
Variety and Distribution of Diatom-Based Sea Ice Proxies in Antarctic Marine Sediments of the Past 2000 Years


Abstract
:1. Introduction
- −
- In the first section, we collate all the published marine sediment records spanning all, or part, of the past 2000 years with sea ice reconstructions derived from diatom-based proxies. We review the composition and variety of proxies used to reconstruct sea ice and map their distribution.
- −
- We then consider the advantages and limitations of the proxies and discuss the potential for improving consistency/standardization between records and the scope for regional syntheses.
2. Results
2.1. Proxy Records
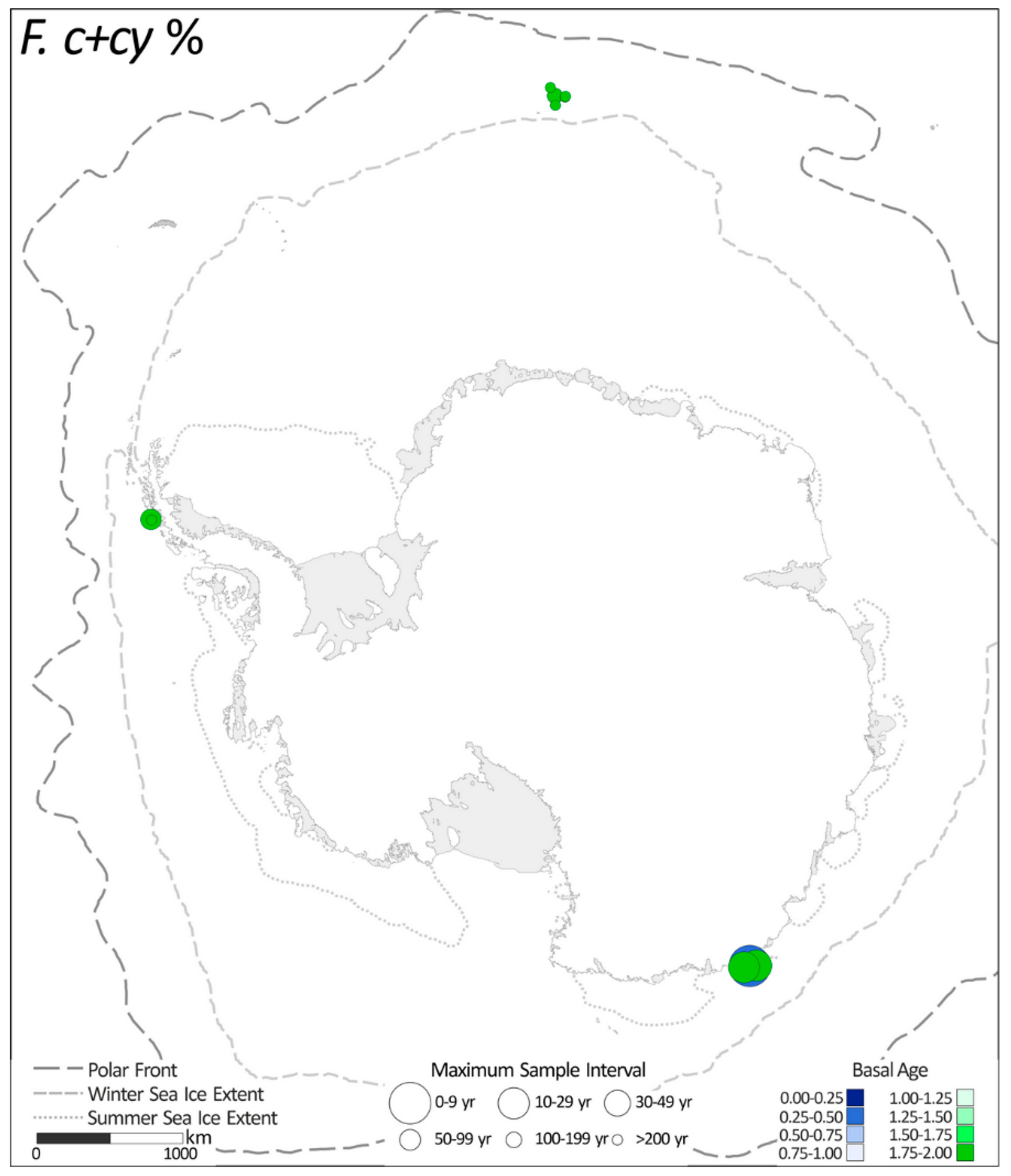
2.1.1. Fragilariopsis curta + F. cylindrus %
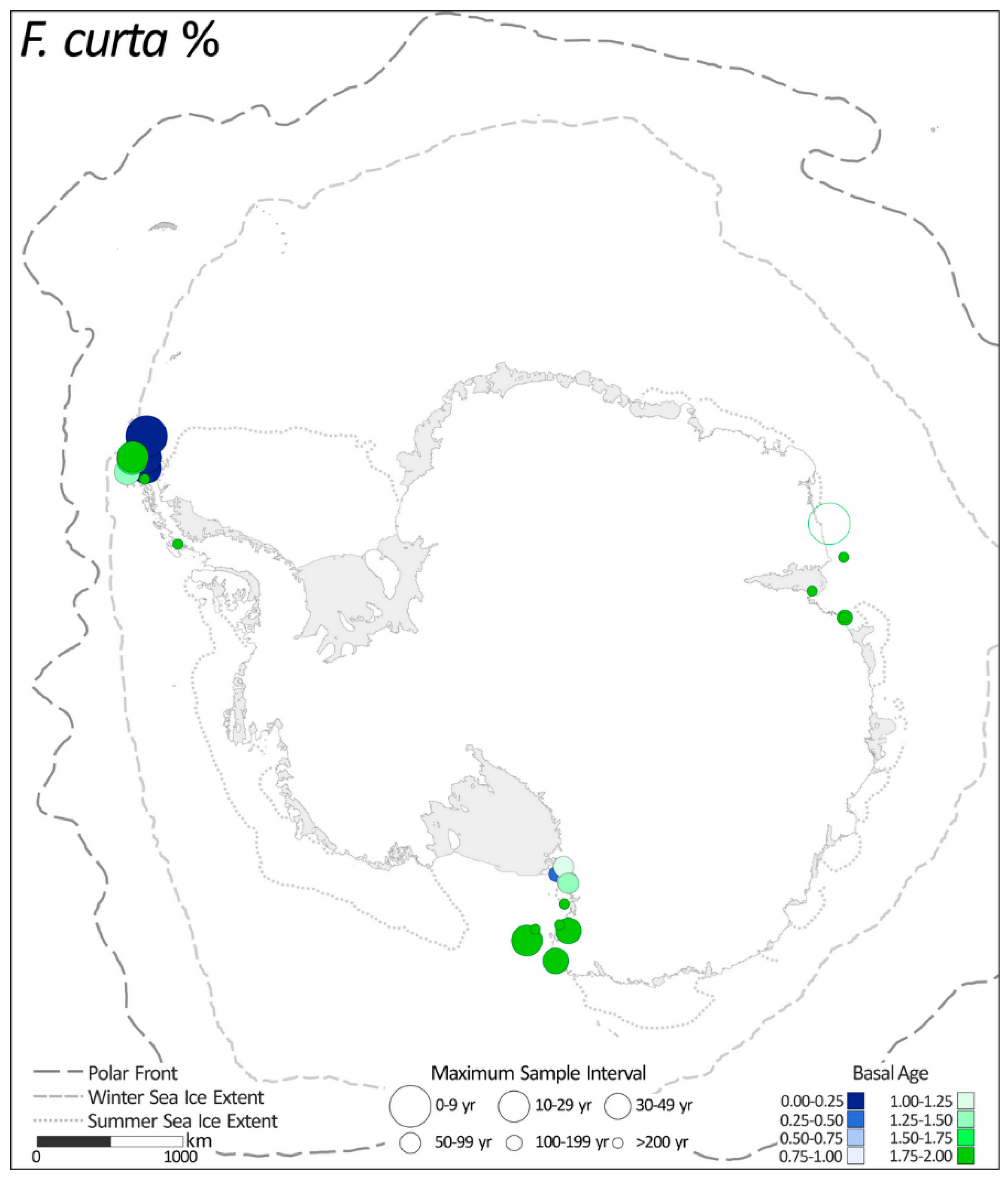
2.1.2. Fragilariopsis curta %
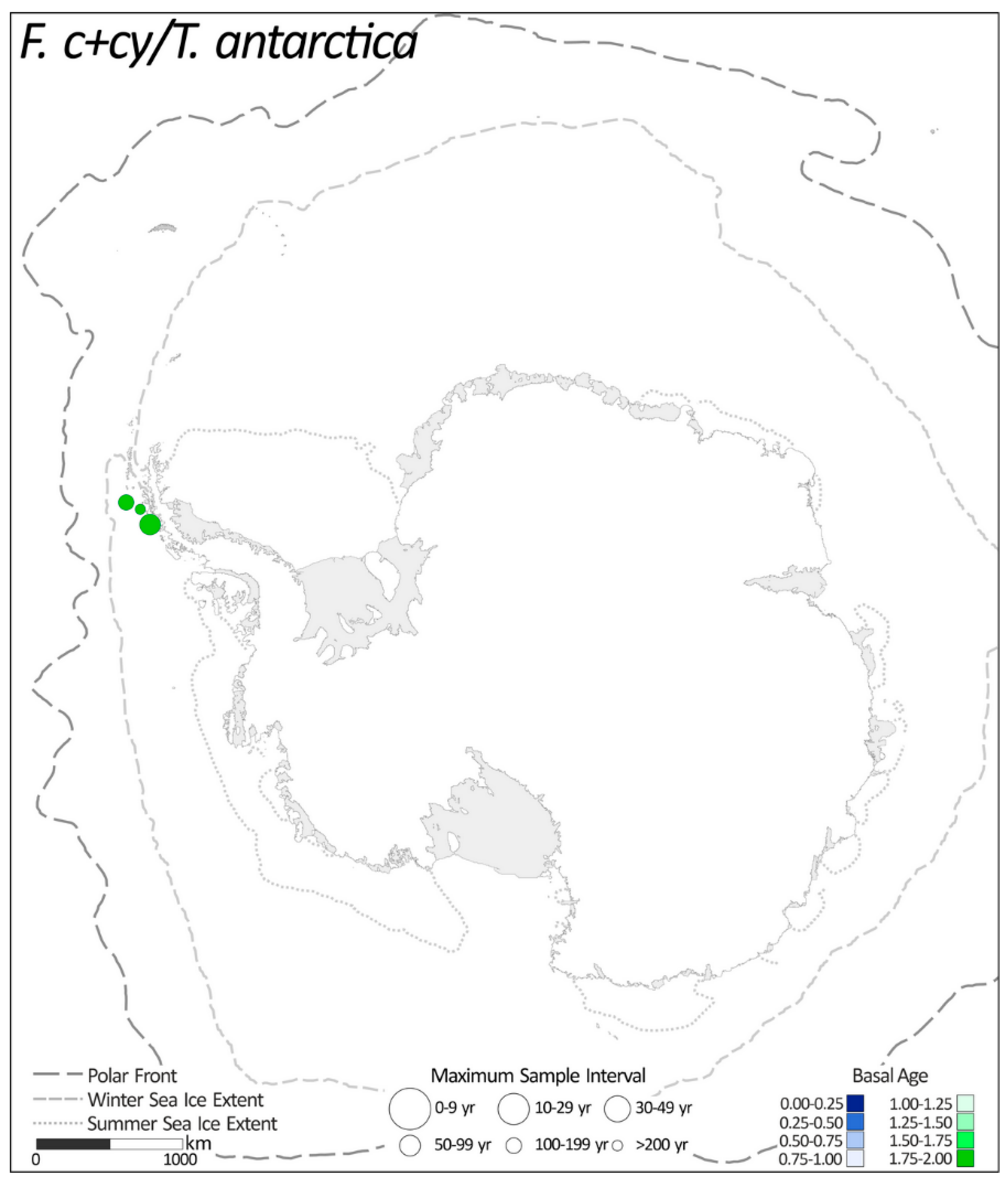
2.1.3. Fragilariopsis curta + F. cylindrus/T. antarctica
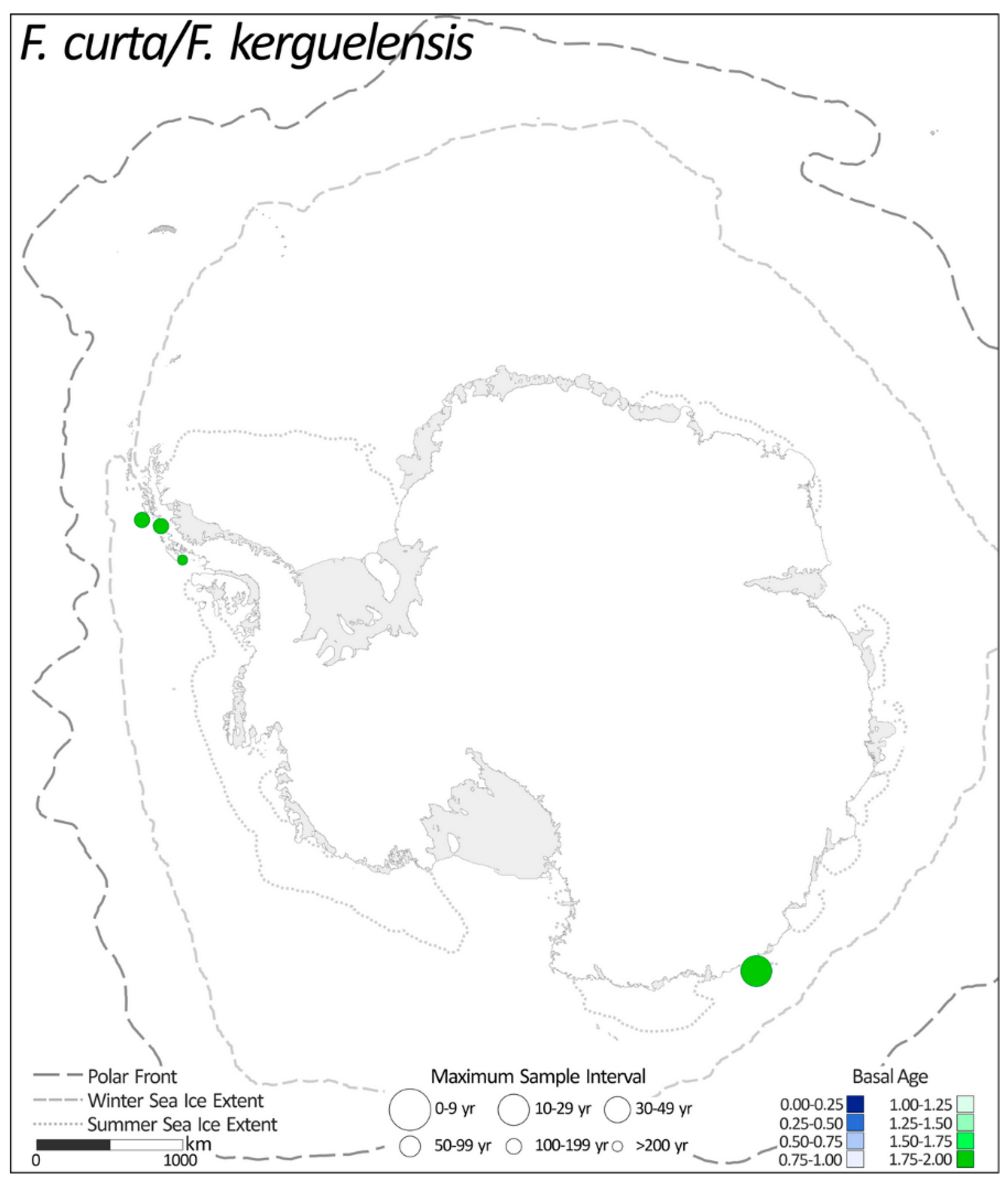
2.1.4. Fragilariopsis curta/Fragilariopsis kerguelensis
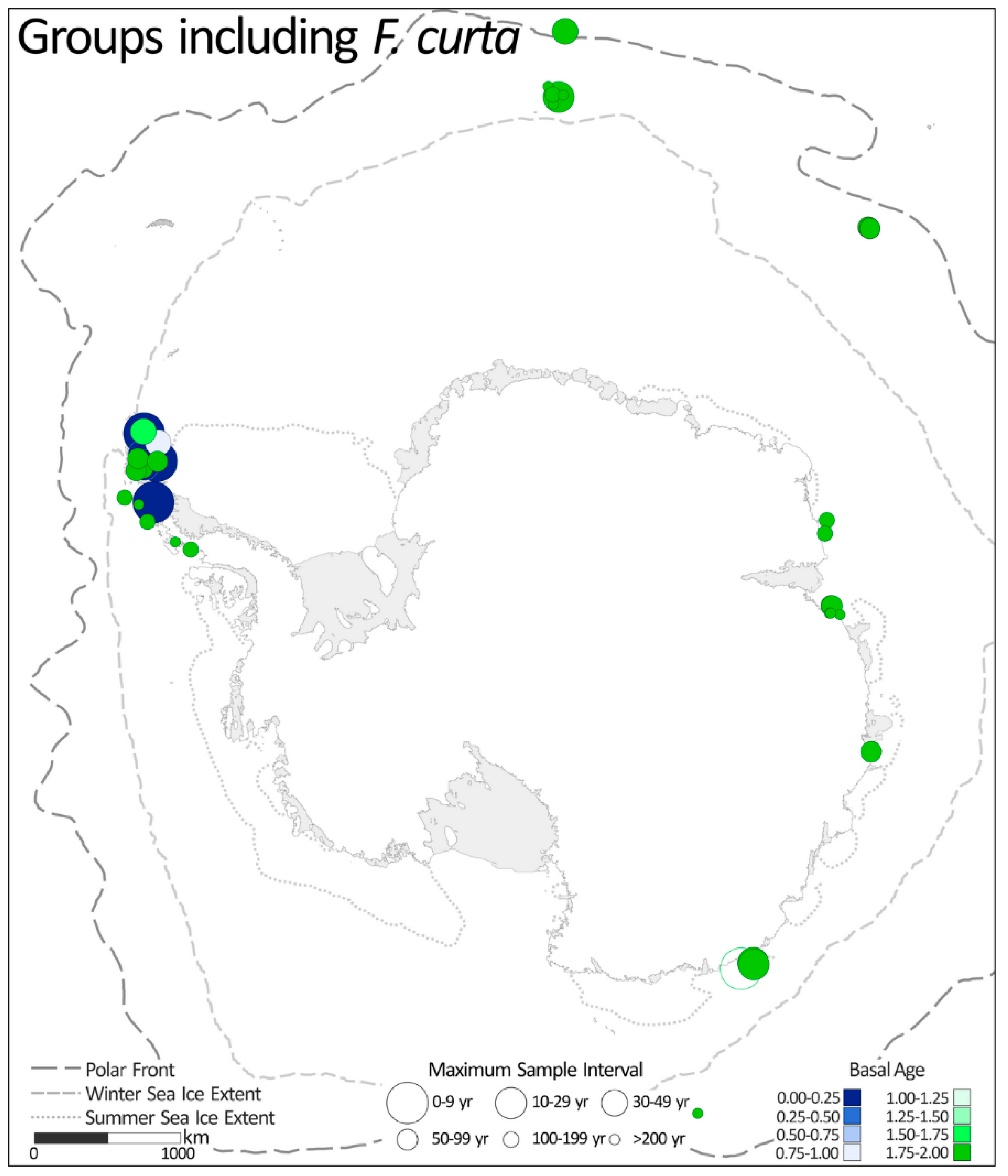
2.1.5. Groups including Fragilariopsis curta
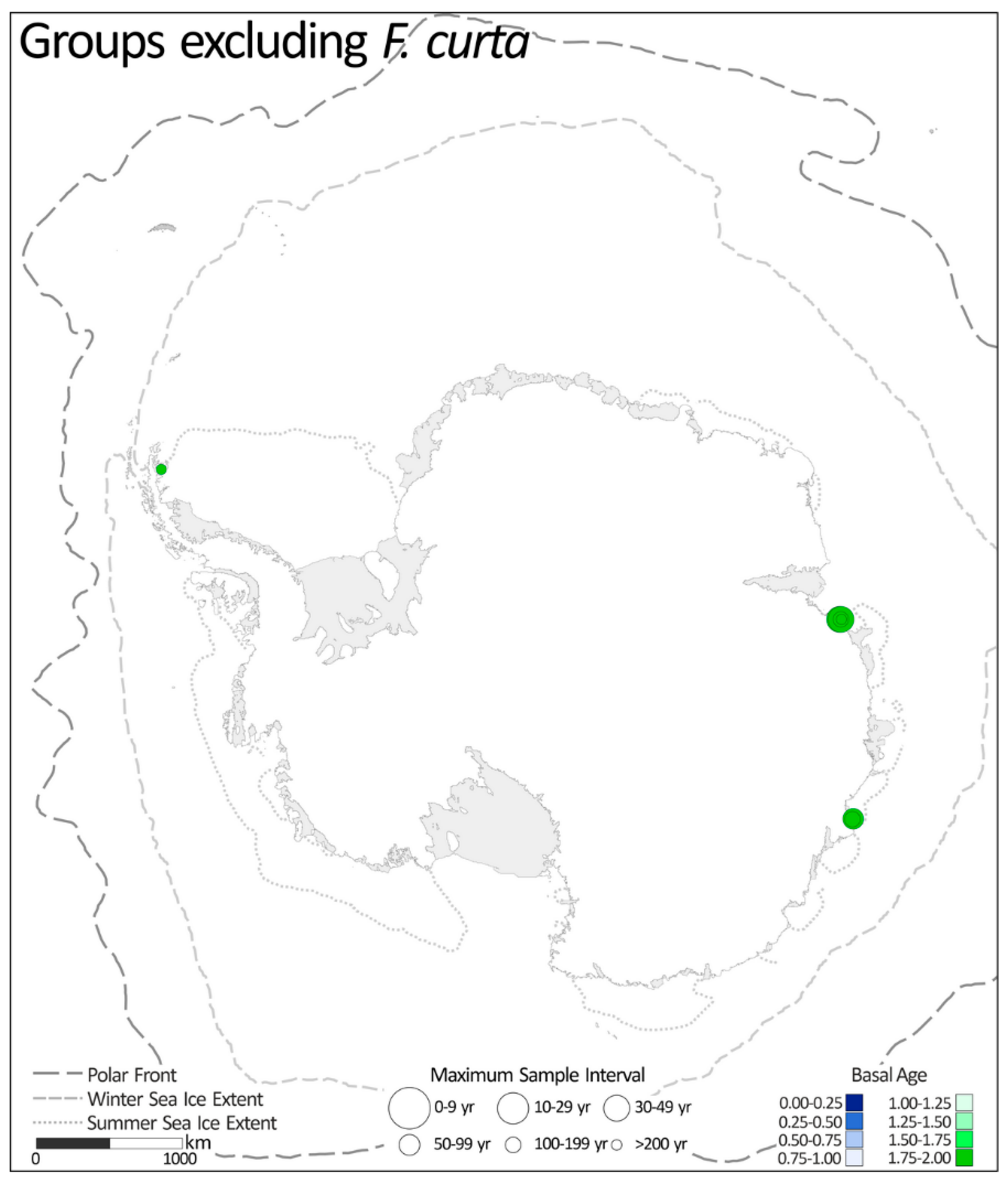
2.1.6. Groups Excluding Fragilariopsis curta
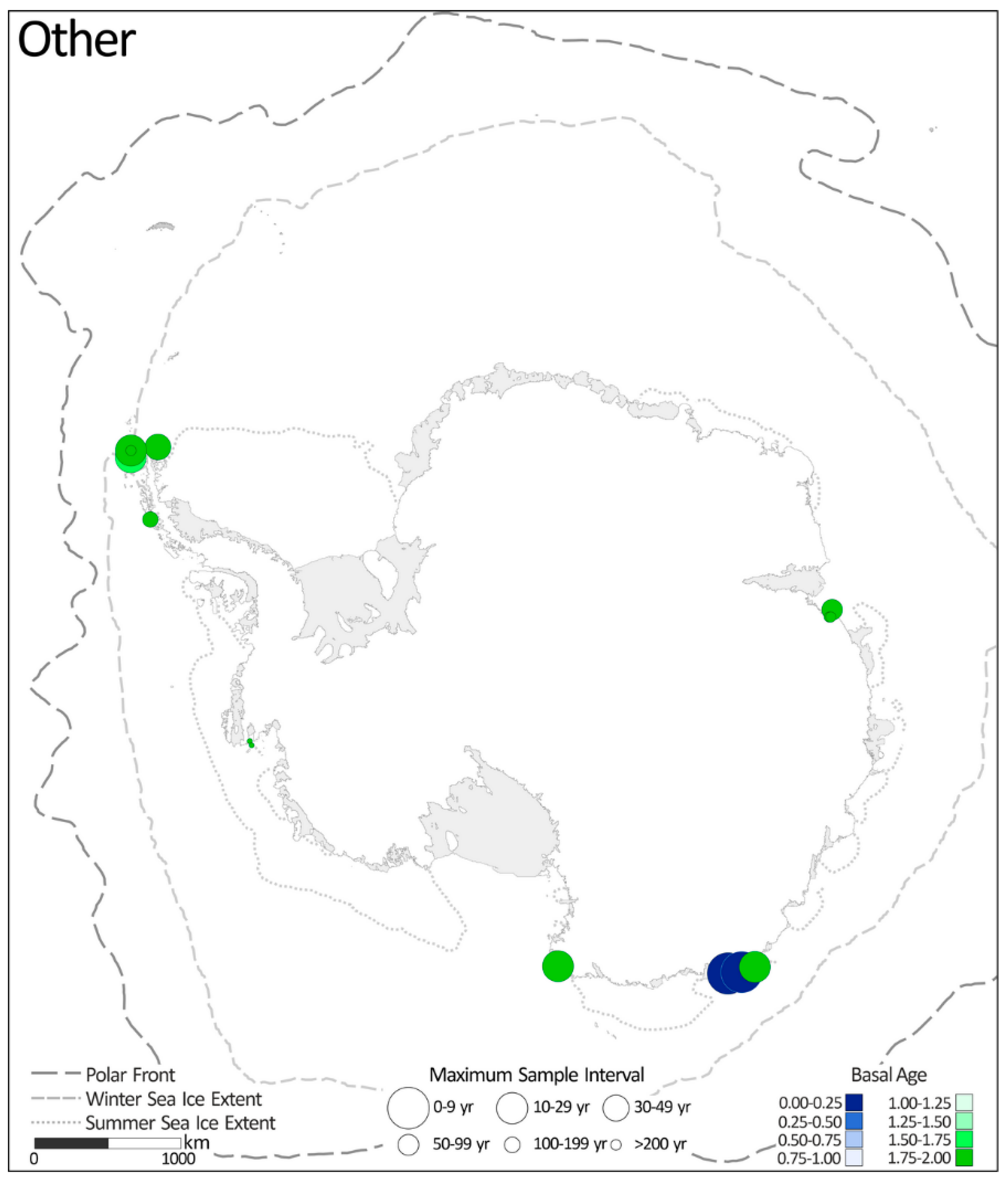
2.1.7. Other
- −
- Entomoneis kjellmannii % (previously Amphiprora kjellmannii) is primarily used as an indicator of perennial sea ice, especially fast ice [17,22,23]. E. kjellmannii is reported as a common, sometimes dominant, member of the sea ice diatom community in coastal areas of the Ross Sea and is also abundant in the sea ice and sea ice adjacent waters of the Ingrid Christensen coast and in parts of the Victoria Land coast [74,75,76]. E. kjellmannii is a component of the ‘fast ice index’ (one of the non-F. curta groups—Section 2.1.6 and Table 6) presented by McMinn [22] and McMinn et al. [23] and used by Berg et al. [17] as evidence for the presence of fast ice on the Rauer Group coast, Prydz Bay.
- −
- Eucampia index is introduced by Kaczmarska et al. [77] and based on the distinct morphology of the ‘pointed’ or ‘horned’ terminal valves and ‘flat’ intercalary valves of both the polar E. antarctica var. recta and the sub-polar E. antarctic var. antarctica morphotypes [78]. The Eucampia index refers to the ratio of terminal to intercalary (‘pointy to flat’) valves. E. antarctica colonies growing in colder waters with greater sea ice cover are characterized by shorter chains with a higher relative proportion of terminal valves [79,80]. Milliken et al. [57] use the Eucampia index to infer changes in sea ice cover over Maxwell Bay in the South Shetland Islands (Site 51). Resolution of this record ranges between 475 and 950 years and spans between 2.0 and 0.1 ka BP (Figure 8 and Table 4).
- −
- Fragilariopsis cylindrus % is used as the primary indicator of prolonged sea ice cover (>8.5 months) by Campagne et al. [30,33], owing to F. cylindrus’s high statistical significance in explaining variability in the diatom assemblages of the past 40 years in the Adélie Margin (Sites 29 and 33) where F. curta was not significant. Yoon et al. [58,60] present separate down-core abundance plots for F. curta and F. cylindrus% (Sites 52 and 54), showing that peaks are often off-set and highlighting the sensitive habitat preferences of the two species.
- −
- The Fragilariopsis group/Thalassiosira antarctica (T2) is similar to the more commonly applied F. curta + F. cylindrus/T. antarctica ratio. The Fragilariopsis group/T. antarctica (T2) ratio also aims to characterize the major shifts in surface ocean conditions with elevated (reduced) contributions of the Fragilariopsis group (T. antarctica) indicative of more persistent (ephemeral) seasonal sea ice. As presented in Kamanidou et al. [55], the Fragilariopsis group comprises of the combined abundance of F. curta, F. cylindrus, F. sublinearis, and F. vanheurckii. Applied to the sediment record from the Perseverance Drift north of Joinville Island in the northern-most Antarctic Peninsula (Site 47), the record covers from 0.8 to 0.0 ka BP at a resolution between 40 and 21 years [55].
- −
- Porosira glacialis/Thalassiosira antarctica are both common components of the diatom assemblage in Antarctic continental shelf sediments with P. glacialis preferring slightly cooler ocean climate conditions than T. antarctica [81,82]. As established by Pike et al. [20], the ratio of P. glacialis/T. antarctica reflects the subtle difference in environmental preferences of the two species, with ratios of >0.1 indicating annual sea ice cover greater than 7.5 months/year [20]. Applied to sediment records from the Svenner Channel, Prydz Bay, and the Dumont d’Urville Trough (Sites 16 and 35), the P. glacialis/T. antarctica ratio is used to reconstruct changes in the duration of sea ice cover [20]. The record from the Svenner Channel (Site 16) covers between 2.0 and 0.6 ka BP with a maximum and minimum resolution of 36 and 70 years, respectively [20]. The Dumont d’Urville record (Site 35) spans the period from 2.0 to 1.0 ka BP with sample intervals between 25 and 2 years [20].
- −
- Pennate–centric ratio—in sediments with low diatom concentrations, Minzoni et al. [15] use the prevalence of pennate diatoms in the assemblage to infer the presence of sea ice. The association between elevated contributions of pennate diatoms and heavier sea ice cover is based on analyses of the core top assemblages that are dominated by F. curta [83]. Applied to cores recovered from Ferrero Bay in the Amundsen Sea Embayment (Sites 10 and 11) where sedimentation rates are exceptionally low for the AP, the records of the pennate–centric ratio cover the whole of the last 2 ka at both sites with sample intervals of 500 to 1000 years (Figure 8 and Table 4).
- −
- Diatom concentrations—Sjunneskog and Taylor [42] and Michalchuk et al. [52] use diatom concentrations as a paleoproductivity proxy for the Palmer Deep and Firth of Tay marine cores (Sites 37 and 45) based on the dominant contribution of the sea ice melt bloom to the total diatom production which is reflected in the prevalence of Chaetoceros resting spores (60 to 90% of the total diatom content) in AP sediments [45,84].
- −
- Dark–light laminae—in laminated sediments from Edisto Inlet in the Ross Sea (Site 64) Tesi et al., [66] show that dark and light layers are characterized by distinctive diatom assemblages, biomarkers, and isotopic values that reflect the two dominant seasonal sea ice settings of the site: (1) sea ice break-up and thaw (dark) and (2) open surface waters (light). Tesi, et al.’s [66] record spans the last 2 ka with sample intervals of <20 years (Figure 8 and Table 4).
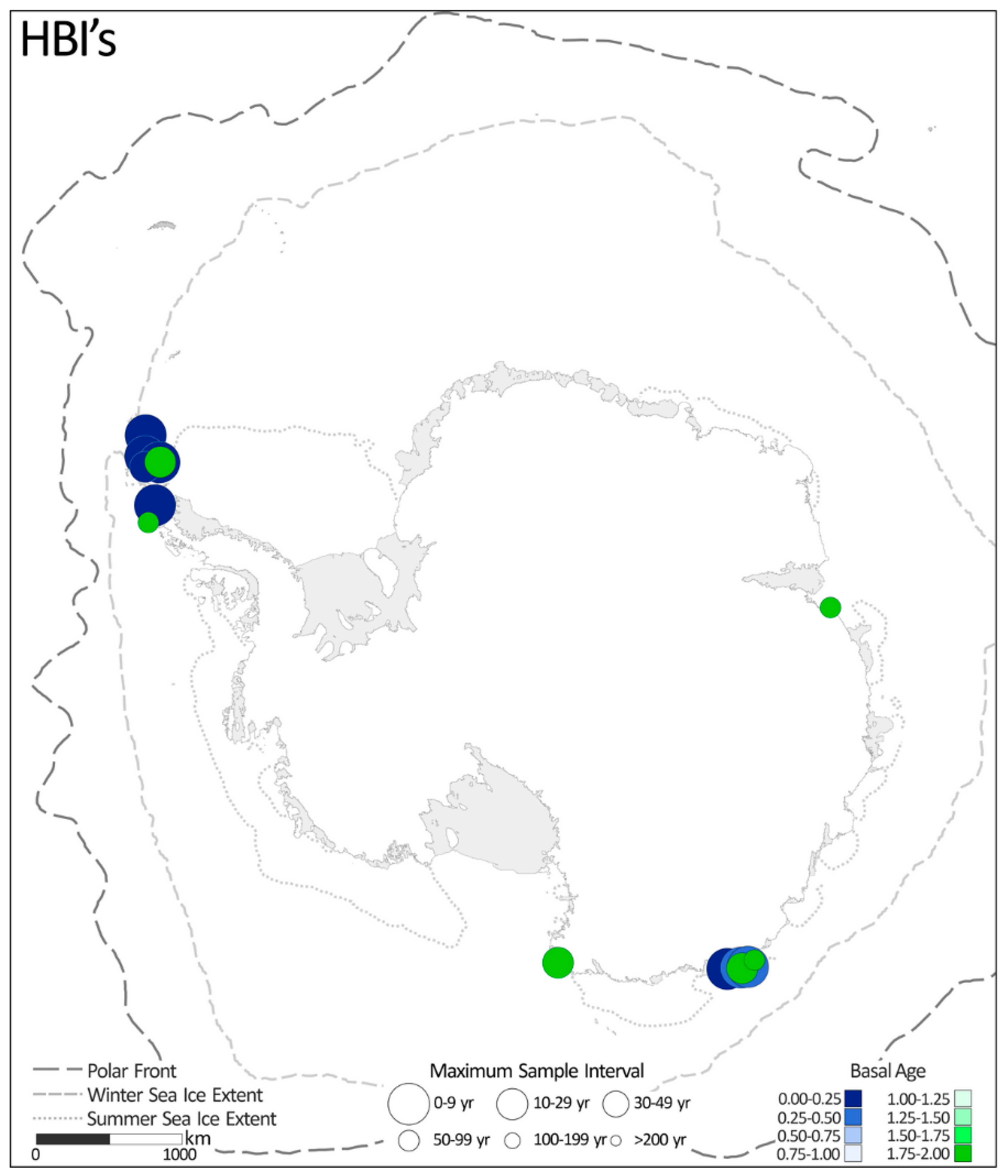
2.1.8. Diatom-Specific Highly Branched Isoprenoids (HBIs)
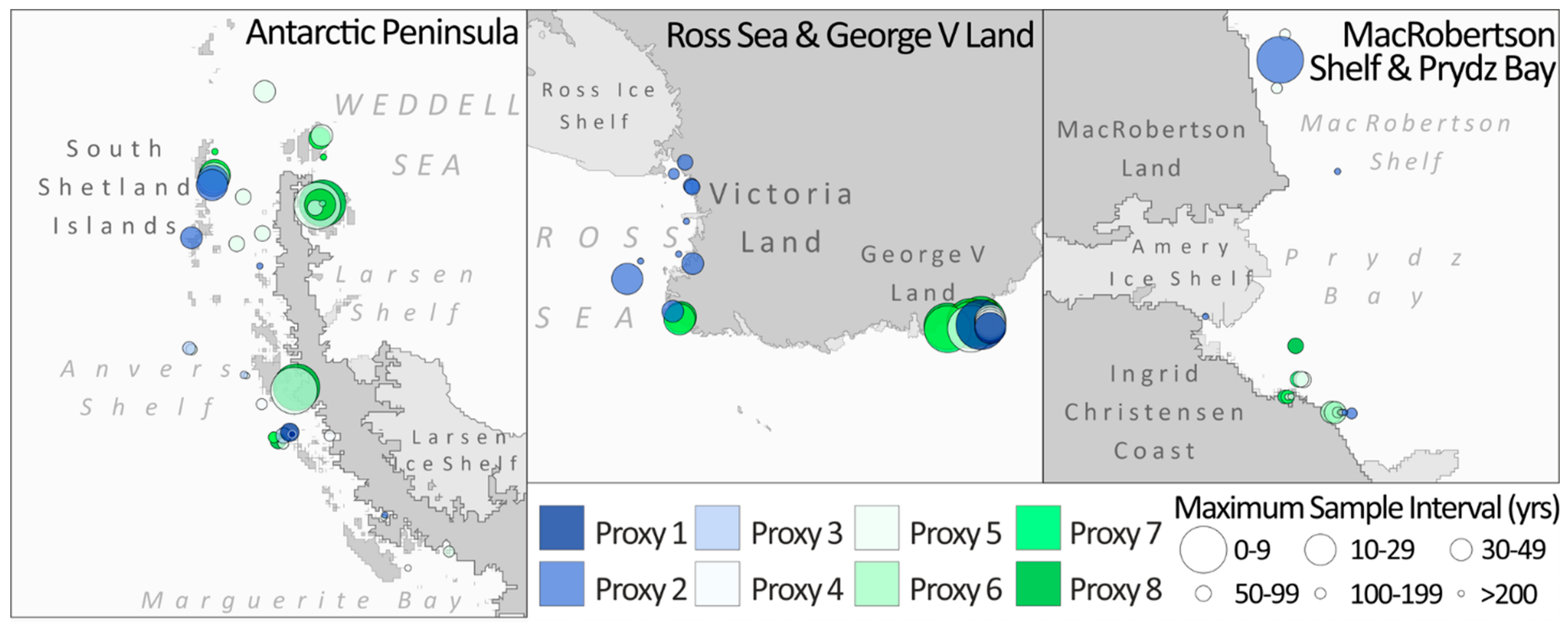
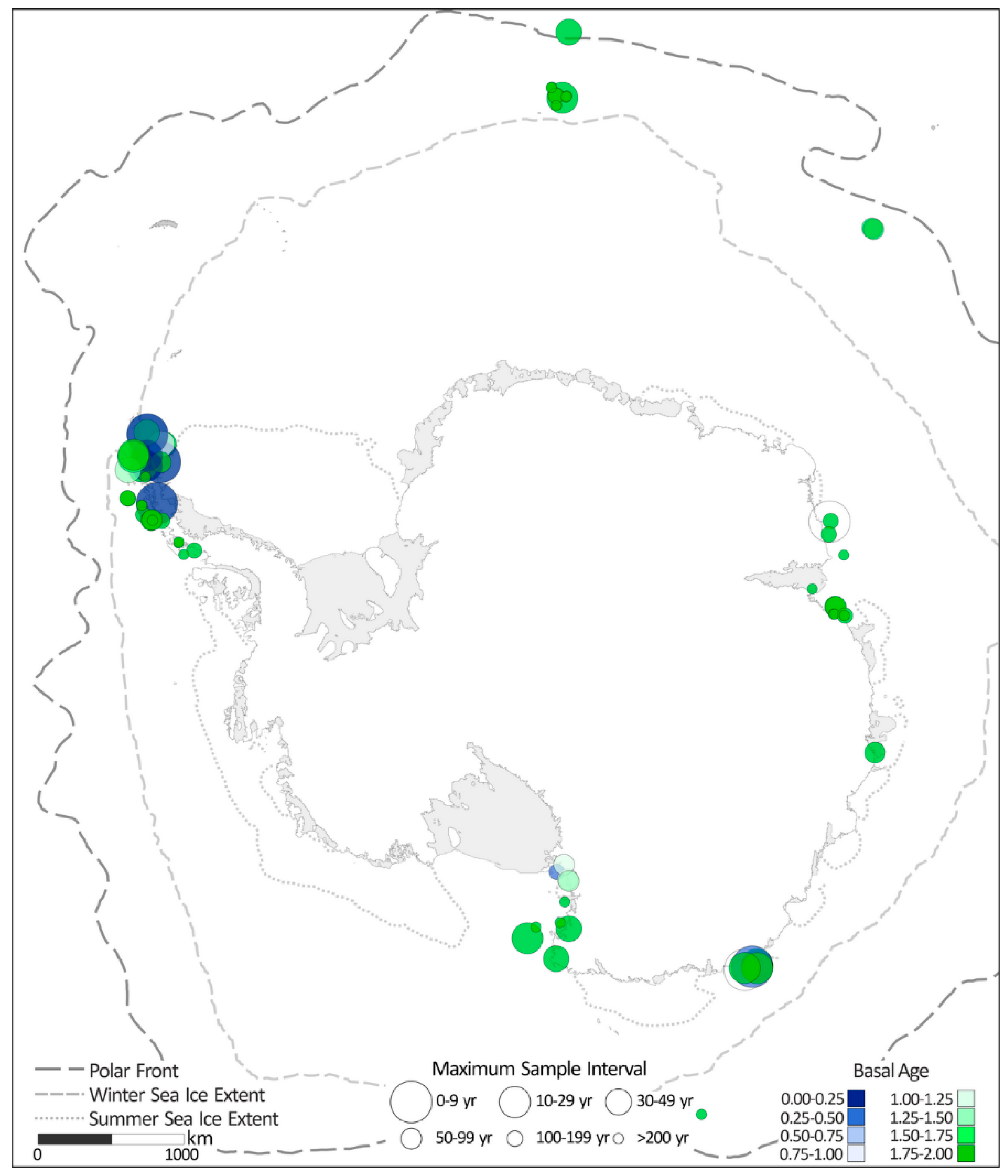
2.2. Distribution of Diatom-Based Proxy Records
3. Advantages and Limitations
3.1. Relative Abundance
3.2. Statistical Approaches
4. Discussion
4.1. Distribution
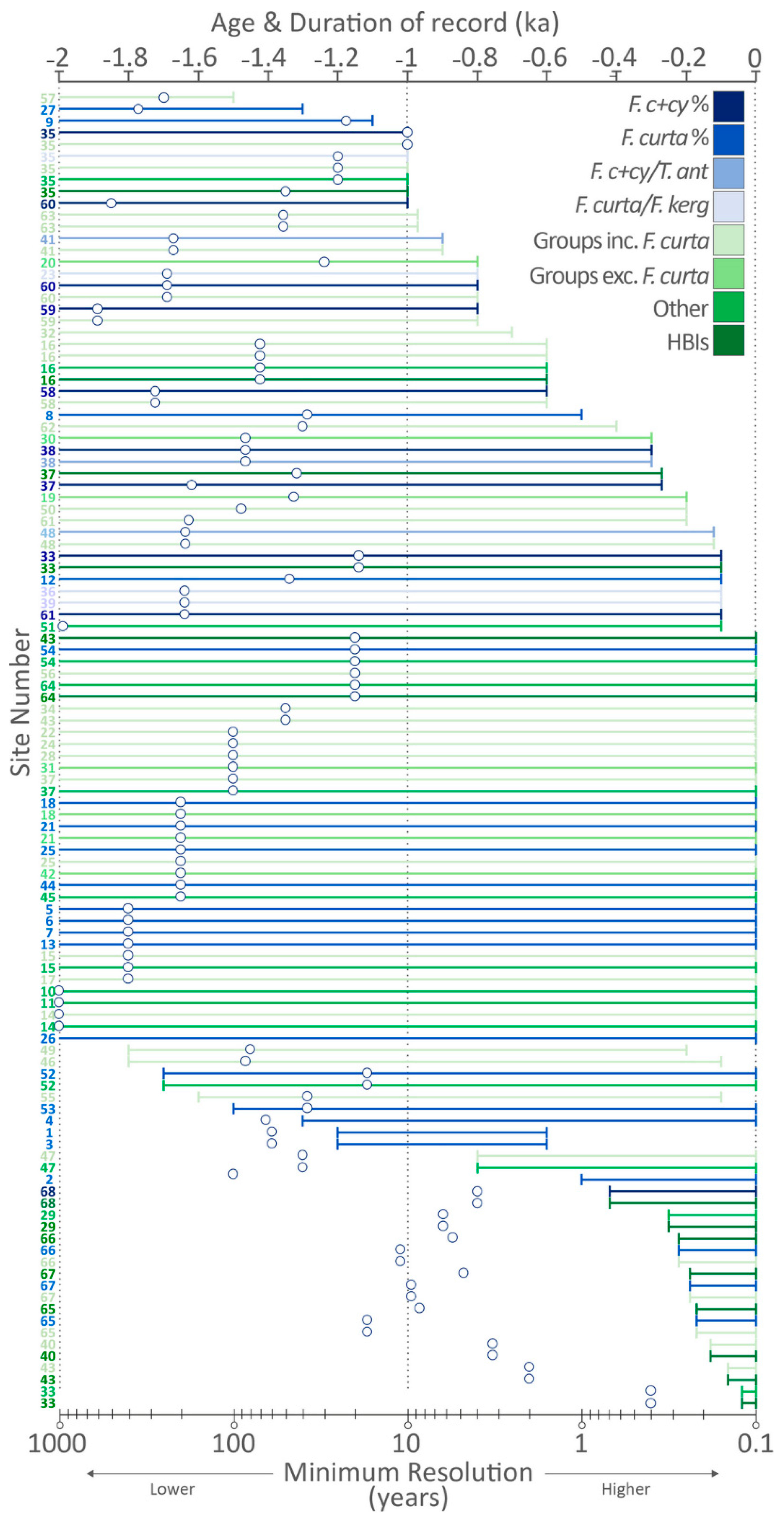
4.2. Age, Duration and Resolution of Records
4.3. Scope for Validation and Standardisation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Fetterer, F.; Knowles, K.; Meier, W.N.; Savoie, M.; Windnagel, A.K. Sea Ice Index, Version 3 (Southern Hemisphere); National Snow and Ice Data Center: Boulder, CO, USA, 2017. [Google Scholar]
- Stammerjohn, S.; Massom, R.; Rind, D.; Martinson, D. Regions of rapid sea ice change: An inter-hemispheric seasonal comparison. Geophys. Res. Lett. 2012, 39, L06501. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.R. The seasonality of Antarctic sea ice trends. Geophys. Res. Lett. 2014, 41, 4230–4237. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, C.L. A 40-y record reveals gradual Antarctic sea ice increases followed by decreases at rates far exceeding the rates seen in the Arctic. Proc. Natl. Acad. Sci. USA 2019, 116, 14414–14423. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, C.L.; Cavalieri, D.J. Antarctic sea ice variability and trends, 1979–2010. Cryosphere 2012, 6, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Rosenblum, E.; Eisenman, I. Sea Ice Trends in Climate Models Only Accurate in Runs with Biased Global Warming. J. Clim. 2017, 30, 6265–6278. [Google Scholar] [CrossRef]
- Fox-Kemper, B.; Hewitt, H.T.; Xiao, C.; Aðalgeirsdóttir, G.; Drijfhout, S.S.; Edwards, T.L.; Golledge, N.R.; Hemer, M.; Kopp, R.E.; Krinner, G.; et al. Ocean, Cryosphere and Sea Level Change. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Thomas, E.R.; Allen, C.S.; Etourneau, J.; King, A.C.F.; Severi, M.; Winton, V.H.L.; Müller, J.; Crosta, X.; Peck, V.L. Antarctic Sea Ice Proxies from Marine and Ice Core Archives Suitable for Reconstructing Sea Ice over the past 2000 Years. Geosciences 2019, 9, 509. [Google Scholar] [CrossRef] [Green Version]
- Zonneveld, K.A.F.; Versteegh, G.J.M.; Kasten, S.; Eglinton, T.I.; Emeis, K.C.; Huguet, C.; Koch, B.P.; de Lange, G.J.; de Leeuw, J.W.; Middelburg, J.J.; et al. Selective preservation of organic matter in marine environments; processes and impact on the sedimentary record. Biogeosciences 2010, 7, 483–511. [Google Scholar] [CrossRef] [Green Version]
- Belt, S.T.; Smik, L.; Brown, T.A.; Kim, J.H.; Rowland, S.J.; Allen, C.S.; Gal, J.K.; Shin, K.H.; Lee, J.I.; Taylor, K.W.R. Source identification and distribution reveals the potential of the geochemical Antarctic sea ice proxy IPSO25. Nat. Commun. 2016, 7, 12655. [Google Scholar] [CrossRef] [Green Version]
- Leventer, A.; Dunbar, R.B.; DeMaster, D.J. Diatom Evidence for Late Holocene Climatic Events in Granite Harbor, Antarctica. Paleoceanography 1993, 8, 373–386. [Google Scholar] [CrossRef]
- Leventer, A.; Dunbar, R.B. Recent Diatom Record of McMurdo Sound, Antarctica: Implications for History of Sea Ice Extent. Paleoceanography 1988, 3, 259–274. [Google Scholar] [CrossRef]
- Cunningham, W.L.; Leventer, A.; Andrews, J.T.; Jennings, A.E.; Licht, K. Late Pleistocene-Holocene marine condtions in the Ross Sea, Antarctica: Evidence from the diatom record. Holocene 1999, 9, 129–139. [Google Scholar] [CrossRef]
- Mezgec, K.; Stenni, B.; Crosta, X.; Masson-Delmotte, V.; Baroni, C.; Braida, M.; Ciardini, V.; Colizza, E.; Melis, R.; Salvatore, M.C.; et al. Holocene sea ice variability driven by wind and polynya efficiency in the Ross Sea. Nat. Commun. 2017, 8, 1334. [Google Scholar] [CrossRef] [Green Version]
- Minzoni, R.T.; Majewski, W.; Anderson, J.B.; Yokoyama, Y.; Fernandez, R.; Jakobsson, M. Oceanographic influences on the stability of the Cosgrove Ice Shelf, Antarctica. Holocene 2017, 27, 1645–1658. [Google Scholar] [CrossRef]
- Hemer, M.A.; Harris, P.T. Sediment core from beneath the Amery Ice Shelf, East Antarctica, suggests mid-Holocene ice-shelf retreat. Geology 2003, 31, 127–130. [Google Scholar] [CrossRef]
- Berg, S.; Wagner, B.; Cremer, H.; Leng, M.; Melles, M. Late Quaternary environmental and climate history of Rauer Group, East Antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 201–213. [Google Scholar] [CrossRef]
- Crosta, X.; Crespin, J.; Swingedouw, D.; Marti, O.; Masson-Delmotte, V.; Etourneau, J.; Goosse, H.; Braconnot, P.; Yam, R.; Brailovski, I.; et al. Ocean as the main driver of Antarctic ice sheet retreat during the Holocene. Glob. Planet. Change 2018, 166, 62–74. [Google Scholar] [CrossRef]
- Denis, D.; Crosta, X.; Barbara, L.; Massé, G.; Renssen, H.; Ther, O.; Giraudeau, J. Sea ice and wind variability during the Holocene in East Antarctica: Insight on middle–high latitude coupling. Quat. Sci. Rev. 2010, 29, 3709–3719. [Google Scholar] [CrossRef]
- Pike, J.; Crosta, X.; Maddison, E.J.; Stickley, C.E.; Denis, D.; Barbara, L.; Renssen, H. Observations on the relationship between the Antarctic coastal diatoms Thalassiosira antarctica Comber and Porosira glacialis (Grunow) Jørgensen and sea ice concentrations during the late Quaternary. Mar. Micropaleontol. 2009, 73, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Taylor, F.; McMinn, A. Late Quaternary Diatom Assemblages from Prydz Bay, Eastern Antarctica. Quat. Res. 2002, 57, 151–161. [Google Scholar] [CrossRef]
- McMinn, A. Late Holocene increase in sea ice extent in fjords of the Vestfold Hills, eastern Antarctica. Antarct. Sci. 2000, 12, 80–88. [Google Scholar] [CrossRef]
- McMinn, A.; Heijnis, H.; Harle, K.; McOrist, G. Late-Holocene climatic change recorded in sediment cores from Ellis Fjord, eastern Antarctica. Holocene 2001, 11, 291–300. [Google Scholar] [CrossRef]
- Allen, C.S.; Oakes-Fretwell, L.M.; Anderson, J.B.; Hodgson, D.A. A record of Holocene glacial and oceanographic variability in Neny Fjord, Antarctic Peninsula. Holocene 2010, 20, 551–564. [Google Scholar] [CrossRef]
- Peck, V.L.; Allen, C.S.; Kender, S.; McClymont, E.L.; Hodgson, D.A. Oceanographic variability on the West Antarctic Peninsula during the Holocene and the influence of upper circumpolar deep water. Quat. Sci. Rev. 2015, 119, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Taylor, F.; McMinn, A. Evidence from diatoms for Holocene climate fluctuation along the East Antarctic margin. Holocene 2001, 11, 455–466. [Google Scholar] [CrossRef]
- Taylor, F.; Whitehead, J.; Domack, E. Holocene paleoclimate change in the Antarctic Peninsula: Evidence from the diatom, sedimentary and geochemical record. Mar. Micropaleontol. 2001, 41, 25–43. [Google Scholar] [CrossRef]
- Alley, K.; Patacca, K.; Pike, J.; Dunbar, R.; Leventer, A. Iceberg Alley, East Antarctic Margin: Continuously laminated diatomaceous sediments from the late Holocene. Mar. Micropaleontol. 2018, 140, 56–68. [Google Scholar] [CrossRef]
- Rathburn, A.E.; Pichon, J.J.; Ayress, M.A.; DeDeckker, P. Microfossil and stable-isotope evidence for changes in Late Holocene palaeoproductivity and palaeoceanographic conditions in the Prydz Bay region of Antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 131, 485–510. [Google Scholar] [CrossRef]
- Campagne, P.; Crosta, X.; Houssais, M.N.; Swingedouw, D.; Schmidt, S.; Martin, A.; Devred, E.; Capo, S.; Marieu, V.; Closset, I.; et al. Glacial ice and atmospheric forcing on the Mertz Glacier Polynya over the past 250 years. Nat. Commun. 2015, 6, 6642. [Google Scholar] [CrossRef] [Green Version]
- Cremer, H.; Gore, D.; Melles, M.; Roberts, D. Palaeoclimatic significance of late Quaternary diatom assemblages from southern Windmill Islands, East Antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 195, 261–280. [Google Scholar] [CrossRef]
- Maddison, E.J.; Pike, J.; Dunbar, R. Seasonally laminated diatom-rich sediments from Dumont d’Urville Trough, East Antarctic Margin: Late-Holocene Neoglacial sea-ice conditions. Holocene 2012, 22, 857–875. [Google Scholar] [CrossRef]
- Campagne, P.; Crosta, X.; Schmidt, S.; Noëlle Houssais, M.; Ther, O.; Massé, G. Sedimentary response to sea ice and atmospheric variability over the instrumental period off Adélie Land, East Antarctica. Biogeosciences 2016, 13, 4205–4218. [Google Scholar] [CrossRef] [Green Version]
- Crosta, X.; Etourneau, J.; Orme, L.C.; Dalaiden, Q.; Campagne, P.; Swingedouw, D.; Goosse, H.; Massé, G.; Miettinen, A.; McKay, R.M.; et al. Multi-decadal trends in Antarctic sea-ice extent driven by ENSO–SAM over the last 2000 years. Nat. Geosci. 2021, 14, 156–160. [Google Scholar] [CrossRef]
- Kulbe, T.; Melles, M.; Verkulich, S.R.; Pushina, Z.V. East Antarctic Climate and Environmental Variability over the Last 9400 Years Inferred from Marine Sediments of the Bunger Oasis. Arct. Antarct. Alp. Res. 2001, 33, 223–230. [Google Scholar] [CrossRef]
- Crosta, X.; Crespin, J.; Billy, I.; Ther, O. Major factors controlling Holocene delta C-13(org) changes in a seasonal sea-ice environment, Adelie Land, East Antarctica. Glob. Biogeochem. Cycles 2005, 19, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Crosta, X.; Debret, M.; Denis, D.; Courty, M.A.; Ther, O. Holocene long- and short-term climate changes off Adélie Land, East Antarctica. Geochem. Geophys. Geosystems 2007, 8, Q11009. [Google Scholar] [CrossRef]
- Crosta, X.; Denis, D.; Ther, O. Sea ice seasonality during the Holocene, Adelie Land, East Antarctica. Mar. Micropaleontol. 2008, 66, 222–232. [Google Scholar] [CrossRef]
- Denis, D.; Crosta, X.; Schmidt, S.; Carson, D.S.; Ganeshram, R.S.; Renssen, H.; Bout-Roumazeilles, V.; Zaragosi, S.; Martin, B.; Cremer, M.; et al. Holocene glacier and deep water dynamics, Adélie Land region, East Antarctica. Quat. Sci. Rev. 2009, 28, 1291–1303. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, K.-C.; Lee, J.I.; Khim, B.-K.; Bak, Y.-S.; Lee, M.K.; Lee, J.; Domack, E.W.; Christ, A.J.; Yoon, H.I. Holocene paleoceanography of Bigo Bay, west Antarctic Peninsula: Connections between surface water productivity and nutrient utilization and its implication for surface-deep water mass exchange. Quat. Sci. Rev. 2018, 192, 59–70. [Google Scholar] [CrossRef]
- Etourneau, J.; Collins, L.G.; Willmott, V.; Kim, J.H.; Barbara, L.; Leventer, A.; Schouten, S.; Sinninghe Damsté, J.S.; Bianchini, A.; Klein, V.; et al. Holocene climate variations in the western Antarctic Peninsula: Evidence for sea ice extent predominantly controlled by changes in insolation and ENSO variability. Clim. Past 2013, 9, 1431–1446. [Google Scholar] [CrossRef] [Green Version]
- Sjunneskog, C.; Taylor, F. Postglacial marine diatom record of the Palmer Deep, Antarctic Peninsula (ODP Leg 178, Site 1098) 1. Total diatom abundance. Paleoceanography 2002, 17, PAL-4. [Google Scholar] [CrossRef]
- Taylor, F.; Sjunneskog, C. Postglacial marine diatom record of the Palmer Deep, Antarctic Peninsula (ODP Leg 178, Site 1098) 2. Diatom assemblages. Paleoceanography 2002, 17, PAL-2. [Google Scholar] [CrossRef]
- Leventer, A. The Fate of Antarctic “Sea-ice diatoms” and their use as palaeoenvironmental indicators. In Antarctic Sea Ice Biological, Processes, Interactions and Variability; Union, A.G., Ed.; American Geophysical Union: Washington, DC, USA, 1998; pp. 121–137. [Google Scholar]
- Leventer, A.; Domack, E.W.; Ishman, S.E.; Brachfeld, S.; McClennen, C.E.; Manley, P. Productivity cycles of 200–300 years in the Antarctic Peninsula region: Understanding linkages among the sun, atmosphere, oceans, sea ice, and biota. Geol. Soc. Am. Bull. 1996, 108, 1626–1644. [Google Scholar] [CrossRef]
- Roberts, S.J.; Monien, P.; Foster, L.C.; Loftfield, J.; Hocking, E.P.; Schnetger, B.; Pearson, E.J.; Juggins, S.; Fretwell, P.; Ireland, L.; et al. Past penguin colony responses to explosive volcanism on the Antarctic Peninsula. Nat. Commun. 2017, 8, 14914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbara, L.; Crosta, X.; Schmidt, S.; Masse, G. Diatoms and biomarkers evidence for major changes in sea ice conditions prior the instrumental period in Antarctic Peninsula. Quat. Sci. Rev. 2013, 79, 99–110. [Google Scholar] [CrossRef]
- Yoon, H.I.; Park, B.-K.; Kim, Y.; Kang, C.Y. Glaciomarine sedimentation and its paleoclimatic implications on the Antarctic Peninsula shelf over the last 15,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 185, 235–254. [Google Scholar] [CrossRef]
- Minzoni, R.T.; Anderson, J.B.; Fernandez, R.; Wellner, J.S. Marine record of Holocene climate, ocean, and cryosphere interactions: Herbert Sound, James Ross Island, Antarctica. Quat. Sci. Rev. 2015, 129, 239–259. [Google Scholar] [CrossRef]
- Barbara, L.; Crosta, X.; Leventer, A.; Schmidt, S.; Etourneau, J.; Domack, E.; Masse, G. Environmental responses of the Northeast Antarctic Peninsula to the Holocene climate variability. Paleoceanography 2016, 31, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Heroy, D.C.; Sjunneskog, C.; Anderson, J.B. Holocene climate change in the Bransfield Basin, Antarctic Peninsula: Evidence from sediment and diatom analysis. Antarct. Sci. 2008, 20, 69–87. [Google Scholar] [CrossRef]
- Michalchuk, B.R.; Anderson, J.B.; Wellner, J.S.; Manley, P.L.; Majewski, W.; Bohaty, S. Holocene climate and glacial history of the northeastern Antarctic Peninsula: The marine sedimentary record from a long SHALDRIL core. Quat. Sci. Rev. 2009, 28, 3049–3065. [Google Scholar] [CrossRef]
- Barcena, M.A.; Fabres, B.; Isla, E.; Flores, J.A.; Sierro, F.J.; Canals, M.; Palanques, A. Holocene neoglacial events in the Bransfield Strait (Antarctica). Palaeocenographic and palaeoclimatic significance. Sci. Mar. 2006, 70, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Barcena, M.A.; Isla, E.; Plaza, A.; Flores, J.A.; Sierro, F.J.; Masque, P.; Sanchez-Cabeza, J.A.; Palanques, A. Bioaccumulation record and paleoclimatic significance in the Western Bransfield Strait. The last 2000 years. Deep-Sea Res. Part Ii-Top. Stud. Oceanogr. 2002, 49, 935–950. [Google Scholar] [CrossRef]
- Kyrmanidou, A.; Vadman, K.J.; Ishman, S.E.; Leventer, A.; Brachfeld, S.; Domack, E.W.; Wellner, J.S. Late Holocene oceanographic and climatic variability recorded by the Perseverance Drift, northwestern Weddell Sea, based on benthic foraminifera and diatoms. Mar. Micropaleontol. 2018, 141, 10–22. [Google Scholar] [CrossRef]
- Barcena, M.A.; Gersonde, R.; Ledesma, S.; Fabres, J.; Calafat, A.M.; Canals, M.; Sierro, F.J.; Flores, J.A. Record of Holocene glacial oscillations in Bransfield Basin as revealed by siliceous microfossil assemblages. Antarct. Sci. 1998, 10, 269–285. [Google Scholar] [CrossRef]
- Milliken, K.T.; Anderson, J.B.; Wellner, J.S.; Bohaty, S.M.; Manley, P.L. High-resolution Holocene climate record from Maxwell Bay, South Shetland Islands, Antarctica. Geol. Soc. Am. Bull. 2009, 121, 1711–1725. [Google Scholar] [CrossRef]
- Yoon, H.I.; Yoo, K.C.; Park, B.K.; Kim, Y.; Khim, B.K.; Kang, C.Y. The origin of massive diamicton in Marian and Potter coves, King George Island, West Antarctica. Geosci. J. 2004, 8, 1–10. [Google Scholar] [CrossRef]
- Yoo, K.-C.; Yoon, H.I.; Kim, J.-K.; Khim, B.-K. Sedimentological, geochemical and palaeontological evidence for a neoglacial cold event during the late Holocene in the continental shelf of the northern South Shetland Islands, West Antarctica. Polar Res. 2009, 28, 177–192. [Google Scholar] [CrossRef]
- Yoon, H.I.; Yoo, K.C.; Bak, Y.S.; Lim, H.S.; Kim, Y.; Lee, J.I. Late Holocene cyclic glaciomarine sedimentation in a subpolar fjord of the South Shetland Islands, Antarctica, and its paleoceanographic significance: Sedimentological, geochemical, and paleontological evidence. Geol. Soc. Am. Bull. 2010, 122, 1298–1307. [Google Scholar] [CrossRef]
- Divine, D.V.; Koç, N.; Isaksson, E.; Nielsen, S.; Crosta, X.; Godtliebsen, F. Holocene Antarctic climate variability from ice and marine sediment cores: Insights on ocean-atmosphere interaction. Quat. Sci. Rev. 2010, 29, 303–312. [Google Scholar] [CrossRef]
- Ferry, A.J.; Crosta, X.; Quilty, P.G.; Fink, D.; Howard, W.; Armand, L.K. First records of winter sea ice concentration in the southwest Pacific sector of the Southern Ocean. Paleoceanography 2015, 30, 1525–1539. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.S.; Esper, O.; Gersonde, R. Last Glacial-Holocene climate variability in the Atlantic sector of the Southern Ocean. Quat. Sci. Rev. 2016, 135, 115–137. [Google Scholar] [CrossRef]
- Bianchi, C.; Gersonde, R. Climate evolution at the last deglaciation: The role of the Southern Ocean. Earth Planet. Sci. Lett. 2004, 228, 407–424. [Google Scholar] [CrossRef]
- Orme, L.C.; Crosta, X.; Miettinen, A.; Divine, D.V.; Husum, K.; Isaksson, E.; Wacker, L.; Mohan, R.; Ther, O.; Ikehara, M. Sea surface temperature in the Indian sector of the Southern Ocean over the Late Glacial and Holocene. Clim. Past 2020, 16, 1451–1467. [Google Scholar] [CrossRef]
- Tesi, T.; Belt, S.T.; Gariboldi, K.; Muschitiello, F.; Smik, L.; Finocchiaro, F.; Giglio, F.; Colizza, E.; Gazzurra, G.; Giordano, P.; et al. Resolving sea ice dynamics in the north-western Ross Sea during the last 2.6 ka: From seasonal to millennial timescales. Quat. Sci. Rev. 2020, 237, 106299. [Google Scholar] [CrossRef]
- Vorrath, M.E.; Müller, J.; Rebolledo, L.; Cárdenas, P.; Shi, X.; Esper, O.; Opel, T.; Geibert, W.; Muñoz, P.; Haas, C.; et al. Sea ice dynamics in the Bransfield Strait, Antarctic Peninsula, during the past 240 years: A multi-proxy intercomparison study. Clim. Past 2020, 16, 2459–2483. [Google Scholar] [CrossRef]
- Ashley, K.E.; Crosta, X.; Etourneau, J.; Campagne, P.; Gilchrist, H.; Ibraheem, U.; Greene, S.E.; Schmidt, S.; Eley, Y.; Massé, G.; et al. Exploring the use of compound-specific carbon isotopes as a palaeoproductivity proxy off the coast of Adélie Land, East Antarctica. Biogeosciences 2021, 18, 5555–5571. [Google Scholar] [CrossRef]
- Hogg, A.G.; Heaton, T.J.; Hua, Q.; Palmer, J.G.; Turney, C.S.M.; Southon, J.; Bayliss, A.; Blackwell, P.G.; Boswijk, G.; Bronk Ramsey, C.; et al. SHCal20 Southern Hemisphere Calibration, 0–55,000 Years cal BP. Radiocarbon 2020, 62, 759–778. [Google Scholar] [CrossRef]
- Esper, O.; Gersonde, R. New tools for the reconstruction of Pleistocene Antarctic sea ice. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 399, 260–283. [Google Scholar] [CrossRef]
- Gersonde, R.; Zielinski, U. The reconstruction of late Quaternary Antarctic sea-ice distribution-the use of diatoms as a proxy for sea-ice. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 162, 263–286. [Google Scholar] [CrossRef]
- Crosta, X.; Shukla, S.K.; Ther, O.; Ikehara, M.; Yamane, M.; Yokoyama, Y. Last Abundant Appearance Datum of Hemidiscus karstenii driven by climate change. Mar. Micropaleontol. 2020, 157, 101861. [Google Scholar] [CrossRef]
- Crosta, X.; Sturm, A.; Armand, L.; Pichon, J.J. Late Quaternary sea ice history in the Indian sector of the Southern Ocean as recorded by diatom assemblages. Mar. Micropaleontol. 2004, 50, 209–223. [Google Scholar] [CrossRef]
- Lazzara, L.; Nardello, I.; Ermanni, C.; Mangoni, O.; Saggiomo, V. Light environment and seasonal dynamics of microalgae in the annual sea ice at Terra Nova Bay, Ross Sea, Antarctica. Antarct. Sci. 2007, 19, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Perrin, R.A.; Lu, P.; Marchant, H.J. Seasonal variation in marine phytoplankton and ice algae at a shallow antarctic coastal site. Hydrobiologia 1987, 146, 33–46. [Google Scholar] [CrossRef]
- Ryan, K.G.; Hegseth, E.N.; Martin, A.; Davy, S.K.; O’Toole, R.; Ralph, P.J.; McMinn, A.; Thorn, C.J. Comparison of the microalgal community within fast ice at two sites along the Ross Sea coast, Antarctica. Antarct. Sci. 2006, 18, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarska, I.; Barbrick, N.E.; Ehrman, J.M.; Cant, G.P. Eucampia Index as an indicator of the Late Pleistocene oscillations of the winter sea-ice extent at the ODP Leg 119 Site 745B at the Kerguelen Plateau. Hydrobiologia 1993, 269/270, 103–112. [Google Scholar] [CrossRef]
- Fryxell, G.A.; Prasad, A.K.S.K. Eucampia antarctica var. recta (Mangin) stat. nov. (Biddulphiaceae, Bacillariophyceae): Life stages at the Weddell Sea ice edge. Phycologia 1990, 29, 27–38. [Google Scholar] [CrossRef]
- Allen, C.S. Proxy development: A new facet of morphological diversity in the marine diatom Eucampia antarctica (Castracane) Mangin. J. Micropalaeontology 2014, 33, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Leventer, A.; Domack, E.; Barkoukis, A.; McAndrews, B.; Murray, J. Laminations from the Palmer Deep: A diatom-based interpretation. Paleoceanography 2002, 17, 1–15. [Google Scholar] [CrossRef]
- Armand, L.K.; Crosta, X.; Romero, O.; Pichon, J.-J. The biogeography of major diatom taxa in Southern Ocean sediments: 1. Sea ice related species. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 223, 93–126. [Google Scholar] [CrossRef]
- Zielinski, U.; Gersonde, R. Diatom distribution in Southern Ocean surface sediments (Atlantic sector): Implications for palaeoenvironmental reconstructions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 129, 213–250. [Google Scholar] [CrossRef]
- Świło, M.; Majewski, W.; Minzoni, R.T.; Anderson, J.B. Diatom assemblages from coastal settings of West Antarctica. Mar. Micropaleontol. 2016, 125, 95–109. [Google Scholar] [CrossRef]
- Crosta, X.; Pichon, J.; Labracherie, M. Distribution of Chaetoceros resting spores in modern peri-Antarctic sediments. Mar. Micropaleontol. 1997, 29, 283–299. [Google Scholar] [CrossRef]
- Belt, S.T.; Masse, G.; Rowland, S.J.; Poulin, M.; Michel, C.; LeBlanc, B. A novel chemical fossil of palaeo sea ice: IP25. Org. Geochem. 2007, 38, 16–27. [Google Scholar] [CrossRef]
- Brown, T.A.; Belt, S.T.; Tatarek, A.; Mundy, C.J. Source identification of the Arctic sea ice proxy IP25. Nat. Commun. 2014, 5, 4197. [Google Scholar] [CrossRef] [PubMed]
- Cabedo-Sanz, P.; Belt, S.T.; Knies, J.; Husum, K. Identification of contrasting seasonal sea ice conditions during the Younger Dryas. Quat. Sci. Rev. 2013, 79, 74–86. [Google Scholar] [CrossRef]
- Fahl, K.; Stein, R. Modern seasonal variability and deglacial/Holocene change of central Arctic Ocean sea-ice cover: New insights from biomarker proxy records. Earth Planet. Sci. Lett. 2012, 351, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Fahl, K.; Stein, R. Biomarker distributions in surface sediments from the Kara and Laptev seas (Arctic Ocean): Indicators for organic-carbon sources and sea-ice coverage. Quat. Sci. Rev. 2013, 79, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Massé, G.; Rowland, S.J.; Sicre, M.A.; Jacob, J.; Jansen, E.; Belt, S.T. Abrupt climate changes for Iceland during the last millennium: Evidence from high resolution sea ice reconstructions. Earth Planet. Sci. Lett. 2008, 269, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Barbara, L.; Crosta, X.; Massé, G.; Ther, O. Deglacial environments in eastern Prydz Bay, East Antarctica. Quat. Sci. Rev. 2010, 29, 2731–2740. [Google Scholar] [CrossRef]
- Cárdenas, P.; Lange, C.B.; Vernet, M.; Esper, O.; Srain, B.; Vorrath, M.-E.; Ehrhardt, S.; Müller, J.; Kuhn, G.; Arz, H.W.; et al. Biogeochemical proxies and diatoms in surface sediments across the Drake Passage reflect oceanic domains and frontal systems in the region. Prog. Oceanogr. 2018, 174, 72–88. [Google Scholar] [CrossRef]
- Collins, L.G.; Allen, C.S.; Pike, J.; Hodgson, D.A.; Weckström, K.; Massé, G. Evaluating highly branched isoprenoid (HBI) biomarkers as a novel Antarctic sea-ice proxy in deep ocean glacial age sediments. Quat. Sci. Rev. 2013, 79, 87–98. [Google Scholar] [CrossRef]
- Massé, G.; Belt, S.T.; Crosta, X.; Schmidt, S.; Snape, I.; Thomas, D.N.; Rowland, S.J. Highly branched isoprenoids as proxies for variable sea ice conditions in the Southern Ocean. Antarct. Sci. 2011, 23, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Sinninghe Damsté, J.S.; Rijpstra, W.I.C.; Coolen, M.J.L.; Schouten, S.; Volkman, J.K. Rapid sulfurisation of highly branched isoprenoid (HBI) alkenes in sulfidic Holocene sediments from Ellis Fjord, Antarctica. Org. Geochem. 2007, 38, 128–139. [Google Scholar] [CrossRef]
- Smik, L.; Belt, S.T.; Lieser, J.L.; Armand, L.K.; Leventer, A. Distributions of highly branched isoprenoid alkenes and other algal lipids in surface waters from East Antarctica: Further insights for biomarker-based paleo sea-ice reconstruction. Org. Geochem. 2016, 95, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Vorrath, M.E.; Muller, J.; Esper, O.; Mollenhauer, G.; Haas, C.; Schefuss, E.; Fahl, K. Highly branched isoprenoids for Southern Ocean sea ice reconstructions: A pilot study from the Western Antarctic Peninsula. Biogeosciences 2019, 16, 2961–2981. [Google Scholar] [CrossRef] [Green Version]
- Belt, S.T. Source-specific biomarkers as proxies for Arctic and Antarctic sea ice. Org. Geochem. 2018, 125, 277–298. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.S.; Pike, J.; Pudsey, C.J.; Leventer, A. Submillennial variations in ocean conditions during deglaciation based on diatom assemblages from the southwest Atlantic. Paleoceanography 2005, 20, 16. [Google Scholar] [CrossRef]
- Armand, L.; Ferry, A.J.; Leventer, A. Advances in palaeo sea ice estimation. In Sea Ice; Thomas, D.N., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 2017; pp. 600–629. [Google Scholar]
- Ferry, A.J.; Prvan, T.; Jersky, B.; Crosta, X.; Armand, L.K. Statistical modeling of southern ocean marine diatom proxy and winter sea ice data: Model comparison and developments. Prog. Oceanogr. 2015, 131, 100–112. [Google Scholar] [CrossRef]
- Kang, S.H.; Fryxell, G.A. Fragilariopsis cylindrus (Grunow) Krieger-the most abundant diatom in water column assemblages of Antarctic marginal ice-edge zones. Polar Biol. 1992, 12, 609–627. [Google Scholar] [CrossRef]
- Pike, J.; Allen, C.S.; Leventer, A.; Stickley, C.E.; Pudsey, C.J. Comparison of contemporary and fossil diatom assemblages from the western Antarctic Peninsula shelf. Mar. Micropaleontol. 2008, 67, 274–287. [Google Scholar] [CrossRef]
- von Quillfeldt, C.H. The diatom Fragilariopsis cylindrus and its potential as an indicator species for cold water rather than for sea ice. Vie Milieu 2004, 54, 137–143. [Google Scholar]
- Ashley, K.E.; McKay, R.; Etourneau, J.; Jimenez-Espejo, F.J.; Condron, A.; Albot, A.; Crosta, X.; Riesselman, C.; Seki, O.; Massé, G.; et al. Mid-Holocene Antarctic sea-ice increase driven by marine ice sheet retreat. Clim. Past 2021, 17, 1–19. [Google Scholar] [CrossRef]
- Stickley, C.E.; Pike, J.; Leventer, A.; Dunbar, R.; Domack, E.W.; Brachfeld, S.; Manley, P.; McClennan, C. Deglacial ocean and climate seasonality in laminated diatom sediments, Mac.Robertson Shelf, Antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 227, 290–310. [Google Scholar] [CrossRef]
- Anderson, D.M. Attenuation of millennial-scale events by bioturbation in marine sediments. Paleoceanography 2001, 16, 352–357. [Google Scholar] [CrossRef]
- Liu, H.; Meyers, S.R.; Marcott, S.A. Unmixing deep-sea paleoclimate records: A study on bioturbation effects through convolution and deconvolution. Earth Planet. Sci. Lett. 2021, 564, 116883. [Google Scholar] [CrossRef]
- Crosta, X.; Pichon, J.J.; Burckle, L.H. Application of modern analog technique to marine Antarctic diatoms: Reconstruction of maximum sea-ice extent at the Last Glacial Maximum. Paleoceanography 1998, 13, 284–297. [Google Scholar] [CrossRef]
- Garrison, D.L. Antarctic Sea Ice Biota. Am. Zool. 1991, 31, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Hegseth, E.N.; Von Quillfeldt, C.H. Low phytoplankton biomass and ice algal blooms in the Weddell Sea during the ice-filled summer of 1997. Antarct. Sci. 2002, 14, 231–243. [Google Scholar] [CrossRef]
- Kang, S.H.; Kang, J.S.; Lee, S.; Chung, K.H.; Kim, D.; Park, M.G. Antarctic phytoplankton assemblages in the marginal ice zone of the northwestern Weddell Sea. J. Plankton Res. 2001, 23, 333–352. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-H.; Fryxell, G.A. Phytoplankton in the Weddell Sea, Antarctica: Composition, abundance and distribution in water-column assemblages of the marginal ice-edge zone during austral autumn. Mar. Biol. 1993, 116, 335–348. [Google Scholar] [CrossRef]
- Kellogg, D.E.; Kellogg, T.B. Microfossil distributions in modern Amundsen Sea sediments. Mar. Micropaleontol. 1987, 12, 203–222. [Google Scholar] [CrossRef]
- Leventer, A.; Dunbar, R. Diatom flux in McMurdo Sound, Antarctica. Mar. Micropaleontol. 1987, 12, 49–64. [Google Scholar] [CrossRef]
- Ligowski, R.; Godlewski, M.; Lukowski, A. Sea ice diatoms and ice edge planktonic diatoms at the northern limit of the Weddell Sea pack ice. Proc. NIPR Symp. Polar Biol. 1992, 5, 9–20. [Google Scholar]
- Saggiomo, M.; Poulin, M.; Mangoni, O.; Lazzara, L.; De Stefano, M.; Sarno, D.; Zingone, A. Spring-time dynamics of diatom communities in landfast and underlying platelet ice in Terra Nova Bay, Ross Sea, Antarctica. J. Mar. Syst. 2017, 166, 26–36. [Google Scholar] [CrossRef]
- Schloss, I.; Estrada, M. Phytoplankton composition in the Weddell-Scotia Confluence area during austral spring in relation to hyrdrography. Polar Biol. 1994, 14, 77–90. [Google Scholar] [CrossRef]
- Smetacek, V.; Scharek, R.; Gordon, L.I.; Eicken, H.; Fahrbach, E.; Rohardt, G.; Moore, S. Early spring phytoplankton blooms in ice platelet layers of the southern Weddell Sea, Antarctica. Deep-Sea Res. Part A-Oceanogr. Res. Pap. 1992, 39, 153–168. [Google Scholar] [CrossRef]
- Cefarelli, A.O.; Ferrario, M.E.; Almandoz, G.O.; Atencío, A.G.; Akselman, R.; Vernet, M. Diversity of the diatom genus Fragilariopsis in the Argentine Sea and Antarctic waters: Morphology, distribution and abundance. Polar Biol. 2010, 33, 1463–1484. [Google Scholar] [CrossRef] [Green Version]
- Rigual-Hernández, A.S.; Pilskaln, C.H.; Cortina, A.; Abrantes, F.; Armand, L.K. Diatom species fluxes in the seasonally ice-covered Antarctic Zone: New data from offshore Prydz Bay and comparison with other regions from the eastern Antarctic and western Pacific sectors of the Southern Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 161, 92–104. [Google Scholar] [CrossRef]
- Garrison, D.L.; Buck, C.E.; Fryxell, G. Algal assemblages in Antarctic pack ice and in ice-edge plankton. J. Phycol. 1987, 23, 564–572. [Google Scholar] [CrossRef]
- Cunningham, W.L.; Leventer, A. Diatom assemblages in surface sediments of the Ross Sea: Relationship to present oceanographic conditions. Antarct. Sci. 1998, 10, 134–146. [Google Scholar] [CrossRef]
- Leventer, A. Modern distribution of diatoms in sediments from the George-V-Coast, Antarctica. Mar. Micropaleontol. 1992, 19, 315–332. [Google Scholar] [CrossRef]
- Gleitz, M.; Grossmann, S.; Scharek, R.; Smetacek, V. Ecology of diatom and bacterial assemblages in water associated with melting summer sea ice in the Weddell Sea, Antarctica. Antarct. Sci. 1996, 8, 135–146. [Google Scholar] [CrossRef]
- Maddison, E.J.; Pike, J.; Leventer, A.; Domack, E.W. Deglacial seasonal and sub-seasonal diatom record from Palmer Deep, Antarctica. J. Quat. Sci. 2005, 20, 435–446. [Google Scholar] [CrossRef]
- Garrison, D.L.; Jeffries, M.O.; Gibson, A.; Coale, S.L.; Neenan, D.; Fritsen, C.; Okolodkov, Y.B.; Gowing, M.M. Development of sea ice microbial communities during autumn ice formation in the Ross Sea. Mar. Ecol. Prog. Ser. 2003, 259, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Assmy, P.; Smetacek, V.; Montresor, M.; Klaas, C.; Henjes, J.; Strass, V.H.; Arrieta, J.M.; Bathmann, U.; Berg, G.M.; Breitbarth, E.; et al. Thick-shelled, grazer-protected diatoms decouple ocean carbon and silicon cycles in the iron-limited Antarctic Circumpolar Current. Proc. Natl. Acad. Sci. USA 2013, 110, 20633–20638. [Google Scholar] [CrossRef] [Green Version]
- Bathmann, U.V.; Scharek, R.; Klaas, C.; Dubischar, C.D.; Smetacek, V. Spring development of phytoplankton biomass and composition in major water masses of the Atlantic sector of the Southern Ocean. Deep-Sea Res. Part Ii-Top. Stud. Oceanogr. 1997, 44, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Crosta, X.; Romero, O.; Armand, L.K.; Pichon, J.-J. The biogeography of major diatom taxa in Southern Ocean sediments: 2. Open ocean related species. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 223, 66–92. [Google Scholar] [CrossRef]
- Mohan, R.; Quarshi, A.A.; Meloth, T.; Sudhakar, M. Diatoms from the surface waters of the Southern Ocean during the austral summer of 2004. Curr. Sci. 2011, 100, 1323–1327. [Google Scholar]
- Fischer, G.; Gersonde, R.; Wefer, G. Organic carbon, biogenic silica and diatom fluxes in the marginal winter sea-ice zone and in the Polar Front Region: Interannual variations and differences in composition. Deep-Sea Res. Part Ii-Top. Stud. Oceanogr. 2002, 49, 1721–1745. [Google Scholar] [CrossRef]
- Burckle, L.H.; Cirilli, J. Origin of diatom ooze belt in the Southern Ocean; implications for late Quaterary paleoceanography. Micropaleontology 1987, 33, 82–86. [Google Scholar] [CrossRef]
- Mohan, R.; Shanvas, S.; Thamban, M.; Sudhakar, M. Spatial distribution of diatoms in surface sediments from the Indian sector of Southern Ocean. Curr. Sci. 2006, 91, 1495–1502. [Google Scholar]
- Medlin, L.; Priddle, J. (Eds.) Polar Marine Diatoms; British Antarctic Survey: Cambridge, UK, 1990. [Google Scholar]
- Esper, O.; Gersonde, R. Quaternary surface water temperature estimations: New diatom transfer functions for the Southern Ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 414, 1–19. [Google Scholar] [CrossRef]
- Escutia, C.; Warnke, D.; Acton, G.D.; Barcena, A.; Burckle, L.; Canals, M.; Frazee, C.S. Sediment distribution and sedimentary processes across the Antarctic Wilkes Land margin during the Quaternary. Deep-Sea Res. Part Ii-Top. Stud. Oceanogr. 2003, 50, 1481–1508. [Google Scholar] [CrossRef]
- Garrison, D.L.; Buck, K.R. Sea-ice algal communities in the Weddell Sea: Species composition in ice and plankton assemblages. In Marine Biology of Polar Regions and Effects of Stress on Marine Organisms; Gray, J.S., Christiansen, M.E., Eds.; Wiley & Jons Ltd: Chichester, UK, 1985; pp. 103–122. [Google Scholar]
- Buffen, A.; Leventer, A.; Rubin, A.; Hutchins, T. Diatom assemblages in surface sediments of the northwestern Weddell Sea, Antarctic Peninsula. Mar. Micropaleontol. 2007, 62, 7–30. [Google Scholar] [CrossRef]
- Taylor, F.; McMinn, A.; Franklin, D. Distribution of diatoms in surface sediments of Prydz Bay, Antarctica. Mar. Micropaleontol. 1997, 32, 209–229. [Google Scholar] [CrossRef]
- Krebs, W.N.; Lipps, J.H.; Burckle, L.H. Ice diatom floras, Arthur Harbor, Antarctica. Polar Biol. 1987, 7, 163–171. [Google Scholar] [CrossRef]
- Maddison, E.J.; Pike, J.; Leventer, A.; Dunbar, R.; Brachfeld, S.; Domack, E.W.; Manley, P.; McClennen, C. Post-glacial seasonal diatom record of the Mertz Glacier Polynya, East Antarctica. Mar. Micropaleontol. 2006, 60, 66–88. [Google Scholar] [CrossRef]
- Arrigo, K.R. Sea Ice Ecosystems. Annu. Rev. Mar. Sci. 2014, 6, 439–467. [Google Scholar] [CrossRef]
- McMinn, A.; Hodgson, D. Summer phytoplankton succession in Ellis Fjord, eastern Antarctica. J. Plankton Res. 1993, 15, 925–938. [Google Scholar] [CrossRef]
- Rózanska, M.; Gosselin, M.; Poulin, M.; Wiktor, J.M.; Michel, C. Influence of environmental factors on the development of bottom ice protist communities during the winter–spring transition. Mar. Ecol. -Prog. Ser. 2009, 386, 43–59. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwe, M.A.; Tedesco, L.; Arrigo, K.R.; Assmy, P.; Campbell, K.; Meiners, K.M.; Rintala, J.-M.; Selz, V.; Thomas, D.N.; Stefels, J. Microalgal community structure and primary production in Arctic and Antarctic sea ice: A synthesis. Elem. Sci. Anthr. 2018, 6, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Ahn, I.Y.; Chung, H.; Kang, J.S.; Kang, S.H. Diatom composition and biomass variability in nearshore waters of Maxwell Bay, Antarctica, during the 1992/1993 austral summer. Polar Biol. 1997, 17, 123–130. [Google Scholar] [CrossRef]
- Whitaker, T.M.; Richardson, M.G. Morphology and Chemical composition of a natural population of an ice-associated Antarctic marine diatom Navicula glaciei. J. Phycol. 1980, 16, 250–257. [Google Scholar] [CrossRef]
- Scott, P.; McMinn, A.; Hosie, G. Physical parameters influencing diatom community structure in Eastern Antarctic sea-ice. Polar Biol. 1994, 14, 507–517. [Google Scholar] [CrossRef]
- Medlin, L.; Hasle, G. Some Nitzschia and related diatom species from fast ice samples in the Arctic and Antarctic. Polar Biol. 1990, 10, 451–479. [Google Scholar] [CrossRef]
- Fiala, M.; Kuosa, H.; Kopczynska, E.E.; Oriol, L.; Delille, D. Spatial and seasonal heterogeneity of sea ice microbial communities in the first-year ice of Terre Adelie area (Antarctica). Aquat. Microb. Ecol. 2006, 43, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Medlin, L. The nomenclature and type locality of Berkeleya adeliensis (Bacillariophyceae): A correction. Plant Ecol. Evol. 2019, 152, 409–411. [Google Scholar] [CrossRef]
- Tanimura, Y.; Fukuchi, M.; Watanabe, K.; Moriwaki, K. Diatoms in Water Column and Sea-ice in Lutzhow-Holm Bay, Antarctica, and their Preservation in the Underlying Sediments. Bull. Natl. Sci. Museum. Tokyo Ser. C 1990, 18, 15–39. [Google Scholar]
- McMinn, A.; Bleakley, N.; Steinburner, K.; Roberts, D.; Trenerry, L. Effect of permanent sea ice cover and different nutrient regimes on the phytoplankton succession of fjords of the Vestfold Hills Oasis, eastern Antarctica. J. Plankton Res. 2000, 22, 287–303. [Google Scholar] [CrossRef] [Green Version]
- Fryxell, G.A. Thalssiosira australis Peragallo and T. lentiginosa (Janisch) G. Fryxell, comb. nov.: Two Antarctic diatoms (Bacillariophyceae). Phycologia 1977, 16, 95–104. [Google Scholar] [CrossRef]
- Ichinomiya, M.; Nakamachi, M.; Fukuchi, M.; Taniguchi, A. Population dynamics of an ice-associated diatom, Thalassiosira australis Peragallo, under fast ice near Syowa Station, East Antarctica, during austral summer. Polar Biol. 2008, 31, 1051–1058. [Google Scholar] [CrossRef]
- McMinn, A. Preliminary investigation of the contribution of fast-ice algae to the spring phytoplankton bloom in Ellis Fjord, eastern Antarctica. Polar Biol. 1996, 16, 301–307. [Google Scholar] [CrossRef]
- Ichinomiya, M.; Gomi, Y.; Nakamachi, M.; Honda, M.; Fukuchi, M.; Taniguchi, A. Temporal variations in the abundance and sinking flux of diatoms under fast ice in summer near Syowa Station, East Antarctica. Polar Sci. 2008, 2, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Hasle, G.R.; Syvertsen, E.E. Chapter 2: Marine Diatoms. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Leventer, A.; Armand, L.; Harwood, D.; Jordan, R.W.; Ligowski, R. (Eds.) New Approaches and Progress in the Use of Polar Marine Diatoms in Reconstructing Sea-Ice Distribution. In Antarctica: A Keystone in a Changing World – Online Proceedings of the 10th ISAES X, Santa Barbara, CA, USA, 26 August–1 September 2007; USGS Open-File Report 2007-1047, Extended Abstract 005, 4p; USGS: Liston, VA, USA, 2007. [Google Scholar]
- Cremer, H.; Roberts, D.; McMinn, A.; Gore, D.; Melles, M. The Holocene Diatom Flora of Marine Bays in the Windmill Islands, East Antarctica. Bot. Mar. 2003, 46, 82–106. [Google Scholar] [CrossRef]
- Rivera, P.; Cruces, F. Fragilaria striatula lyngbye: Una diatomea marina muy poco conocida para chile. Gayana. Botánica 2002, 59, 35–41. [Google Scholar] [CrossRef]
- Kang, J.S.; Kang, S.H.; Lee, J.H.; Lee, S. Seasonal variation of microalgal assemblages at a s fixed station in King George Island, Antarctica, 1996. Mar. Ecol. Prog. Ser. 2002, 229, 19–32. [Google Scholar] [CrossRef]
- Hasle, G.R.; Medlin, L.K.; Syvertsen, E.E. Synedropsis gen. nov., a genus of araphid diatoms associated with sea ice. Phycologia 1994, 33, 248–270. [Google Scholar] [CrossRef]
- Cefarelli, A.O.; Ferrario, M.E.; Vernet, M. Diatoms (Bacillariophyceae) associated with free-drifting Antarctic icebergs: Taxonomy and distribution. Polar Biol. 2016, 39, 443–459. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
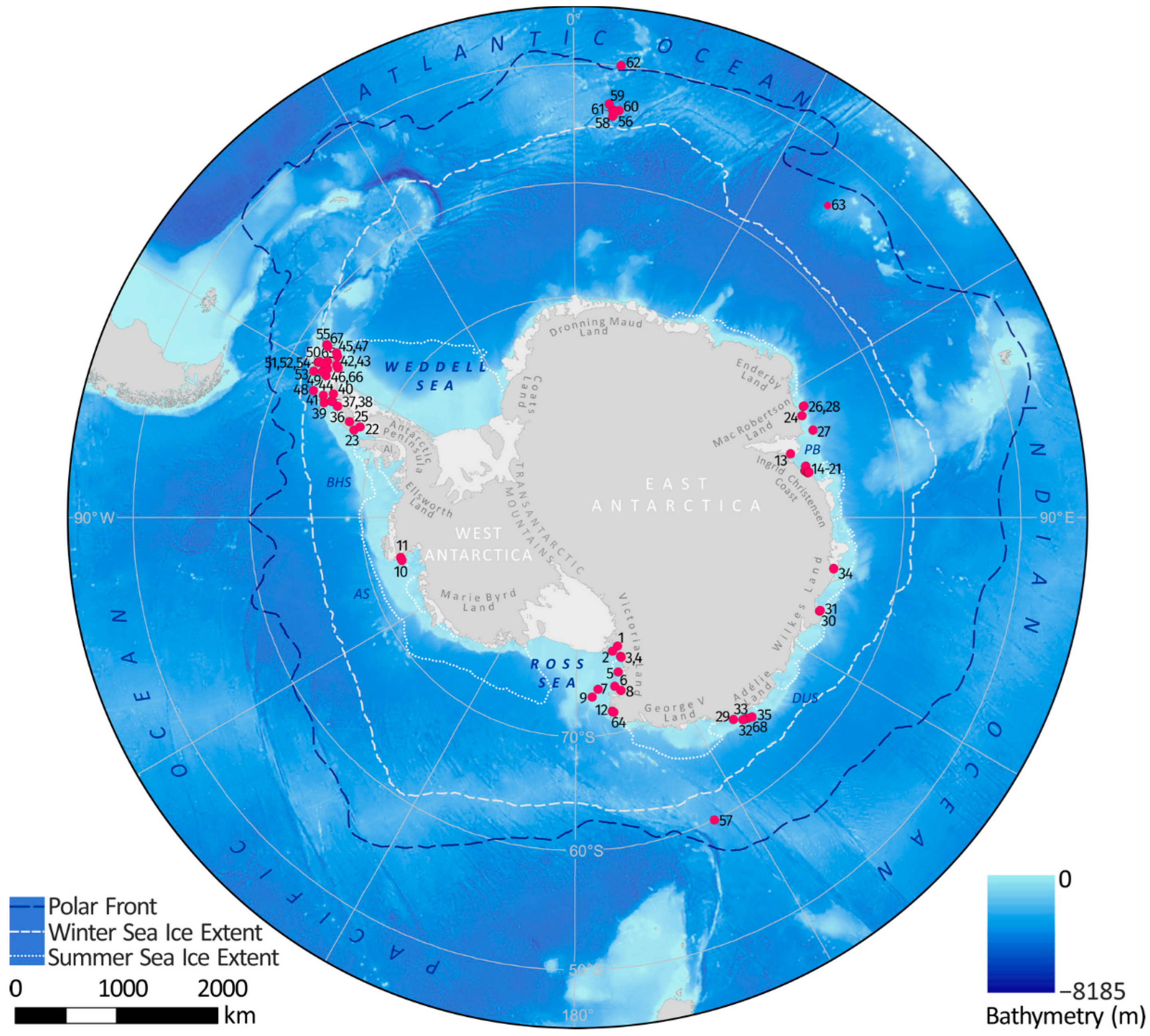
| Map Ref. | Core/Site ID | Lat | Long | Location Name | References |
|---|---|---|---|---|---|
| 1 | WG35 | −77.989 | 162.853 | Granite Harbor, Western Ross Sea | [11] |
| 2 | Multiple | −77.668 | 165.500 | McMurdo Sound; Western Ross Sea | [12] |
| 3 | WG17 | −77.000 | 162.850 | Granite Harbor, Western Ross Sea | [11] |
| 4 | KC208.09 | −76.972 | 162.876 | Granite Harbor, Western Ross Sea | [11] |
| 5 | KC31 | −75.700 | 165.418 | Western Ross Sea | [13] |
| 6 | KC37 | −74.499 | 167.744 | Western Ross Sea | [13] |
| 7 | KC39 | −74.474 | 173.474 | Western Ross Sea | [13] |
| 8 | BAY05-43c | −74.000 | 166.050 | Wood Bay, Western Ross Sea | [14] |
| 9 | ANTA99-cJ5 | −73.817 | 175.650 | Joides Basin, Western Ross Sea | [14] |
| 10 | KC17 | −73.420 | −102.827 | Ferrero Bay, Amundsen Sea Embayment | [15] |
| 11 | KC15 | −73.360 | −101.836 | Ferrero Bay, Amundsen Sea Embayment | [15] |
| 12 | BAY05-20c | −72.300 | 170.050 | Cape Hallet, Ross Sea | [14] |
| 13 | AM02 | −69.713 | 72.640 | Amery Ice Shelf, Prydz Bay | [16] |
| 14 | CO1011 | −68.827 | 77.760 | Flag Island Inlet, Prydz Bay | [17] |
| 15 | CO1010 | −68.817 | 77.833 | Filla Island Inlet, Prydz Bay | [17] |
| 16 | JPC24 | −68.694 | 76.709 | Svenner Channel, Prydz Bay | [18,19,20] |
| 17 | KROCK-15-GC29 | −68.664 | 76.696 | Prydz Bay | [21] |
| 18 | Abel Bay | −68.650 | 78.400 | Long Fjord, Ingrid Christensen Coast | [22] |
| 19 | Watts Basin | −68.603 | 78.213 | Ellis Fjord, Ingrid Christensen Coast | [23] |
| 20 | Deep Basin | −68.560 | 78.199 | Ellis Fjord, Ingrid Christensen Coast | [23] |
| 21 | Platcha Bay | −68.515 | 78.478 | Long Fjord, Ingrid Christensen Coast | [22] |
| 22 | JPC43 | −68.257 | −66.962 | Neny Fjord, Marguerite Bay, AP | [24] |
| 23 | TPC522 | −67.856 | −68.205 | Marguerite Bay, AP | [25] |
| 24 | KROCK-125-GC2 | −67.474 | 64.973 | Nielsen Bay, MacRobertson Land | [26] |
| 25 | GC1 | −67.180 | −66.797 | Lallemand Fjord, AP | [27] |
| 26 | JPC41 | −67.131 | 62.990 | Iceberg Alley, MacRobertson Land | [28] |
| 27 | GC 5 | −67.059 | 69.016 | MacRobertson Shelf, Prydz Bay | [29] |
| 28 | KROCK-128-GC1 | −66.983 | 63.154 | Iceberg Alley, MacRobertson Land | [26] |
| 29 | CB2010 | −66.906 | 142.436 | Commonwealth Bay, Adélie Land | [30] |
| 30 | PG1433 | −66.465 | 110.572 | Browning Bay, Windmill Is., Wilkes Land | [31] |
| 31 | PG1430 | −66.453 | 110.498 | Peterson Inlet, Windmill Is., Wilkes Land | [31] |
| 32 | MD03-2597 | −66.412 | 140.421 | Dumont d’Urville Trough, Adélie Land | [32] |
| 33 | DTCI2010; 318-1357B | −66.411 | 140.445 | Dumont d’Urville Trough, Adélie Land | [33,34] |
| 34 | PG1173 | −66.267 | 100.750 | Bunger Oasis, Wilkes Land | [35] |
| 35 | MD03-2601 | −66.052 | 138.557 | Dumont d’Urville Trough, Adélie Land | [20,36,37,38,39] |
| 36 | WAP13-GC47 | −65.613 | −64.759 | Bigo Bay, AP | [40] |
| 37 | JPC10/178-1098 | −64.883 | −64.200 | Palmer Deep, AP | [41,42,43] |
| 38 | PD92-30 | −64.862 | −64.208 | Palmer Deep, AP | [44,45] |
| 39 | TC46/GC47 | −64.588 | −64.805 | Anvers Shelf, AP | [46] |
| 40 | MTC18A | −64.772 | −62.829 | Andvord Drift, Gerlache Strait, AP | [47] |
| 41 | GC 02 | −64.000 | −64.000 | Anvers Shelf, AP | [48] |
| 42 | KC2B | −63.971 | −57.759 | Herbert Sound, James Ross Island, AP | [49] |
| 43 | MTC38C, JPC38 | −63.717 | −57.411 | Vega Drift, Prince Gustav Channel, AP | [47,50] |
| 44 | PC61 | −63.389 | −60.319 | Western Basin, Bransfield Strait, AP | [51] |
| 45 | JPC02 | −63.343 | −55.887 | Firth Of Tay, Joinville Island, AP | [52] |
| 46 | A-3 | −63.168 | −59.302 | Orleans Trough, Bransfield Strait, AP | [53,54] |
| 47 | JPC36 | −63.089 | −55.411 | Perseverance Drift, Joinville Island, AP | [55] |
| 48 | GC 03 | −63.000 | −64.000 | Anvers Shelf, AP | [48] |
| 49 | A-6 | −62.912 | −59.97 | Western Basin, Bransfield Strait, AP | [53,54] |
| 50 | Gebra-1 | −62.589 | −58.542 | Central Basin, Bransfield Strait, AP | [53,56] |
| 51 | 1B | −62.282 | −58.754 | Maxwell Bay, South Shetland Is. | [57] |
| 52 | MC-01 | −62.202 | −58.727 | Marian Cove, South Shetland Is. | [58] |
| 53 | WB2 | −62.200 | −60.700 | Outer Shelf, South Shetland Is. | [59] |
| 54 | CB2 | −62.191 | −58.833 | Collins Harbour, South Shetland Is. | [60] |
| 55 | Gebra-2 | −61.943 | −55.17 | Eastern Basin, Bransfield Strait, AP | [53,56] |
| 56 | 13PC (TN057-13PC4) | −53.200 | 5.1000 | Atlantic-Indian Ridge, S Atlantic Ocean | [61] |
| 57 | E27-23 | −59.618 | 155.238 | Emerald Basin, SE Indian Ocean | [62] |
| 58 | PS1652-2 | −53.664 | 5.100 | Atlantic-Indian Ridge, S Atlantic Ocean | [63] |
| 59 | PS1768-8 | −52.593 | 4.476 | Atlantic-Indian Ridge, S Atlantic Ocean | [63] |
| 60 | 177-1094/PS2090-1 | −53.179 | 5.132 | Atlantic-Indian Ridge, S Atlantic Ocean | [63,64] |
| 61 | PS2102-2 | −53.073 | 4.986 | Atlantic-Indian Ridge, S Atlantic Ocean | [63] |
| 62 | 17PC (TN057-17PC1) | −50.000 | 6.000 | Cape Basin, S Atlantic Ocean | [61] |
| 63 | COR1GC | −54.267 | 39.766 | Conrad Rise, SW Indian Ocean | [65] |
| 64 | HLF17-1 | −72.308 | −172.054 | Edisto Inlet, Ross Sea | [66] |
| 65 | PS97/056-1 | −64.757 | −60.442 | Gilbert Strait, Bransfield Strait, AP | [67] |
| 66 | PS97/068-2 | −63.168 | −59.302 | Orleands Trough, Bransfield Strait, AP | [67] |
| 67 | PS97/072-2 | −62.007 | −56.065 | Eastern Basin, Bransfield Strait, AP | [67] |
| 68 | DTGC2011 | −66.408 | 140.441 | Dumont d’Urville Trough, Adélie Land | [68] |
| Diatom Species | n |
|---|---|
| Fragilariopsis curta | 77 |
| Fragilariopsis cylindrus | 54 |
| Thalassiosira antarctica | 21 |
| Fragilariopsis sublinearis | 21 |
| Fragilariopsis kerguelensis | 19 |
| Fragilariopsis obliquecostata | 19 |
| Fragilariopsis rhombica | 16 |
| Fragilariopsis ritscheri | 14 |
| Fragilariopsis vanheurckii | 14 |
| Actinocyclus actinochilus | 12 |
| Porosira glacialis | 12 |
| Fragilariopsis separanda | 10 |
| Categories | n | |
|---|---|---|
| 1. | F. curta + F. cylindrus (F. c + cy) % | 10 |
| 2. | F. curta % | 23 |
| 3. | F. c + cy/T. antarctica | 3 |
| 4. | F. curta/F. kerguelensis | 4 |
| 5. | Groups including F. curta | 36 |
| 6. | Groups excluding F. curta | 7 |
| 7. | Other | 15 |
| 8. | HBIs | 14 |
| TOTAL | 112 |
| Map Ref. | Core/Site ID | Approximate Dates * | Approx. SR (m/ka) | Resolution (Lowest) ∆ | Resolution (Highest) ∆ | Proxy Categories † | References |
|---|---|---|---|---|---|---|---|
| 1 | WG35 | 1.2 to ~0.6 ka BP | 1.67 | 60 | 32 | 2 | [11] |
| 2 | Multiple | 0.5 to ~0 ka BP | 1 | 100 | 56 | 2 | [12] |
| 3 | WG17 | 1.2 to ~0.6 ka BP | 1.17 | 60 | 32 | 2 | [11] |
| 4 | KC208.09 | 1.3 to ~0 ka BP | 2.46 | 65 | 33 | 2 | [11] |
| 5 | KC31 | 2.0 to ~0 14C ka BP | 0.19 | 400 | 222 | 2 | [13] |
| 6 | KC37 | 2.0 to ~0 14C ka BP | 0.11 | 400 | 222 | 2 | [13] |
| 7 | KC39 | 2.0 to ~0 14C ka BP | 0.33 | 400 | 222 | 2 | [13] |
| 8 | BAY05-43c | 2.0 to ~0.0 ka BP | 2.13 | 38 | 15 | 2 | [14] |
| 9 | ANTA99-cJ5 | 2.0 to ~1.0 ka BP | 0.56 | 23 | 9 | 2 | [14] |
| 10 | KC17 | 2.0 to ~0 ka BP | 0.13 | 1000 | 500 | 7 | [15] |
| 11 | KC15 | 2.0 to ~0 ka BP | 0.13 | 1000 | 500 | 7 | [15] |
| 12 | BAY05-20c | 2.0 to ~0.1 ka BP | 1.58 | 48 | 19 | 2 | [14] |
| 13 | AM02 | 2.0 to ~0.0 14C ka BP | 0.05 | 400 | 222 | 2 | [16] |
| 14 | CO1011 | 2.0 to ~0 ka BP | 0.6 | 1000, 1000 | 500, 500 | 5, 7 | [17] |
| 15 | CO1010 | 2.0 to ~0 ka BP | 2 | 400, 400 | 222, 222 | 5, 7 | [17] |
| 16 | JPC24 | 2.0 to ~0.6 ka BP | 1.43 | 70, 70, 70, 70 | 36, 36, 36, 36 | 5, 5, 7, 8 | [18,19,20] |
| 17 | KROCK-15-GC29 | 2.0 to ~0 14C ka BP | 0.13 | 400 | 222 | 5 | [21] |
| 18 | Abel Bay | 2.0 to ~0 14C ka BP | 0.45 | 200, 200 | 51, 51 | 2, 6 | [22] |
| 19 | Watts Basin | 2.0 to ~0.2 14C ka BP | 1.39 | 45 | 18 | 6 | [23] |
| 20 | Deep Basin | 2.0 to ~0.8 14C ka BP | 2.42 | 30 | 12 | 6 | [23] |
| 21 | Platcha Bay | 2.0 to ~0.0 14C ka BP | 0.4 | 200, 200 | 105, 105 | 2, 6 | [22] |
| 22 | JPC43 | 2.0 to ~0 ka BP | 2.35 | 100 | 51 | 5 | [24] |
| 23 | TPC522 | 2.0 to ~0.8 ka BP | 0.33 | 240 | 133 | 4 | [25] |
| 24 | KROCK-125-GC2 | 2.0 to ~0 14C ka BP | 0.9 | 100 | 51 | 5 | [26] |
| 25 | GC1 | 2.0 to ~0 ka BP | 0.5 | 200, 200 | 105, 105 | 2, 5 | [27] |
| 26 | JPC41 | 2.0 to ~0 ka BP^ | 12 | <1^ | 2 | [28] | |
| 27 | GC 5 | 1.3 to ~0 14C ka BP | 0.14 | 700 | 175 | 2 | [29] |
| 28 | KROCK-128-GC1 | 2.0 to ~0.2 14C ka BP | 0.65 | 100 | 51 | 5 | [26] |
| 29 | CB2010 | 0.25 to ~0 ka BP | 1.4 | 6, 6 | 3, 3 | 7, 8 | [30] |
| 30 | PG1433 | 2.0 to ~0.3 ka BP | 2.18 | 85 | 44 | 6 | [31] |
| 31 | PG1430 | 2.0 to ~0 ka BP | 2.65 | 100 | 51 | 6 | [31] |
| 32 | MD03-2597 | 2.0 to ~0.7 ka BP^ | 20.54 | <1^ | 5 | [32] | |
| 33 | DTCI2010; 318-1357B | 0.04 to ~0 ka BP; 2,0 to 0.1 ka BP ** | 18.13, 19.14 | 19, 0.4, 19, 0.4 | 10, <0.4, 3, <0.4 | 1, 7, 8, 8 | [33,34] |
| 34 | PG1173 | 2.0 to ~0 ka BP | 1.45 | 50 | 20 | 5 | [35] |
| 35 | MD03-2601 | 2.0 to ~1.0 ka BP | 5 | 10, 25, 10, 25, 25, 50 | <10, 10, <10, 10, 10, 26 | 1, 4, 5, 5, 7, 8 | [20,36,37,38,39] |
| 36 | WAP13-GC47 | 2.0 to ~0.1 ka BP | 0.84 | 190 | 100 | 4 | [40] |
| 37 | JPC10/178-1098 | 2.0 to ~0.2 ka BP, 2.0 to 0 ka BP ** | 1.16, 2.00 | 173, 100, 100, 43 | 91, 51, 51, 17 | 1, 5, 7, 8 | [41,42,43] |
| 38 | PD92-30 | 2.0 to ~0.3 14C ka BP | 2.82 | 85, 85 | 44, 44 | 1, 3 | [44,45] |
| 39 | TC46/GC47 | 2.0 to ~0.1 ka BP | 0.42 | 100 | 190 | 4 | [46] |
| 40 | MTC18A | 0.13 to ~0 ka BP | 3.46 | 3, 3 | 1, 1 | 5, 8 | [47] |
| 41 | GC 02 | 2.0 to ~0.9 14C ka BP | 0.16 | 220, 220 | 122, 122 | 3, 5 | [48] |
| 42 | KC2B | 2.0 to ~0 ka BP | 1.15 | 200 | 105 | 6 | [49] |
| 43 | MTC38C, JPC38 | 0.08 to ~0 ka BP, 2.0 to ~0 ka BP ** | 5.75, 2.5 | 2, 2, 50, 20 | <1, <1, 20, <20 | 5, 8, 5, 8 | [47,50] |
| 44 | PC61 | 2.0 to ~0 ka BP | 0.5 | 200 | 105 | 2 | [51] |
| 45 | JPC02 | 2.0 to ~0 ka BP | 11 | 200 | 105 | 7 | [52] |
| 46 | A-3 | 1.7 to ~0 ka BP | 2.59 | 85 | 44 | 5 | [53,54] |
| 47 | JPC36 | 0.8 to ~0 ka BP | 17.5 | 40, 40 | 21, 21 | 5, 7 | [55] |
| 48 | GC 03 | 2.0 to ~0.1 14C ka BP | 0.32 | 188, 188 | 99, 99 | 3, 5 | [48] |
| 49 | A-6 | 1.8 to ~0.1 ka BP | 0.81 | 80 | 41 | 5 | [53,54] |
| 50 | Gebra-1 | 2.0 to ~0.2 ka BP | 0.72 | 90 | 46 | 5 | [53,56] |
| 51 | 1B | 2.0 to ~0.1 ka BP | 4.21 | 950 | 475 | 7 | [57] |
| 52 | MC-01 | 1.7 to ~0 14C ka BP | 2.18 | 17, 17 | <17, <17 | 2, 7 | [58] |
| 53 | WB2 | 1.5 to ~0 14C ka BP | 1.8 | 38 | 15 | 2 | [59] |
| 54 | CB2 | 2.0 to ~0 14C ka BP | 2.2 | 20, 20 | <20, <20 | 2, 7 | [60] |
| 55 | Gebra-2 | 1.6 to ~0.1 ka BP | 2.93 | 90 | 15 | 5 | [53,56] |
| 56 | 13PC (TN057-13PC4) | 2.0 to ~0 ka BP | 0.35 | 20 | <20 | 5 | [61] |
| 57 | E27-23 | 2.0 to ~1.5 ka BP | 0.5 | 500 | 100 | 5 | [62] |
| 58 | PS1652-2 | 2.0 to ~0.6 ka BP | 0.64 | 280, 280 | 156, 156 | 1, 5 | [63] |
| 59 | PS1768-8 | 2.0 to ~0.8 ka BP | 0.1 | 600, 600 | 300, 300 | 1, 5 | [63] |
| 60 | 177-1094/PS2090-1 | 2.0 to 1.0 ka BP, 2.0 to ~0.8 ka BP ** | 0.25, 0.30 | 240, 500, 240 | 133, 250, 133 | 1, 1, 5 | [63,64] |
| 61 | PS2102-2 | 2.0 to ~0.2 ka BP | 0.32 | 190, 190 | 100, 100 | 1, 5 | [63] |
| 62 | 17PC (TN057-17PC1) | 2.0 to ~0.0 ka BP | 0.25 | 40 | 16 | 5 | [61] |
| 63 | COR1GC | 2.0 to 1.0 ka BP | 0.25 | 52, 52 | 26, 26 | 5, 5 | [65] |
| 64 | HLF17-1 | 2.0 to 0.0 ka BP | 7.25 | 20, 20 | <20, <20 | 7, 8 | [66] |
| 65 | PS97/056-1 | 0.17 to ~0 ka BP | 2 | 17, 17, 9 | 9, 9, 4 | 2, 5, 8 | [67] |
| 66 | PS97/068-2 | 0.22 to ~0 ka BP | 1.95 | 11, 11, 6 | 6, 6, 2 | 2, 5, 8 | [67] |
| 67 | PS97/072-2 | 0.19 to ~0 ka BP | 2.05 | 10, 10, 5 | 6, 6, 2 | 2, 5, 8 | [67] |
| 68 | DTG2011 | 0.42 to ~0 ka BP | 11.17 | 4, 4 | <4, <4 | 1, 8 | [68] |
| Composition | Sites | References |
|---|---|---|
| Fragilariopsis curta, F. cylindrus, Navicula glaceii, and F. rhombica. | 14, 15 | [17] |
| Fragilariopsis curta, F. cylindrus, F. sublinearis, F. obliquecostata, F. vanheurckii, and Porosira glacialis | 16, 35 | [18,19] |
| Cluster groups: Coastal-CRS—Fragilariopsis curta, F. cylindrus, F. rhombica, and Pseudonitzschia turgiduloides; Shelf-CRS—F. curta, F. cylindrus, F. rhombica, Pentalamina corona a, Porosira glacialis, and Thalassiosira antarctica | 17, 24, 28 | [21,26] |
| PCA: Fragilariopsis curta, F. cylindrus, F. obliquecostata, F. ritscheri, F. sublinearis, and F. vanheurckii | 22 | [24] |
| Cluster group 1: dominated by Thalassiosira antarctica (T1 and T2) and Fragilariopsis curta, with F. cylindrus, F. rhombica, Navicula spp., Pentalamina corona a, Pseudonitzschia turgiduloides, Rhizosolenia spp., and Synedra spp. also present | 25 | [27] |
| Spring sea ice represented by laminae types A1, A2, and A3: CRS and Fragilariopsis spp. (with F. curta, F. cylindrus, and F. rhombica dominant) | 32 | [32] |
| Actinocyclus actinochiIus, Berkeleya rutilans, Entomoneis kufferathiib, Eucampia antarctica, Fragilariopsis angulatac, F. curta, F. cylindrus, F. obliquecostata, F. ritscheri, F. sublinearis, Porosira glacialls, P. pseudodenticulata, and Distephanus speculumd | 34 | [35] |
| The Fragilariopsis curta group: F. curta, F. cylindrus, and F. vanheurckii; and the Fragilariopsis cryophilic group: F. obliquecostata, F. ritscheri, and F. sublinearis | 35 | [38] |
| Cluster groups: Cocconeis assemblage dominated by Fragilariopsis curta and Thalassiosira antarctica, with Cocconeis as a unique indicator, and Corethron assemblage dominated by F. curta and F. cylindrus, with Corethron spp. and Pseudonitzschia turgiduloides as indicators | 37 | [43] |
| Fragilariopsis curta, F. cylindrus, and F. vanheurckii | 40, 43 | [47] |
| Sea ice taxa (not itemised) assume at least: Fragilariopsis curta and F. cylindrus (based on the use of ‘F. c + cy/T. antarctica’ ratio in the same publication) | 41, 48 | [48] |
| Fragilariopsis curta, F. cylindrus, F. sublinearis, and F. vanheurckii | 43, 47 | [50,55] |
| The sea ice taxa group: Fragilariopsis curta, F. cylindrus, F. sublinearis, F. obliquecostata, and F. vanheurckii | 46, 49, 50, 55 | [53,54,56] |
| MAT (31–33 taxa): Actinocyclus actinochilus, Alveus marinus e, Azpeitia tabularis, the Chaetoceros resting spore group, Fragilariopsis curta, F. cylindrus, F. doliolus, F. kerguelensis, F. obliquecostata, F. rhombica, F. ritscheri, F. separanda, F. sublinearis, Hemidiscus cuneiformis, Porosira glacialis, P. pseudodenticulata, Rhizosolenia antennata f. semispina, R. styliformis, Roperia tesselata, Stellarima microtrias, Thalassionema nitzschioides, T. nitzschioides var. lanceolata, T. nitzschioides var. parva, the Thalassiosira antarctica group (warm and cold morphologies e), the T. eccentrica group, Shionodiscus gracilis f, T. lentiginosa, S. oestrupii f, T. oliveriana, T. tumida, Thalassiothrix spp., and Trichotoxon reinboldii [72,73] | 56, 62, 63 | [61,65] |
| GAM: Actinocyclus actinochilus, Fragilariopsis curta, F. cylindrus, and Thalassiosira lentiginosa | 57 | [62] |
| MAT (28 taxa): Actinocyclus actinochilus, Azpeitia tabularis, Fragilariopsis curta, F. cylindrus, F. doliolus, F. kerguelensis, F. obliquecostata, F. rhombica, F. ritscheri, F. separanda, F. sublinearis, Hemidiscus cuneiformis, Nitzschia bicapitata, Porosira pseudodenticulata, Pseudonitzschia turgiduloides, Rhizosolenia spp., R. antennata f. semispina, R. bergonii, Roperia tesselata, Thalassionema nitzschioides f. 1, T. nitzschioides var. parva, T. nitzschioides var. lanceolata + T. nitzschioides var. capitulata, Thalassiosira antarctica, Shionodiscus gracilis e, T. lentiginosa, S. oestrupii e, T. oliveriana, and Thalassiothrix antarctica [70] | 58, 59, 60, 61, 65, 66, 67 | [63,67] |
| The sea ice group: Fragilariopsis curta, F. cylindrus, F. obliquecostata, F. ritscheri, Porosira glacialis, and Thalassiosira tumida | 63 | [65] |
| Composition | Sites | References |
|---|---|---|
| Fast ice index: Entomoneis kjellmannii, Nitzschia stellata, Berkeleya adelienses, Thalassiosira australis, Pleurosigma directum and Pinnularia quadreata | 18–21 | [22,23] |
| Fragilaria striulata, Navicula glaceii, and Synedropsis spp. | 30, 31 | [31] |
| Prolonged sea ice inferred from Navicula spp. | 42 | [49] |
| Composition | Sites | References |
|---|---|---|
| Entomoneis kjellmanii % | 14 and 15 | [17] |
| Eucampia index | 51 | [57] |
| F. cylindrus % | 29, 33, 52, and 54 | [30,33,58,60] |
| F. group/T. antarctica (T2) | 47 | [55] |
| Porosira glacialis/T antarctica | 16 and 35 | [20] |
| Pennate–centric ratios | 10 and 11 | [15] |
| Diatom concentrations | 37 and 45 | [42,52] |
| Frequency of dark–light laminae | 64 | [66] |
| Proxy Type | Advantages | Limitations |
|---|---|---|
| 1. F. curta + F. cylindrus | Well-established links between F. curta and F. cylindrus with sea ice melt waters, relative abundance in sediments over large areas of the deep Southern Ocean consistently linked with seasonal sea ice, based on large reference dataset of sediment traps and core tops throughout the deep South Atlantic | Proxy only ‘calibrated’ for Atlantic sector of the deep Southern Ocean |
| Both F. curta and F. cylindrus are widely preserved in marine sediments of the deep and continental shelf areas of the Southern Ocean | F. cylindrus not exclusive to sea ice meltwater but also found in glacial meltwaters and in stratified waters of the Ross Sea | |
| Can be successfully applied at several sites in the deep South Atlantic and over a variety of late Quaternary timescales | F. nana often not differentiated from F. cylindrus | |
| Well-defined morphology that minimizes risks of misidentification | Excludes broader assemblage information | |
| Small changes in the relative abundances of dominant species can have a pronounced impact on the percentages of minor taxa | ||
| 2. F. curta | Well-established association with seasonal sea ice throughout the deep Southern Ocean and the continental shelf | Association with seasonal sea ice fails over short/annual timeframe |
| Widely preserved and common in sediments throughout the deep Southern Ocean and the continental shelf | Relative abundances vary greatly from site to site | |
| Well-defined morphology that minimizes risks of misidentification | Excludes broader assemblage information | |
| 3. F. c + cy/T. antarctica | Ratio reduces influence of % changes in dominant species | Several morphotypes of T. antarctica and ambiguity over exact ecological associations—complicate identification and interpretation of ratio |
| Well-established association between F. c + cy and seasonal sea ice (see above) | Subjective boundaries between T. antarctica morphotypes compound identification difficulties and in particular compromise cross-site comparison | |
| 4. F. curta/F.kerguelensis | Ratio reduces influence of % changes in dominant species | Abundance of F. kerguelensis (strongly silicified) can be increased in sediments affected by dissolution |
| F. kerguelensis is well established as an open ocean diatom, so the ratio is a robust indicator for the relative influence of open ocean versus sea ice conditions | ||
| 5. Groups including F. curta | Groups incorporate information of broader assemblage | Difficult to have a group that is appropriate for a wide range of continental shelf sites |
| Groups can be tailored to local ocean conditions and/or assemblage composition | Transfer functions primarily built on reference data from deep ocean sites where seasonal sea ice expands and retreats along the north–south axis are not necessarily appropriate for continental shelf sites where sea ice distribution is more complex | |
| Many groups are statistically defined | Transfer functions only reliable within area of reference sites | |
| Transfer functions incorporate most species and produce quantitative results | ||
| 6. Groups excluding F. curta | Groups mostly tailored to specific nearshore conditions or particular types of sea ice | Most groups not suitable for application in offshore regions of the continental shelf or deep ocean regions |
| Sensitive to different types of sea ice | Sensitive to different types of sea ice rather than duration or distribution of sea ice cover | |
| 7. Other | Many based on routine information that can be applied alongside other species or assemblage proxies (e.g., Diatom concentration, pennate–centric, P. glaciailis/T. antarctica) | Most are only indirectly linked to sea ice |
| Can be widely applied throughout the continental shelf and deep ocean sites | May require additional analyses beyond the standard assemblage composition (e.g., morphometrics of the marine diatom Eucampia antarctica) | |
| 8. HBIs | Less time-intensive analyses, creating generally higher resolution records than traditional diatom assemblage data | Still requires considerable effort and personnel time to produce records |
| Absence of diene (HBI-II) can result from both open ocean and permanent sea ice conditions | ||
| Diene (HBI-II) linked to the sea ice diatom Berkeleya adeliensis | Environmental controls on the production of triene (HBI-III) are still ambiguous | |
| Widely preserved in sediments and robust at timescales up to at least 100 ka | Generally requires some validation with diatom assemblage data to aid interpretation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allen, C.S.; Weich, Z.C. Variety and Distribution of Diatom-Based Sea Ice Proxies in Antarctic Marine Sediments of the Past 2000 Years. Geosciences 2022, 12, 282. https://doi.org/10.3390/geosciences12080282
Allen CS, Weich ZC. Variety and Distribution of Diatom-Based Sea Ice Proxies in Antarctic Marine Sediments of the Past 2000 Years. Geosciences. 2022; 12(8):282. https://doi.org/10.3390/geosciences12080282
Chicago/Turabian StyleAllen, Claire S., and Zelna C. Weich. 2022. "Variety and Distribution of Diatom-Based Sea Ice Proxies in Antarctic Marine Sediments of the Past 2000 Years" Geosciences 12, no. 8: 282. https://doi.org/10.3390/geosciences12080282
APA StyleAllen, C. S., & Weich, Z. C. (2022). Variety and Distribution of Diatom-Based Sea Ice Proxies in Antarctic Marine Sediments of the Past 2000 Years. Geosciences, 12(8), 282. https://doi.org/10.3390/geosciences12080282