duijnstee, I.A.P.; van der Zwaan, G.J. Novel application of MTT reduction: A viability assay for temperate shallow-water benthic foraminifera. J. Foraminifer. Res. 2006, 36, 195–200. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Ostermann, D.R.; Williams, D.S.; Blanks, J.K. Comparison of two methods to identify live benthic foraminifera: A test between Rose Bengal and CellTracker Green with implications for stable isotope paleoreconstructions. Paleoceanography 2006, 21, PA4210. [Google Scholar] [CrossRef]
- Scott, D.B.; Medioli, F.S.; Schafer, C.T. Monitoring in Coastal Environments Using Foraminifera and Thecamoebian Indicators; Cambridge University Press: New York, NY, USA, 2001; pp. 1–177. [Google Scholar]
- Dolven, J.K.; Alve, E.; Rygg, B.; Magnusson, J. Defining past ecological status and in situ reference conditions using benthic foraminifera: A case study from the Oslofjord, Norway. Ecol. Indic. 2013, 29, 219–233. [Google Scholar] [CrossRef]
- Avnaim-Katav, S.; Almogi-Labin, A.; Sandler, A.; Sivan, D. Benthic foraminifera as palaeoenvironmental indicators during the last million years in the eastern Mediterranean inner shelf. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 386, 512–530. [Google Scholar] [CrossRef]
- Sousa, S.H.M.; Yamashita, C.; Semensatto, D.L., Jr.; Santarosa, A.C.A.; Iwai, F.S.; Omachi, C.Y.; Disaró, S.T.; Martins, M.V.A.; Barbosa, C.F.; Bonetti, C.H.C.; et al. Members of the BIOFOM group. Opportunities and challenges in incorporating benthic foraminifera in marine and coastal environmental biomonitoring of soft sediments: From science to regulation and practice. J. Sediment. Environ. 2020, 5, 267–275. [Google Scholar] [CrossRef]
- Zeppilli, D.; Leduc, D.; Fontanier, C.; Fontaneto, D.; Fuchs, S.; Gooday, A.J.; Goineau, A.; Ingels, J.; Ivanenko, V.N.; Møbjerg Kristensen, R.; et al. Characteristics of meiofauna in extreme marine ecosystems: A review. Mar. Biodiv. 2018, 48, 35–71. [Google Scholar] [CrossRef] [Green Version]
- Proctor, C.J.; Smart, P.L. A dated cave sediment record of Pleistocene transgressions on Berry Head, Southwest England. J. Quat. Sci. 1991, 6, 233–244. [Google Scholar] [CrossRef]
- Rosso, A.; Sanfilippo, R.; Ruggieri, R.; Maniscalco, R.; Vertino, A. Exceptional record of submarine cave communities from the Pleistocene of Sicily (Italy). Lethaia 2014, 48, 133–144. [Google Scholar] [CrossRef]
- Rosso, A.; Sanfilippo, R.; Vertino, A.; Zibrowius, H. Hanging coral gardens of a Tyrrhenian submarine cave from Sicily (Italy). Boll. Soc. Paleont. It. 2017, 56, 1–12. [Google Scholar]
- Rubin, C.M.; Horton, B.; Sieh, K.; Pilarczyk, J.E.; Daly, P.; Ismail, N.; Parnell, A.C. Highly variable recurrence of tsunamis in the 7400 years before the 2004 Indian Ocean tsunami. Nat. Commun. 2017, 8, 16019. [Google Scholar] [CrossRef] [PubMed]
- Switzer, A.D.; Felix, R.P.; Soria, J.L.A.; Shaw, T.A. A comparative study of the 2013 typhoon Haiyan overwash sediments from a coastal cave and beach system at Salcedo, Eastern Samar, central Philippines. Mar. Geol. 2020, 419, 106083. [Google Scholar] [CrossRef]
- van Hengstum, P.J.; Scott, D.B.; Javaux, E.J. Foraminifera in elevated Bermudian caves provide further evidence for +21 m eustatic sea level during Marine Isotope Stage. Quat. Sci. Rev. 2009, 28, 1850–1860. [Google Scholar] [CrossRef]
- Omori, A.; Kitamura, A.; Fujita, K.; Honda, K.; Yamamoto, N. Reconstruction of light conditions within a submarine cave during the past 7000 years based on the temporal and spatial distribution of algal symbiont-bearing large benthic foraminifers. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 292, 443–452. [Google Scholar] [CrossRef] [Green Version]
- van Hengstum, P.J.; Scott, D.B. Sea-level rise and coastal circulation controlled Holocene groundwater development in Bermuda and caused a meteoric lens to collapse 1600 years ago. Mar Micropal. 2012, 90-91, 29–43. [Google Scholar] [CrossRef]
- van Hengstum, P.J.; Reinhardt, E.G.; Beddows, P.A.; Schwarcz, H.P.; Gabriel, J.J. Foraminifera and testate amoebae (thecamoebians) in an anchialine cave: Surface distributions from Aktun Ha (Carwash) cave system, Mexico. Limnol. Oceanogr. 2009, 54, 391–396. [Google Scholar] [CrossRef]
- Brankovits, D.; Little, S.N.; Winkler, T.S.; Tamalavage, A.E.; Mejía-Ortíz, L.M.; Maupin, C.R.; Yáñez-Mendoza, G.; van Hengstum, P.J. Changes in organic matter deposition can impact benthic marine meiofauna in karst subterranean estuaries. Front. Environ. Sci. 2021, 9, 670914. [Google Scholar] [CrossRef]
- van Hengstum, P.J.; Reinhardt, E.G.; Beddows, P.A.; Huang, R.J.; Gabriel, J.J. Thecamoebians (testate amoebae) and foraminifera from three anchialine cenotes in Mexico: Low salinity (1.5–4.5 psu) faunal transitions. J. Foraminifer. Res. 2008, 38, 305–317. [Google Scholar] [CrossRef]
- Javaux, E.J.; Scott, D.B. Illustration of modern benthic foraminifera from Bermuda and remarks on distribution in other subtropical/tropical Areas. Palaeontologia Electronica 2003, 6, 29. [Google Scholar]
- Little, S.N.; van Hengstum, P.J. Intertidal and subtidal benthic foraminifera in flooded caves: Implications for reconstructing coastal karst aquifers and cave paleoenvironments. Mar Micropal. 2019, 149, 19–34. [Google Scholar] [CrossRef]
- Cresswell, J.N.; van Hengstum, P.J. Habitat partitioning in the marine sector of karst subterranean estuaries and Bermuda’s marine caves: Benthic foraminiferal evidence. Front. Environ. Sci. 2021, 8, 594554. [Google Scholar] [CrossRef]
- Woodward, J.C.; Lewin, J. Karst geomorphology and environmental change. In The Physical Geography of the Mediterranean, 1st ed.; Woodwars, J., Ed.; Oxford University Press: Oxford, UK, 2009; pp. 287–317. [Google Scholar]
- Belmonte, G.; Ingrosso, G.; Poto, M.; Quarta, G.; D’Elia, M.; Onorato, R.; Calcagnile, L. Biogenic stalactites in submarine caves at the Cape of Otranto (SE Italy): Dating and hypothesis on their formation. Marine Ecology 2009, 30, 376–382. [Google Scholar] [CrossRef]
- Guglielmo, R.; Bergamasco, A.; Minutoli, R.; Patti, F.P.; Belmonte, G.; Spanò, N.; Zagami, G.; Bonanzinga, V.; Guglielmo, L.; Granata, A. The Otranto Channel (South Adriatic Sea), a hot-spot area of plankton biodiversity: Pelagic polychaetes. Sci. Rep. 2019, 9, 19490. [Google Scholar] [CrossRef] [PubMed]
- Balduzzi, A.; Cattaneo, R. Estimation par relèvements photographiques de la distribution de Miniacina miniacea (Sarcomastigophora, Foraminiferida) dans une grotte sous-marine du Golfe de Naples. In Rapports et Proces-Verbaux des Reunions; Commission International pour l’Exploration Scientifique de la Mer Méditerranée: Monaco, Spain, 1985; Volume 29, pp. 127–129. [Google Scholar]
- Gerovasileiou, V.; Chintiroglou, C.C.; Vafidis, D.; Koutsoubas, D.; Sini, M.; Dailianis, T.; Issaris, Y.; Akritopoulou, E.; Dimarchopoulou, D.; Voultsiadou, E. Census of biodiversity in marine caves of the eastern Mediterranean Sea. Medit. Mar. Sci. 2015, 16, 245–265. [Google Scholar] [CrossRef] [Green Version]
- Martí, R.; Uriz, M.J.; Ballesteros, E.; Turon, X. Benthic assemblages in two Mediterranean caves: Species diversity and coverage as a function of abiotic parameters and geographic distance. J. Mar. Biol. Ass. UK 2004, 84, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Bussotti, S.; Terlizzi, A.; Fraschetti, S.; Belmonte, G.; Boero, F. Spatial and temporal variability of sessile benthos in shallow Mediterranean marine caves. Mar. Ecol. Prog. Ser. 2006, 325, 109–119. [Google Scholar] [CrossRef]
- Ricci, S.; Cadeddu, B.; Melis, P.; Manconi, R. The Azzurra cave (Capri): First notes on fouling of Nimpaea. Biol. Mar. Mediterr. 2013, 20, 150–151. [Google Scholar]
- Sanfilippo, R.; Rosso, A.; Guido, A.; Mastandrea, A.; Russo, F.; Riding, R.; Taddei Ruggiero, E. Metazoan/microbial biostalactites from present-day submarine caves in the Mediterranean Sea. Mar. Ecol. 2015, 36, 1277–1293. [Google Scholar] [CrossRef]
- Denitto, F.; Terlizzi, A.; Belmonte, G. Settlement and primary succession in a shallow submarine cave: Spatial and temporal benthic assemblage distinctness. Mar. Ecol. 2007, 28, 35–46. [Google Scholar] [CrossRef]
- Rosso, A.; Gerovasileiou, V.; Sanfilippo, R.; Guido, A. bryozoan assemblages from two submarine caves in the Aegean Sea (Eastern Mediterranean). Mar. Biodiv. 2019, 49, 707–726. [Google Scholar] [CrossRef]
- Taddei Ruggiero, E. Foraminifera of bio and thanatocoenoses from the submarine Isca cave, Sorrento peninsula Southern Italy. Mem. Descr. Carta Geol. d’It. 1994, 52, 373–388. [Google Scholar]
- Romano, E.; Bergamin, L.; Di Bella, L.; Frezza, V.; Marassich, A.; Pierfranceschi, G.; Provenzani, C. Benthic foraminifera as proxies of marine influence in the Orosei marine caves (Sardinia, Italy). Aquat. Conserv. 2020, 30, 701–716. [Google Scholar] [CrossRef]
- Scatolini, A. Sardegna, La Grotta del Bel Torrente ha Raggiunto 8.6 km di Sviluppo e Altri 1500m Sono Stati Appena Esplorati. Scintilena. Available online: https://www.scintilena.com/sardegna-la-grotta-del-bel-torrente-ha-raggiunto-8-6-km-di-sviluppo-e-altri-1500m-sono-stati-appena-esplorati/10/29 (accessed on 4 November 2021).
- Bergamin, L.; Marassich, A.; Provenzani, C.; Romano, E. Foraminiferal ecozones in two submarine caves of the Orosei Gulf (Sardinia, Italy). Rend. Lincei Sci. Fis. Nat. It. 2018, 29, 547–557. [Google Scholar] [CrossRef]
- Romano, E.; Bergamin, L.; Pierfranceschi, G.; Provenzani, C.; Marassich, A. The distribution of benthic foraminifera in Bel Torrente submarine cave (Sardinia, Italy) and their environmental significance. Mar. Environ. Res. 2018, 133, 114–127. [Google Scholar] [CrossRef]
- De Waele, J.; Schafheutle, M.; Waelde, T. Speleogenesis of extensive underwater caves along the gulf of Orosei (Central-east Sardinia, Italy). In Proceedings of the 15th International Congress of Speleology, Kerrville, TX, USA, 19–26 July 2009; Volume 1, pp. 469–474. [Google Scholar]
- Sanna, L.; De Waele, J. Karst landscape and caves in the Gulf of Orosei (Central-East Sardinia): A scientific and cultural resource. In Cuevas: Patrimonio, Naturaleza, Cultura y Turismo; Durán, J.J., Carrasco, F., Eds.; Asociación de Cuevas Turísticas Españolas: Madrid, Spain, 2010; pp. 351–362. [Google Scholar]
- Buosi, C.; Armynot du Châtelet, E.; Cherchi, A. Benthic foraminiferal assemblages in the current-dominated strait of Bonifacio (Mediterranean Sea). J. Foraminifer. Res. 2012, 42, 39–55. [Google Scholar] [CrossRef]
- Buosi, C.; Cherchi, A.; Ibba, A.; Marras, B.; Marrucci, A.; Schintu, M. Benthic foraminiferal assemblages and sedimentological characterization of the coastal system of the Cagliari area (southern Sardinia, Italy). Boll. Soc. Paleontol. Ital. 2013, 52, 1–9. [Google Scholar]
- Buosi, C.; Cherchi, A.; Ibba, A.; Marras, B.; Marrucci, A.; Schintu, M. Preliminary data on benthic foraminiferal assemblages and sedimentological characterization from some polluted and unpolluted coastal areas of Sardinia (Italy). Boll. Soc. Paleontol. Ital. 2013, 52, 35–44. [Google Scholar]
- Cherchi, A.; Da Pelo, S.; Ibba, A.; Mana, D.; Buosi, C.; Floris, N. Benthic foraminifera response and geochemical characterization of the coastal environment surrounding the polluted industrial area of Portovesme (South-Western Sardinia, Italy). Mar. Pollut. Bull. 2009, 59, 281–296. [Google Scholar] [CrossRef]
- Romano, E.; Bergamin, L.; Di Bella, L.; Frezza, V.; Pierfranceschi, G.; Marassich, A.; Provenzani, C. Benthic foraminifera as environmental indicators in extreme environments: The marine cave of Bue Marino (Sardinia, Italy). Ecol. Indic. 2021, 120, 106977. [Google Scholar] [CrossRef]
- Bergamin, L.; Taddei Ruggiero, E.; Pierfranceschi, G.; Andres, B.; Constantino, R.; Crovato, C.; D’Ambrosi, A.; Marassich, A.; Romano, E. Benthic foraminifera and brachiopods from a marine cave in Spain: Environmental significance. Medit. Mar. Sci. 2020, 21, 506–518. [Google Scholar] [CrossRef]
- Gázquez, F.; Calaforra, J.M.; Rodríguez-Estrella, T.; Ros, A.; Llamusí, J.L.; Sánchez, J. Evidence for regional hypogene speleogenesis in Murcia (SE Spain). In Hypogene Karst Regions and Caves of the World, Cave and Karst Systems of the World; Klimchouk, A., Palmer, A.N., Waele, J.D., Auler, A.S., Audra, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 85–97. [Google Scholar]
- Gázquez, F.; Calaforra, J.M.; Ros, A.; Llamusí, J.L.; Sánchez, J. Hypogenic morphologies and speleothems in caves in the Murcia region, south-eastern Spain. Deepkarst 2016, 1–8. [Google Scholar]
- Rosso, A.; Sanfilippo, R.; Taddei Ruggiero, E.; Di Martino, E. Faunas and ecological groups of Serpuloidea, Bryozoa and Brachiopoda from submarine caves in Sicily (Mediterranean Sea). Boll. Soc. Paleontol. Ital. 2013, 52, 167–176. [Google Scholar]
- McGann, M. Potential use of the benthic foraminifers Bulimina denudata and Eggerelloides advenus in marine sediment toxicity testing. Water 2021, 13, 775. [Google Scholar] [CrossRef]
- Uthicke, S.; Momigliano, P.; Fabricius, K.E. High risk of extinction of benthic foraminifera in this century due to ocean acidification. Sci. Rep. 2013, 3, 1769. [Google Scholar] [CrossRef] [Green Version]
- Alve, E.; Goldstein, S.T. Propagule transport as a key method of dispersal in benthic foraminifera (Protista). Limnol. Oceanogr. 2003, 48, 2163–2170. [Google Scholar] [CrossRef] [Green Version]
- Alve, E.; Goldstein, S.T. Dispersal, survival and delayed growth of benthic foraminiferal propagules. J. Sea Res. 2010, 63, 36–51. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bel Torrente | Bue Marino | CT12 | |
|---|---|---|---|
| General features | |||
| Cave origin and characters | karst; Anchialine cave, with development strongly favored by hyperkarst processes (freshwater-saltwater mixing) | karst; Anchialine cave, with development strongly favored by hyperkarst processes (freshwater-saltwater mixing); three main branches | hypogenic origin; submarine cave |
| Sediments | coarse grain size, terrigenous, terrestrial origin | coarse grain size, terrigenous, terrestrial origin; abundant vegetal debris in the Middle Branch | fine grain size, authigenic micrite, bioclasts |
| Foraminiferal ecozones | |||
| Marine ecozone | low-mild absolute abundance, high diversity; P. pertusus, E. crispum, L. lobatula, G. praegeri | low absolute abundance, mild diversity; P. pertusus, G. praegeri, E. crispum, L. lobatula | high absolute abundance, mild diversity; M. subrotunda, L. lobatula, G. praegeri |
| Entrance Ecozone | low absolute abundance, mild diversity; prevalence of calcareous taxa (G. praegeri, Bolivina spp., Quinqueloculina spp.) over the agglutinated (R. dentaliniformis) | low-mild absolute abundance, mild-high diversity; prevalence of calcareous taxa (G. praegeri and R. bradyi) over the agglutinated (R. dentaliniformis) | high absolute abundance, low-mild diversity; P. corrugata and S. vivipara with similar percentages, T. inornata. Increase of epifaunal clinging/attached taxa |
| Transitional ecozone | low absolute abundance, low-mild diversity; prevalence of agglutinated taxa (E. advenus, R. dentaliniformis, C. jeffreysii) over the calcareous ones (G. praegeri, R. brady, A. inflata) | low absolute abundance, mild diversity; prevalence of agglutinated taxa (E. advenus, L. ochracea, R. dentaliniformis, C. jeffreysii) over the calcareous ones (G. praegeri, R. brady) | high absolute abundance, low-mild diversity; dominant P. corrugata. Further increase of epifaunal clinging/attached taxa |
| Confluence ecozone | not present | high absolute abundance, low-mild diversity; dominant opportunistic agglutinated taxa (N. polymorphinoides in spring, E. advenus in summer) | not present |
| Beginning of the barren zone | at about 330 m from the entrance | at about 450 m from the entrance | barren zone not present within the 120 m length of the cave |
| Driving environmental factors of foraminiferal zonation | salinity and pH gradient, environmental instability due to seasonal freshwater flows | salinity and pH gradient, environmental instability due to seasonal freshwater flows; organic matter in the Middle Branch | sediment grain size, organic matter |
| Species | Isca Cave (Tyrrhenian Coast) | Gulf of Orosei (Sardinia, Italy) | CT12 Murcia Region (Spain) | Other Mediterranean Caves | Yucatan (Mexico) | Bermuda | Other Caves Outside the Mediterranean |
|---|---|---|---|---|---|---|---|
| Adelosina spp. | X | ||||||
| Ammodiscus tenuis (Brady, 1881) | X | ||||||
| Ammonia beccarii (Linnaeus, 1758) | X | ||||||
| Ammonia inflata (Seguenza, 1862) | X | ||||||
| Ammonia tepida (Cushman, 1926) | X | ||||||
| Ammonia spp. | X | ||||||
| Amphistegina radiata (Fichtel & Moll, 1798) | X | ||||||
| Baculogipsina sphaerulata (Parker & Jones, 1860) | X | ||||||
| Calcarina spp. | X | ||||||
| Bolivina spp. | X | X | |||||
| Bolivina variabilis (Williamson, 1858) | X | X | X | ||||
| Cibicides refulgens Montfort, 1808 | X | ||||||
| Cornuspiramia adherens (Le Calvez, 1935) | X | ||||||
| Cribrostomoides jeffreysii (Williamson, 1858) | X | ||||||
| Eggerelloides advenus (Cushman, 1922) | X | ||||||
| Elphidium crispum (Linnaeus, 1758) | X | ||||||
| Elphidium spp. | X | X | |||||
| Gavelinopsis praegeri (Heron-Allen & Earland, 1913) | X | X | |||||
| Globigerina sp. | X | ||||||
| Jadammina macrescens (Brady, 1870) accepted as Entzia macrescens (Brady, 1870) | X | ||||||
| Lepidodeuterammina ochracea (Williamson, 1858) | X | ||||||
| Lobatula lobatula (Walker & Jacob, 1798) synonim of Cibicidoides lobatulus (Walker & Jacob, 1798) | X | X | |||||
| Melonis barleeanus (Williamson, 1858) accepted as Melonis affinis (Reuss, 1851) | X | X | |||||
| Miliolinella subrotunda (Montagu, 1803) | X | ||||||
| Miniacina miniacea (Pallas, 1766) | X | X | |||||
| Nouria polymorphinoides Heron-Allen & Earland, 1914 | X | ||||||
| Patellina corrugata Williamson, 1858 | X | ||||||
| Peneroplis planatus (Fichtel & Moll, 1798) | X | X | |||||
| Peneroplis pertusus (Forsskål in Niebuhr, 1775) | X | ||||||
| Planogypsina acervalis (Brady, 1884) | X | ||||||
| Polysaccammina ipohalina Scott, 1976 | X | ||||||
| Pseudothurammina limnetis (Scott & Medioli, 1980) | X | ||||||
| Quinqueloculina spp. | X | ||||||
| Reophax dentaliniformis (Brady, 1881) accepted as Nodulina dentaliniformis (Brady, 1881) | X | ||||||
| Rhizammina sp. | X | ||||||
| Rosalina bradyi (Cushman, 1915) | X | ||||||
| Rosalina spp. | X | X | |||||
| Sigmoilina tenuis (Cžjžek, 1848) accepted as Spirosigmoilina tenuis (Cžjžek, 1848) | X | ||||||
| Sejunctella sp. | X | ||||||
| Spirillina vivipara Ehrenberg, 1843 | X | X | |||||
| Spirophthalmidium emaciatum (Haynes, 1973) | X | X | |||||
| Textularia bocky | X | ||||||
| Textularia spp. | X | ||||||
| Trochammina inflata (Montagu, 1808) | X | ||||||
| Tubinella inornata (Brady, 1884) | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, E.; Bergamin, L.; Parise, M. Benthic Foraminifera as Environmental Indicators in Mediterranean Marine Caves: A Review. Geosciences 2022, 12, 42. https://doi.org/10.3390/geosciences12010042
Romano E, Bergamin L, Parise M. Benthic Foraminifera as Environmental Indicators in Mediterranean Marine Caves: A Review. Geosciences. 2022; 12(1):42. https://doi.org/10.3390/geosciences12010042
Chicago/Turabian StyleRomano, Elena, Luisa Bergamin, and Mario Parise. 2022. "Benthic Foraminifera as Environmental Indicators in Mediterranean Marine Caves: A Review" Geosciences 12, no. 1: 42. https://doi.org/10.3390/geosciences12010042
APA StyleRomano, E., Bergamin, L., & Parise, M. (2022). Benthic Foraminifera as Environmental Indicators in Mediterranean Marine Caves: A Review. Geosciences, 12(1), 42. https://doi.org/10.3390/geosciences12010042