

Ostracod and Foraminifer Responses to Late Pleistocene–Holocene Volcanic Activity in Northern Victoria Land as Recorded in Ross Sea (Antarctica) Marine Sediments



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
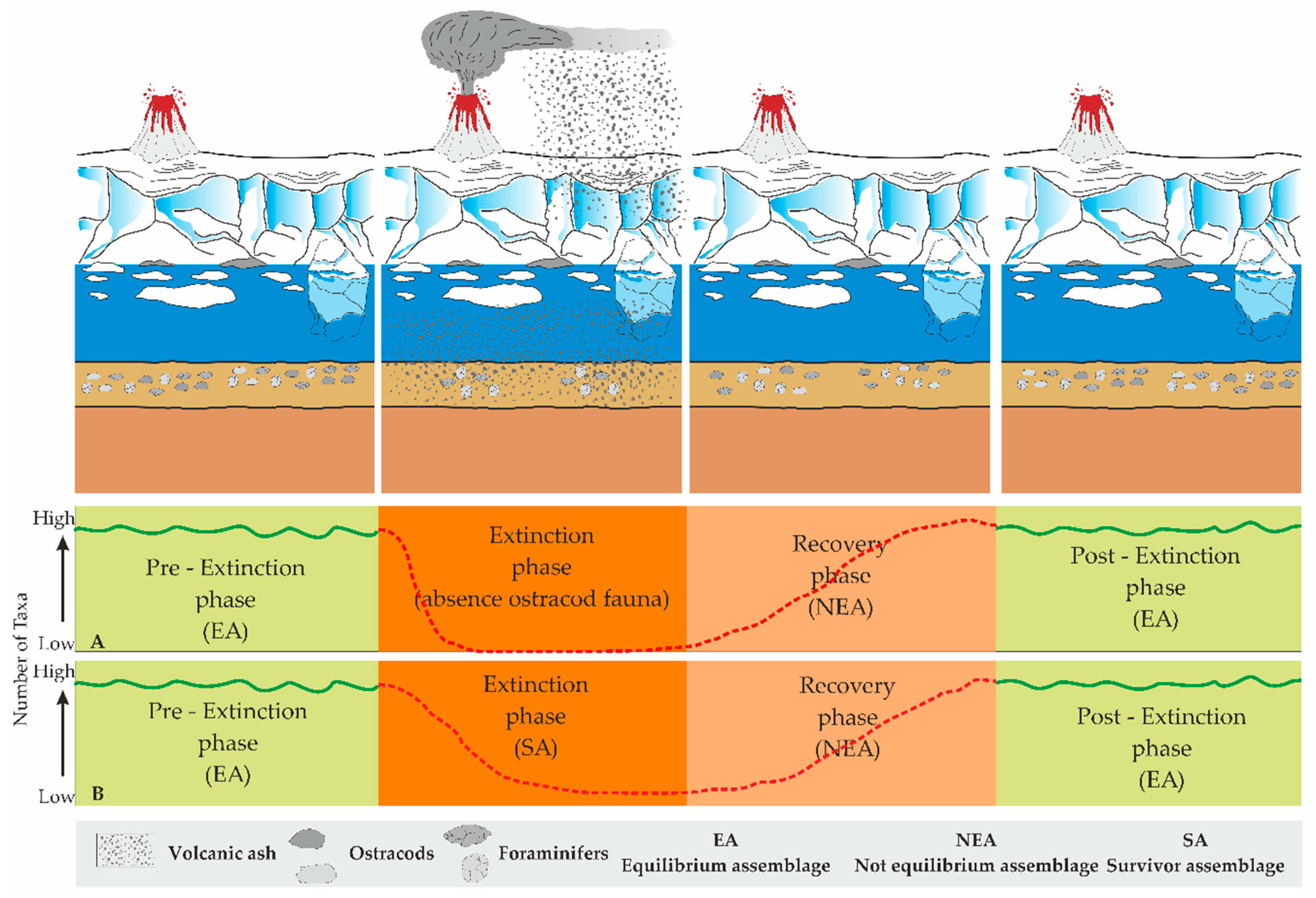
- The effects of the deposition of the tephra derived from explosive volcanic eruptions on ostracods and foraminifers;
- In relation to their response, to demonstrate the dynamics of the diverse phases of the communities in relation to the time of the tephra deposition (i.e., reduction, recolonisation, opportunistic species, and climax phases);
- To verify similarities or differences, if any, in the behaviour of ostracods and foraminifers in relation to volcanic events.
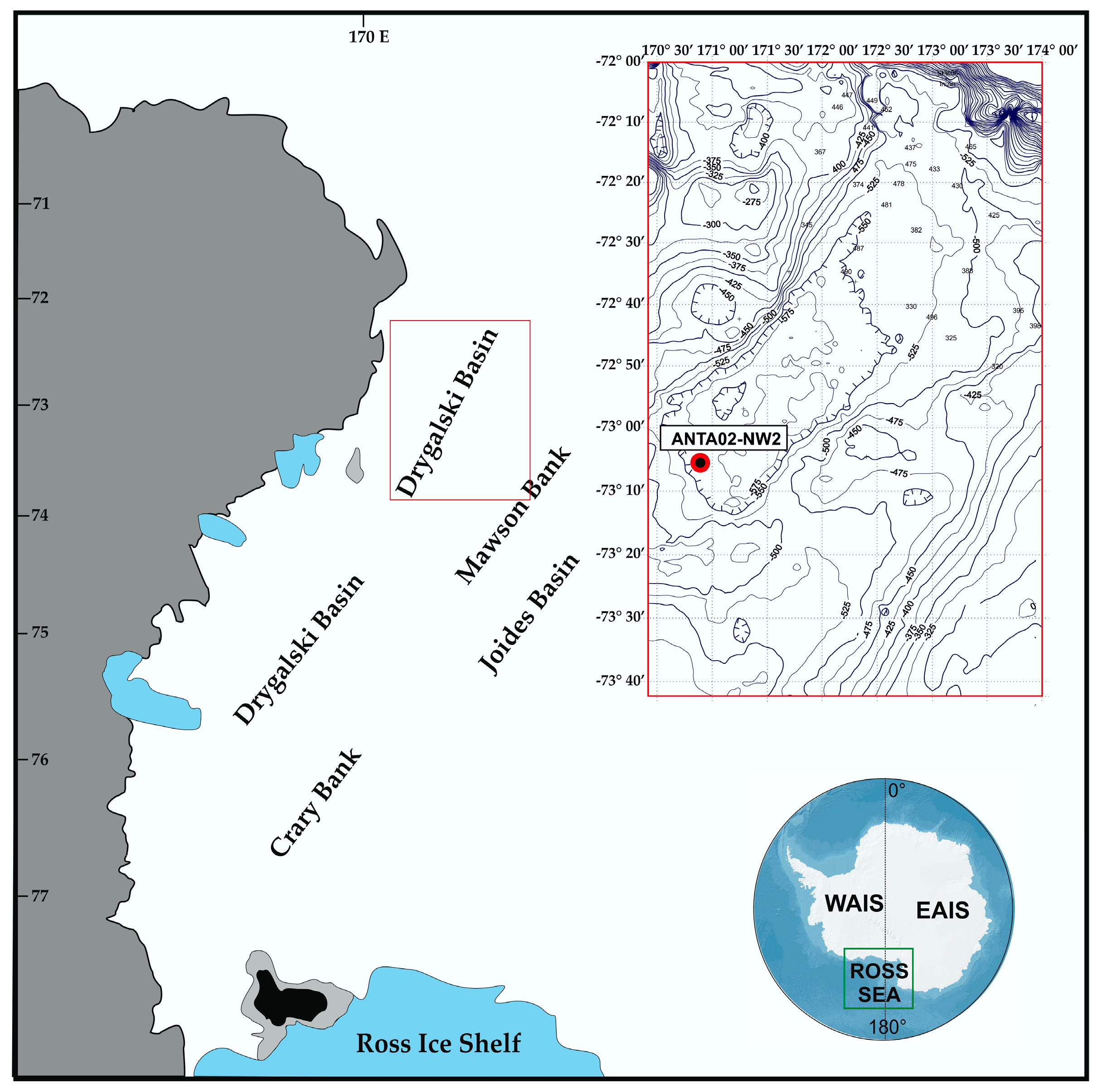
2. Materials and Methods
2.1. Sediment Analyses
- -
- Interval 0–13 cm: 13 cm thick, poorly sorted, massive coarse ash–medium lapilli of trachytic composition;
- -
- Interval 213–230 cm: 17 cm thick massive–normally graded bed of well-sorted medium ash of trachytic composition;
- -
- Interval 255–265 cm: 10 cm thick massive lens of poorly sorted coarse ash–lapilli with trachytic–phonolotic composition.
2.2. Data Analysis
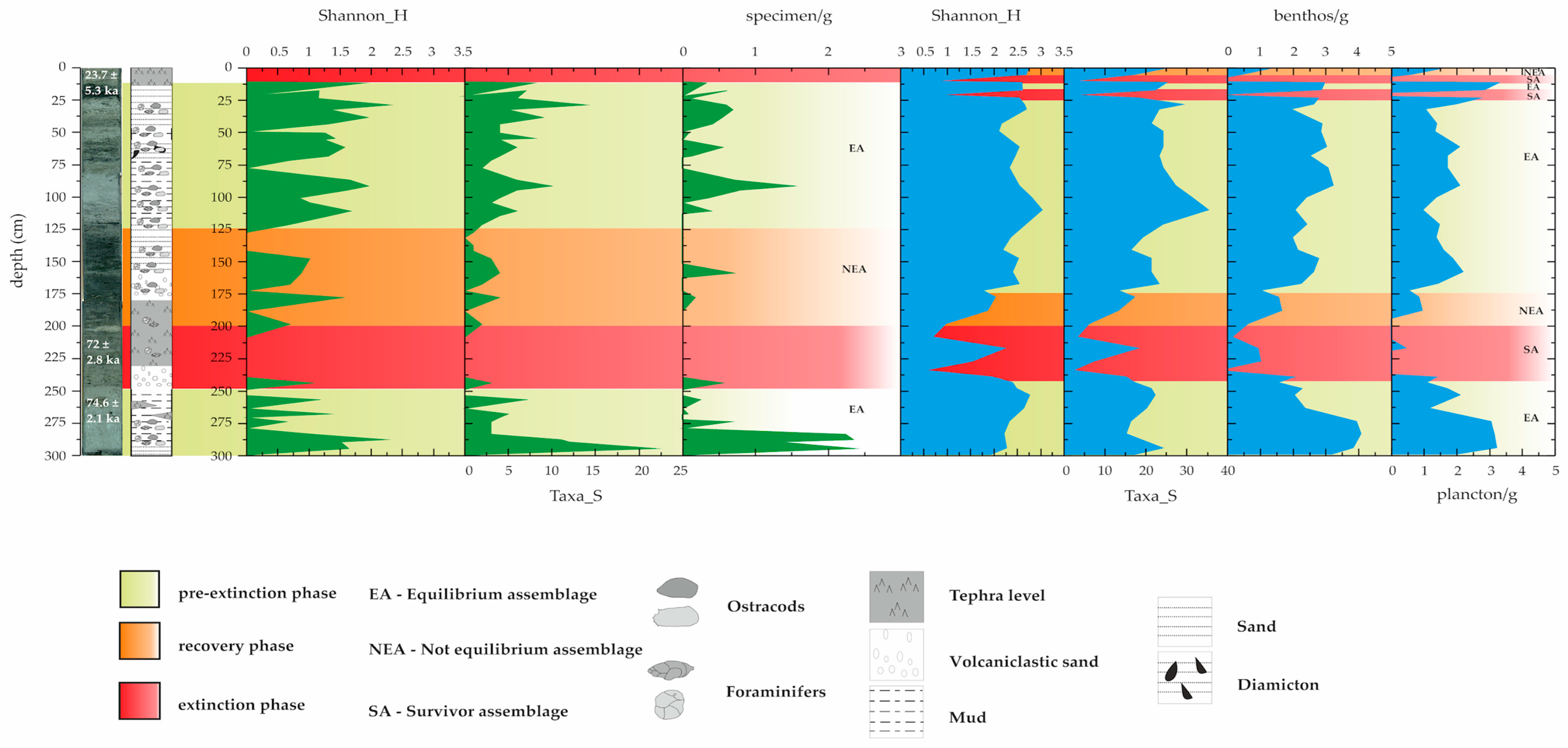
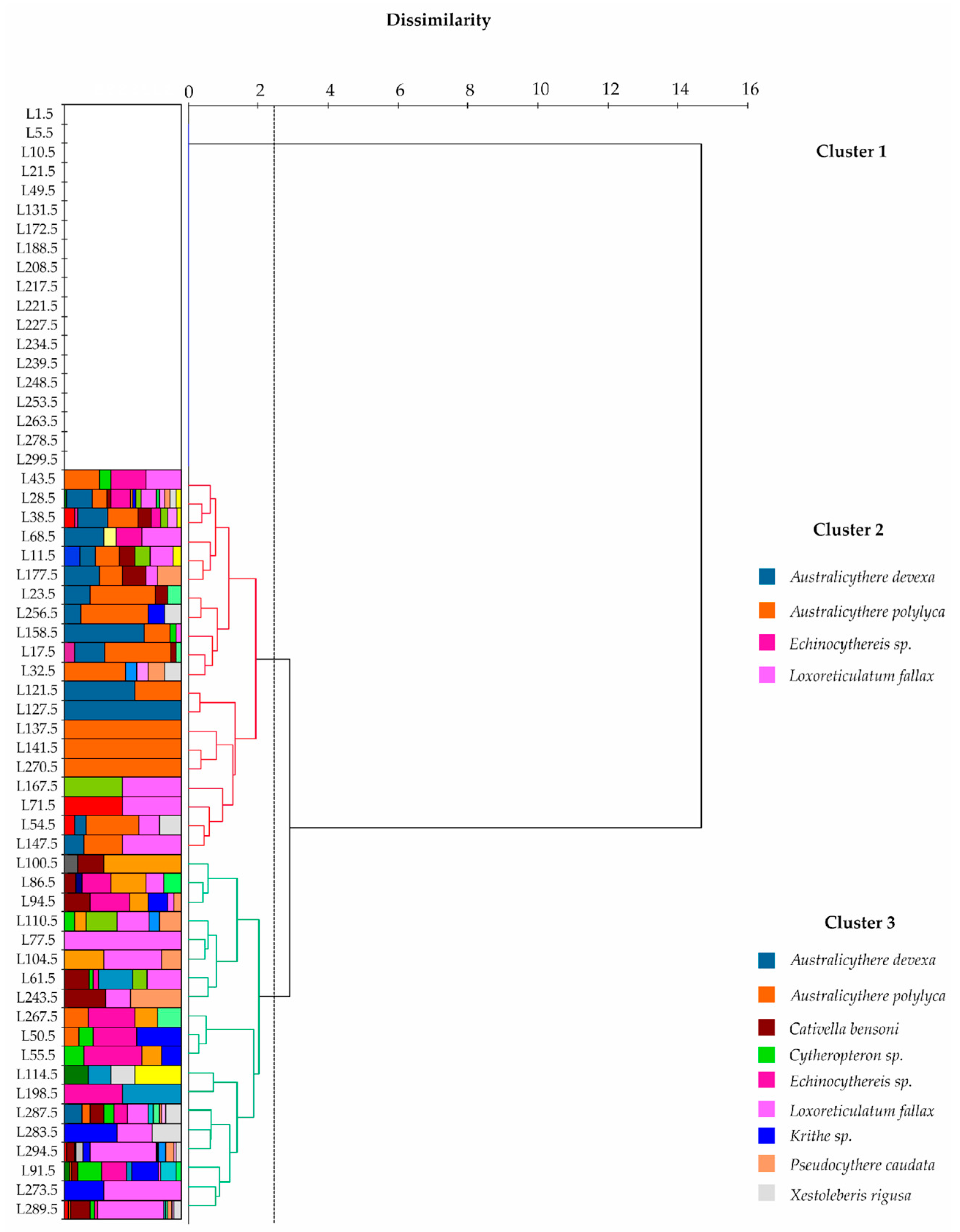
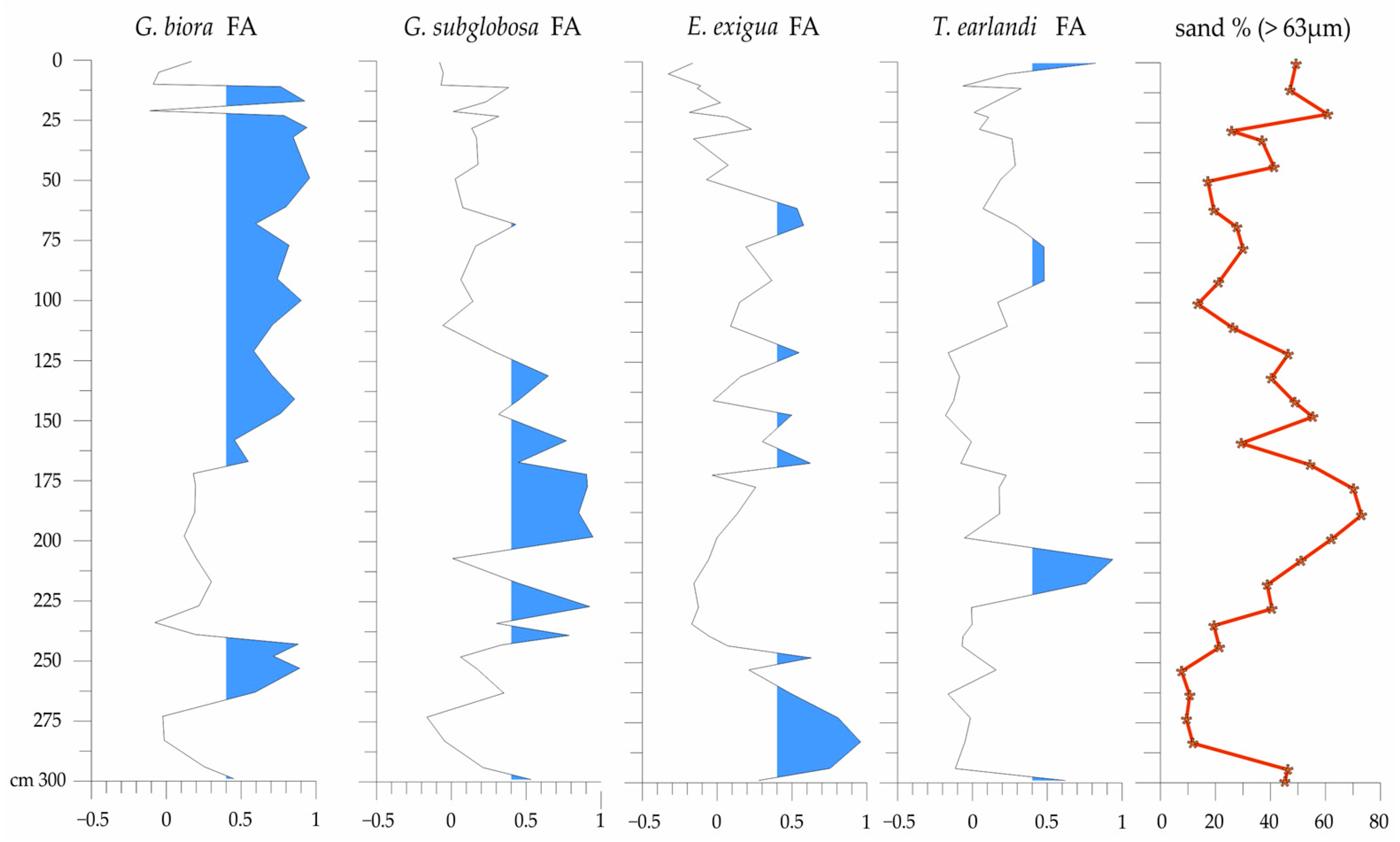
3. Results
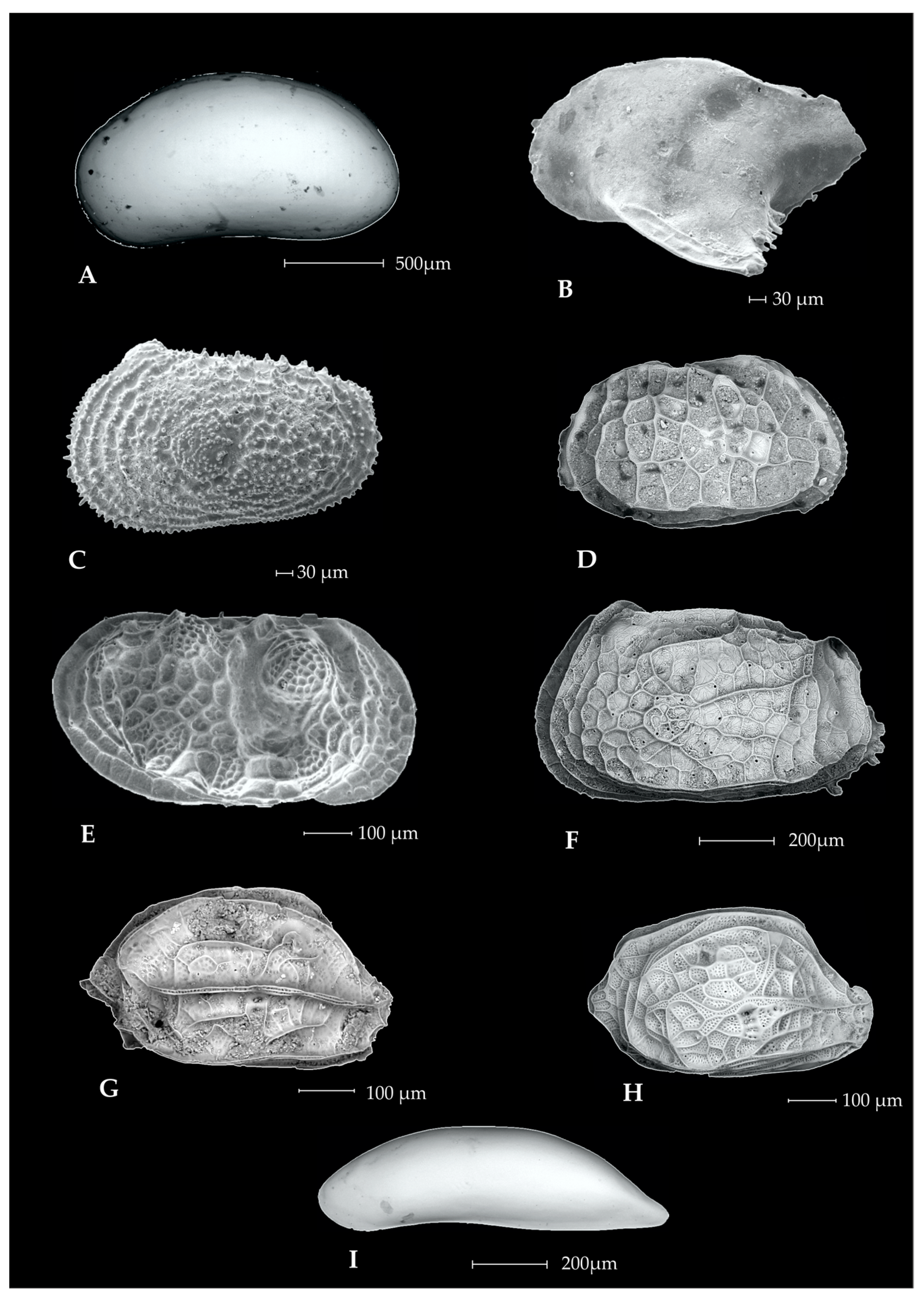
3.1. Ostracods
3.2. Foraminifers
4. Discussion
4.1. Ecological Evolutionary Phases in Sediments from Core NW2
4.2. Analysis of the Ostracod and Foraminiferal Assemblages with Respect to the Intervals of Core NW2
5. Conclusions
- A pre-extinction phase, characterised by high faunal abundance and diversity values;
- An extinction phase, marked by the complete disappearance of the ostracods; foraminifers, on the other hand, appear to be less sensitive than ostracods during volcanic episodes, recording extremely low values of abundance and diversity (SA assemblage);
- A recovery phase, featuring few dominant opportunistic species (NEA assemblage);
- A post-extinction phase, where the re-emergence (occurrence) of specialised taxa underlines the return to environmental stability (EA assemblage).
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Lowery, C.M.; Bralower, T.J.; Owens, J.D.; Rodríguez-Tovar, F.J.; Jones, H.; Smit, J.; Whalen, M.T.; Claeys, P.; Farley, K.; Gulick, S.P.S.; et al. Rapid recovery of life at ground zero of the end-Cretaceous mass extinction. Nature 2018, 558, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Hull, P.; Darroch, S.; Erwin, D. Rarity in mass extinctions and the future of ecosystems. Nature 2015, 528, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Gutt, J.; Bertler, N.; Bracegirdle, T.J.; Buschmann, A.; Comiso, J.; Hosie, G.; Isla, E.; Schloss, I.R.; Smith, C.R.; Tournadre, J.; et al. The Southern Ocean ecosystem under multiple climate stresses—An integrated circumpolar assessment. Glob. Chang. Biol. 2015, 21, 1434–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, H.J. Antarctic Marine Biodiversity—What Do We Know About the Distribution of Life in the Southern Ocean? PLoS ONE 2010, 5, e11683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, D.K.A.; Conlan, K.E. Disturbance, colonization and development of Antarctic benthic communities. Phil. Trans. R. Soc. 2006, 362, 11–38. [Google Scholar] [CrossRef]
- Rowden, A.A.; Kröger, K.; Clark, M.R. Compositional patterns of benthic assemblages on the northwestern Ross Sea shelf, Antarctica: Interacting environmental drivers operating at multiple spatial scales. Hydrobiologia 2015, 761, 211–233. [Google Scholar] [CrossRef] [Green Version]
- Gutt, J. Some “driving forces” structuring communities of the sublittoral Antarctic macrobenthos. Antarct. Sci. 2000, 12, 297–313. [Google Scholar] [CrossRef]
- Gutt, J.; Sirenko, B.; Smirnov, I.; Arntz, W. How many macrozoobenthic species might inhabit the Antarctic shelf? Antarct. Sci. 2004, 16, 11–16. [Google Scholar] [CrossRef]
- Gutt, J.; Griffiths, H.J.; Jones, C.D. Circumpolar overview and spatial heterogeneity of Antarctic macrobenthic communities. Mar. Biodivers. 2013, 43, 481–487. [Google Scholar] [CrossRef]
- Barry, J.P.; Grebmeier, J.M.; Smith, J.; Dunbar, R.B. Oceanographic Versus Seafloor-Habitat Control of Benthic Megafaunal Communities in the S.W. Ross Sea, Antarctica. Biogeochem. Ross Sea Antarct. Res. Ser. 2003, 78, 327–354. [Google Scholar] [CrossRef]
- Cummings, V.J.; Hewitt, J.E.; Thrush, S.F.; Marriott, P.M.; Halliday, N.J.; Norkko, A.M. Linking Ross Sea coastal benthic ecosystems to environmental conditions: Documenting baselines in a changing world. Front. Mar. Sci. 2018, 5, 232. [Google Scholar] [CrossRef]
- Cummings, V.J.; Bowden, D.A.; Pinkerton, M.H.; Halliday, N.J.; Hewitt, J.E. Ross Sea Benthic Ecosystems: Macro and Mega-faunal Community Patterns From a Multi-environment Survey. Front. Mar. Sci. 2021, 8, 629787. [Google Scholar] [CrossRef]
- Finger, K.L.; Lipps, J.H. Foraminiferal decimation and repopulation in an active volcanic caldera, Deception Island, Antarctica. Micropaleontology 1981, 27, 111–139. [Google Scholar] [CrossRef]
- Harper, D.A.T.; Scrutton, C.T.; Williams, D.M. Mass mortalities on an Irish Silurian seafloor. J. Geol. Soc. Lond. 1995, 152, 917–922. [Google Scholar] [CrossRef]
- Heikoop, J.M.; Tsujita, C.J.; Heikoop, C.E.; Risk, M.; Dickin, A. Effects of volcanic ashfall recorded in ancient marine benthic communities: Comparison of a near shore and an offshore environment. Lethaia 1996, 29, 125–139. [Google Scholar] [CrossRef]
- Hess, S.; Kuhnt, W.; Hill, S.; Kaminski, M.A.; Holbourn, A.; Leon, M. Monitoring the recolonization of the Mt Pinatubo 1991 ash layer by benthic foraminifera. Mar. Micropal. 2001, 43, 119–142. [Google Scholar] [CrossRef]
- Galeotti, S.; Bellagamba, M.; Kaminski, M.A.; Montanari, A. Deep-sea benthic foraminiferal recolonisation following a volcaniclastic event in the lower Campanian of the Scaglia Rossa Formation (Umbria–Marche Basin, central Italy). Mar. Micropal. 2002, 44, 57–76. [Google Scholar] [CrossRef]
- Keller, G. Biotic effects of impacts and volcanism. Earth Planet. Sci. Lett. 2003, 215, 249–264. [Google Scholar] [CrossRef]
- Kuhnt, W.; Hess, S.; Holbourn, A.; Paulsen, H.; Salomon, B. The impact of the 1991 Mt. Pinatubo eruption on deep-sea foraminiferal communities: A model for the Cretaceous–Tertiary (K/T) boundary? Palaeogeogr. Palaeoclim. Palaeoecol. 2005, 224, 83–107. [Google Scholar] [CrossRef]
- Wetzel, A. The preservation potential of ash layers in the deep-sea: The example of the 1991-Pinatubo ash in the South China Sea. Sedimentology 2009, 56, 1992–2009. [Google Scholar] [CrossRef]
- Waśkowska, A. Response of Early Eocene deep-water benthic foraminifera to volcanic ash falls in the Polish Outer Carpathians: Palaeocological implications. Palaeogeogr. Palaeoclim. Palaeoecol. 2011, 305, 50–64. [Google Scholar] [CrossRef]
- Charquenõ-Celis, F.; Sigala, I.; Zolitschka, B.; Pérez, L.; Christoph Mayr, C.; Massaferro, J. Responses of testate amoebae assemblages (Amoebozoa: Arcellinida) to recent volcanic eruptions, inferred from the sediment record in Laguna Verde, southern Patagonia, Argentina. J. Paleolimnol. 2022, 67, 115–129. [Google Scholar] [CrossRef]
- Gray, S.C.; Sturz, A.; Bruns, M.D.; Marzan, R.L.; Dougherty, D.; Law, H.B.; Brackett, J.E.; Marcou, M. Composition and distribution of sediments and benthic foraminifera in a submerged caldera after 30 years of volcanic quiescence. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2003, 50, 1727–1751. [Google Scholar] [CrossRef]
- Wilch, T.I.; McIntosh, W.C.; Dunbar, N.W. Late Quaternary volcanic activity in Marie Byrd Land: Potential 40 Ar/ 39 Ar-dated time horizons in West Antarctic ice and marine cores. Bull. Geol. Soc. Am. 1999, 111, 1563–1580. [Google Scholar] [CrossRef]
- Hillenbrand, C.-D.; Moreton, S.G.; Caburlotto, A.; Pudsey, C.J.; Lucchi, R.G.; Smellie, J.L.; Benetti, S.; Grobe, H.; Hunt, J.B.; Larter, R.D. Volcanic time-markers for Marine Isotopic Stages 6 and 5 in Southern Ocean sediments and Antarctic ice cores: Implications for tephra correlations between palaeoclimatic records. Quat. Sci. Rev. 2008, 27, 518–540. [Google Scholar] [CrossRef]
- Del Carlo, P.; Di Roberto, A.; Di Vincenzo, G.; Bertagnini, A.; Landi, P.; Pompilio, M.; Colizza, E.; Giordano, G. Late Pleistocene-Holocene volcanic activity in northern Victoria Land recorded in Ross Sea (Antarctica) marine sediments. Bull. Volcanol. 2015, 77, 36. [Google Scholar] [CrossRef]
- Di Roberto, A.; Colizza, E.; Del Carlo, P.; Petrelli, M.; Finocchiaro, F.; Kuhn, G. First marine cryptotephra in Antarctica found in sediments of the western Ross Sea correlates with englacial tephras and climate records. Sci. Rep. 2019, 9, 10628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Roberto, A.; Albert, P.G.; Colizza, E.; Del Carlo, P.; Di Vincenzo, G.; Gallerani, A.; Giglio, F.; Kuhn, G.; Macrì, P.; Manning, C.J.; et al. Evidence for a large-magnitude Holocene eruption of Mount Rittmann (Antarctica): A volcanological reconstruction using the marine tephra record. Quat. Sci. Rev. 2020, 250, 106629. [Google Scholar] [CrossRef]
- Di Roberto, A.; Del Carlo, P.; Pompilio, M. Marine record of Antarctic volcanism from drill cores. In Geological Society Memoir; Geological Society of London: London, UK, 2021; pp. 631–647. [Google Scholar] [CrossRef]
- Benson, R.H. Ostracods and Palaeoceanography. Ostracoda in the Earth Sciences. In Proceedings of the 11th International Symposium on Ostracoda, Warrnambool, Australia, 8–12 July 1991; De Deckker, P., Colin, J.P., Peypouquet, J.P., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1988; pp. 1–26. [Google Scholar]
- Cronin, T.M.; Holtz, T.R.; Whatley, R.C. Quaternary paleoceanography of the deep Arctic Ocean based on quantitative analysis of Ostracoda. Mar. Geol. 1994, 119, 305–332. [Google Scholar] [CrossRef]
- Cronin, T.M.; De Martino, D.M.; Dwyer, G.S.; Rodriguez-Lazaro, J. Deep-sea ostracode species diversity: Response to late Quaternary climate change. Mar. Micropal. 1999, 37, 231–249. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara, M.; Cronin, T.M.; Hunt, G.; Hodell, D.A. Deep–sea ostracods from the South Atlantic sector of the Southern Ocean during the Last 370,000 years. J. Paleontol. 2009, 83, 914–930. [Google Scholar] [CrossRef]
- Rodriguez-Lazaro, J.; Ruiz-Munõz, F. A General Introduction to Ostracods: Morphology, Distribution, Fossil Record and Applications. In Developments in Quaternary Sciences; Elsevier: Amsterdam, The Netherlands, 2012; Volume 17, pp. 1–14. [Google Scholar] [CrossRef]
- Cronin, T.M.; Raymo, M.E. Orbital forcing of deep-sea benthic species diversity. Nature 1997, 385, 624–627. [Google Scholar] [CrossRef]
- Cronin, T.; Dwyer, G.; Baker, P.A.; Rodriguez Lazaro, J.; Demartino, D.M. Orbital and suborbital variability in North Atlantic bottom water temperature obtained from deep-sea ostracode Mg/Ca ratios. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 162, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Didié, C.; Bauch, H.A.; Helmke, J.P. Late Quaternary deep-sea ostracodes in the polar and subpolar North Atlantic: Paleoecological and paleoenvironmental implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 184, 195–212. [Google Scholar] [CrossRef]
- Cronin, T.M.; Dwyer, G.S.; Farmer, J.; Bauch, H.A.; Spielhagen, R.F.; Jakobsson, M.; Nilsson, J.; Briggs, W.M., Jr.; Stepanova, A. Deep Arctic Ocean warming during the last glacial cycle. Nat. Geosc. 2012, 5, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Swain, F.M. Ostracode speciation following Middle Ordovician extinction events, North-Central United States. Geol. Soc. Lond. Spec. Publ. 1996, 102, 97–104. [Google Scholar] [CrossRef]
- Botting, J.P. The ecological effect of volcanic ash-fall in the marine Ordovician of Central Wales. Acta Univ. Carol. Geol. 1999, 43, 499–502. [Google Scholar]
- Botting, J.P. The role of pyroclastic volcanism in Ordovician diversification. In Palaeobiogeography and Biodiversity Change: The Ordovician and Mesozoic–Cenozoic Radiations; Crame, J.A., Owen, A.W., Eds.; Special Publications; Geological Society of London: London, UK, 2002; Volume 194, pp. 99–113. [Google Scholar]
- Botting, J.P.; Muir, L.A. Unravelling causal components of the Ordovician Radiation: The Builth Inlier (central Wales) as a case study. Lethaia 2008, 41, 111–125. [Google Scholar] [CrossRef]
- Hints, O.; Hints, L.; Meidla, T.; Sohar, K. Biotic effects of the Ordovician Kinnekulle ash-fall recorded in northern Estonia. Bull. Geol. Soc. Den. 2003, 50, 115–123. [Google Scholar] [CrossRef]
- Bodergat, A.M.; Ishizaki, K.; Oki, K.; Rio, M. Currents, civilization, or volcanism? Ostracodes as sentinels in a patchy environment: Kagoshima Bay, Japan. Micropaleontology 2002, 48, 285–299. [Google Scholar] [CrossRef]
- Perrier, V.; Meidla, T.; Oive, T.; Leho, A. Biotic response to explosive volcanism: Ostracod recovery after Ordovician ash-falls. Palaeogeogr. Palaeoclim. Palaeoecol. 2012, 365, 166–183. [Google Scholar] [CrossRef]
- Aiello, G.; Barra, D.; Parisi, R.; Isaia, R.; Marturano, A. Holocene benthic foraminiferal and ostracod assemblages in a paleo-hydrothermal vent system of Campi Flegrei (Campania, South Italy). Palaeont. Electr. 2018, 21, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: New York, NY, USA, 2006; p. 426. [Google Scholar]
- Smith, J.A.; Graham, A.G.C.; Post, A.L.; Hillebrand, C.-D.; Bart, P.J.; Powell, R.D. The marine geological imprint of Antarctic ice shelves. Nat. Commun. 2019, 10, 5635. [Google Scholar] [CrossRef] [Green Version]
- Majewski, W.; Prothro, L.O.; Simkins, L.M.; Demianiuk, E.J.; Anderson, J.B. Foraminiferal patterns in deglacial sediment in the western Ross Sea, Antarctica: Life near grounding lines. Paleoceanogr. Paleoclimatol. 2020, 35, e2019PA003716. [Google Scholar] [CrossRef]
- Melis, R.; Salvi, G. Foraminifer and Ostracod Occurrence in a Cool-Water Carbonate Factory of the Cape Adare (Ross Sea, Antarctica): A Key Lecture for the Climatic and Oceanographic Variations in the Last 30,000 Years. Geosciences 2020, 10, 413. [Google Scholar] [CrossRef]
- Prothro, L.O.; Majewski, W.; Yokoyama, Y.; Simkins, L.M.; Anderson, J.B.; Yamane, M.; Miyairi, Y.; Ohkouchi, N. Timing and pathways of East Antarctic Ice Sheet retreat. Quat. Sci. Rev. 2020, 230, 106166. [Google Scholar] [CrossRef]
- Bart, P.J.; Sjunneskog, C.; Chow, J.M. Piston-core based biostratigraphic constraints on Pleistocene oscillations of the West Antarctic Ice Sheet in western Ross Sea between North Basin and AND-1B drill site. Mar. Geol. 2011, 289, 86–99. [Google Scholar] [CrossRef]
- Halberstadt, A.R.W.; Simkins, L.M.; Greenwood, S.L.; Anderson, J.B. Past ice-sheet behaviour: Retreat scenarios and changing controls in the Ross Sea, Antarctica. Cryosphere 2016, 10, 1003–1020. [Google Scholar] [CrossRef] [Green Version]
- Davey, F.J. (Ed.) Ross Sea Bathymetry 1:2,000,000, Version 1.0; Institute of Geological & Nuclear Sciences Limited: Lower Hutt, New Zealand, 2004. [Google Scholar]
- Marigo, S. Foraminiferi e Dinamica della Piattaforma Glaciale del Mare di Ross Occidentale (Antartide): Considerazioni Paleoambientali. Master’s Thesis, University of Trieste, Trieste, Italy, 2021. [Google Scholar]
- Müller, G. Die Ostracodan der Deutschen Sudpolar-Expedition 1901–1903. Wissenschaftliche Ergebnisse der deutschen Sudpolar expedition. Zoologie 1908, 10, 51–181. [Google Scholar]
- Benson, R.H. Recent Cytheracean Ostracodes from McMurdo Sound and the Ross Sea, Antarctica. Arthropoda 1964, 6, 1–36. [Google Scholar]
- Neale, J.W. An ostracod fauna from Halley Bay, Coats Land, British Antarctic Territory. Br. Antarct. Surv. Sci. Rep. 1967, 58, 1–50. [Google Scholar]
- Hartmann, G. Antarktische benthische Ostracodan III. Auswertung der Reise des FFS “Walther Herwig” 68/1. 3. Teil: Süd-Orkney-Inseln. Mitt. Aus. Dem. Hambg. Zool. Mus. Und. Institut. 1988, 85, 141–162. [Google Scholar]
- Hartmann, G. Antarktische benthische Ostracodan IV. Auswertung der wahrend der Reise von FFS “Walther Herwig” (68/1) bei Sud-Georgien gesammelten Ostracodan. Mitt. Aus. Dem. Hambg. Zool. Mus. Und. Institut. 1989, 86, 209–230. [Google Scholar]
- Hartmann, G. Antarktische benthische Ostracodan, V. Auswertung der Sudwinterreise von FS “Polarstern” (Ps 9/V-1) im Bereich Elephant Island und der Antarktischen Halbinsel. Mitt. Aus. Dem. Hambg. Zool. Mus. Und. Institut. 1989, 86, 231–288. [Google Scholar]
- Hartmann, G. Antarktische benthische Ostracodan VI. Auswertung der Reise der “Polarstern” (Ant. VI-2). (1 Teil, Meiofauna und Zehnerserien sowie Versuch einer vorlaufigen Auswertung aller bislang vorliegenden Daten). Mitt. Aus. Dem. Hambg. Zool. Mus. Und. Institut. 1990, 87, 191–245. [Google Scholar]
- Hartmann, G. Antarktische benthische Ostracodan. VIII. Auswertung der Reise der “Meteor” (Ant. 11/4) in die Gewasser um Elephant Island und der Antarktischen Halbinsel. Helgoländer Meeresunters. 1992, 46, 405–424. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, G. Antarktische benthische Ostracodan IX. Ostracodan von der Antarktischen Halbinsel und von der Isla de los Estados (Feuerland/Argentinien). Auswertung der “Polarstern”- Reise PS ANT/X/1b. Mitt. Aus. Dem. Hambg. Zool. Mus. Institut 1993, 90, 227–237. [Google Scholar]
- Hartmann, G. Antarktische benthische Ostracoden X. Bemerkungen zur Gattung Krithe mit Beschreibung einer neuen Untergattung Austrokrithe. Mitt. Aus. Dem. Hambg. Zool. Mus. Institut 1994, 91, 77–79. [Google Scholar]
- Hartmann, G. Antarktische und Subantarktische Podocopa (Ostracoda). In Antarktische und Subantarktische Podocopa (Ostracoda); Koeltz Scientific Books: Koenigstein, Germany, 1997; Volume 7, pp. 1–355. [Google Scholar]
- Whatley, R.C.; Moguilevsky, A.; Ramos, M.I.F.; Coxill, D.J. Recent deep and shallow water Ostracoda from the Antarctic Peninsula and the Scotia Sea. Rev. Esp. Micropal. 1998, 30, 111–135. [Google Scholar]
- Dingle, R.V. Ostracoda from CRP–1 and CRP–2/2A, Victoria Land Basin, Antarctica. Terra Ant. 2000, 7, 479–492. [Google Scholar]
- Yasuhara, M.; Kato, M.; Ikeya, N.; Seto, K. Modern benthic ostracodes from Lützow-Holm Bay, East Antarctica: Paleoceanographic, paleobiogeographic, and evolutionary significance. Micropaleontology 2007, 53, 469–496. [Google Scholar] [CrossRef]
- Brenchley, P.J.; Harper, D.A.T. Palaeoecology: Ecosystems, Environments and Evolution; Chapman and Hall: London, UK, 1998. [Google Scholar]
- Boomer, I.; Horne, D.J.; Slipper, I.J. The Use of Ostracods in Palaeoenvironmental Studies, or What can you do with an Ostracod Shell? In Bridging the Gap: Trends in the Ostracode Biological and Geological Sciences; The Paleontological Society Papers: Cambridge, UK, 2003; Volume 9, pp. 153–180. [Google Scholar] [CrossRef]
- Brouwers, E.M. Paleobathymetry on the continental shelf based on examples using ostracods from the Gulf of Alaska. In Ostracoda in the Earth Sciences; De Deckker, P., Colin, J.-P., Peypouquet, J.-P., Eds.; Elsevier: Amsterdam, The Netherlands, 1988; pp. 55–76. [Google Scholar]
- Brouwers, E.M. Sediment transport detected from the analysis of ostracod population structures: An example from the Alaskan Continental Shelf. In Ostracoda in the Earth Sciences; De Deckker, P., Colin, J.-P., Peypouquet, J.-P., Eds.; Elsevier: Amsterdam, The Netherlands, 1988; pp. 231–244. [Google Scholar]
- Violanti, D. Taxonomy and distribution of recent benthic foraminifers from Terra Nova Bay (Ross Sea, Antarctica), Oceanographic Campaign 1987/1988. Palaeont. Ital. 1996, 83, 25–71. [Google Scholar]
- Majewski, W. Benthic foraminiferal distribution and ecology in Admiralty Bay, King George Island, West Antarctica. Pol. Polar Res. 2005, 26, 159–214. [Google Scholar]
- Majewski, W.; Bart, P.J.; McGlannan, A.J. Foraminiferal assemblages from ice-proximal paleo-settings in the Whales Deep Basin, Eastern Ross Sea, Antarctica. Palaeogeogr. Palaeoclim. Palaeoecol. 2018, 493, 64–81. [Google Scholar] [CrossRef]
- Malmgren, B.A.; Haq, B.U. Assessment of quantitative techniques in paleobiogeography. Mar. Micropal. 1982, 7, 213–236. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Sousa, W.P. The role of disturbance in natural communities. Ann. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; White, P.S. The Ecology of Natural Disturbance and Patch Dynamics; Academic Press: Orlando, FL, USA, 1985. [Google Scholar] [CrossRef]
- Sergio, F.; Blas, J.; Hiraldo, F. Animal responses to natural disturbance and climate extremes: A review. Glob. Planet. Chang. 2018, 161, 28–40. [Google Scholar] [CrossRef]
- Browning, T.J.; Stone, K.; Bouman, H.A.; Mather, T.A.; Pyle, D.M.; Moore, C.M.; Martinez-Vicente, V. Volcanic ash supply to the surface ocean: Remote sensing of biological responses and their wider biogeochemical significance. Front. Mar. Sci. 2015, 2, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Frogner, P.; Gislason, S.R.; Oskarsson, N. Fertilizing potential of volcanic ash in ocean surface water. Geology 2001, 29, 487–490. [Google Scholar] [CrossRef]
- Olgun, N.; Duggen, S.; Croot, P.L.; Delmelle, P.; Dietze, H.; Schacht, U.; Óskarsson, N.; Siebe, C.; Auer, A.; Garbe-Schönberg, D. Surface ocean iron fertilization: The role of airborne volcanic ash from subduction zone and hotspot volcanoes and related iron fluxes into the Pacific Ocean. Glob. Biogeochem. Cycles 2011, 25. [Google Scholar] [CrossRef]
- Durant, A.J.; Villarosa, G.; Rose, W.I.; Delmelle, P.; Prata, A.J.; Viramonte, J.G. Long-range volcanic ash transport and fallout during the 2008 eruption of Chaiten volcano, Chile. Phys. Chem. Earth 2012, 45–46, 50–64. [Google Scholar] [CrossRef]
- Lindenthal, A.; Langmann, B.; Paetsch, J.; Lorkowski, I.; Hort, M. The ocean response to volcanic iron fertilisation after the eruption of Kasatochi volcano: A regional scale biogeochemical ocean model study. Biogeosc. Discuss. 2012, 9, 9233–9257. [Google Scholar] [CrossRef] [Green Version]
- Achterberg, E.P.; Moore, C.M.; Henson, S.A.; Steigenberger, S.; Stohl, A.; Eckhardt, S.; Avendano, L.C.; Cassidy, M.; Hembury, D.; Klar, J.K.; et al. Natural iron fertilization by the Eyjafjallajokull volcanic eruption. Geophys. Res. Lett. 2013, 40, 921–926. [Google Scholar] [CrossRef] [Green Version]
- Morel, F.M.M.; Milligan, A.J.; Saito, M.A. Marine bioinorganic chemistry: The role of trace metals in the ocean cycles of major nutrients. Treatise Geochem. 2003, 6, 113–143. [Google Scholar] [CrossRef]
- Hoffmann, L.J.; Breitbarth, E.; Ardelan, M.V.; Duggen, S.; Olgun, N.; Hassellov, M.; Wängberg, S.-Å. Influence of trace metal release from volcanic ash on growth of Thalassiosira pseudonana and Emiliania huxleyi. Mar. Chem. 2012, 132, 28–33. [Google Scholar] [CrossRef]
- Mitchell, J.K. Fundamentals of Soil Behavior; Wiley and Sons: New York, NY, USA, 1993; pp. 1–437. [Google Scholar]
- Haeckel, M.; Beusekom, J.V.; Wiesner, M.G.; König, I. The impact of the 1991 Mount Pinatubo tephra fallout on the geochemical environment of the deep-sea sediments in the South China Sea. Earth Planet. Sci. Lett. 2001, 193, 151–166. [Google Scholar] [CrossRef]
- Pasotti, F.; Manini, E.; Giovannelli, D.; Wölfl, A.C.; Monien, D.; Verleyen, E.; Braeckman, U.; Abele, D.; Vanreusel, d. Antarctic shallow water benthos in an area of recent rapid glacier retreat. Mar. Ecol. 2014, 36, 716–733. [Google Scholar] [CrossRef] [Green Version]
- Brandão, S.N.; Dingle, R.V. Biogeographic Atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H.J., Raymond, B., d’Udekem d’Acoz, C., Van de Putte, A.P., Danis, B., David, B., Grant., S., Gutt, J., Eds.; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 142–148. [Google Scholar]
- Ayress, M.; Neil, H.; Passlow, V.; Swanson, K.M. Benthonic ostracods and deep water masses: A qualitative comparison of southwest Pacific, southern and Atlantic Oceans. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 131, 287–302. [Google Scholar] [CrossRef]
- Majewski, W.; Olempska, E. Recent ostracods from Admiralty Bay, King George Island, West Antarctica. Pol. Polar Res. 2005, 26, 13–36. [Google Scholar]
- Brandt, A.; Gooday, A.; Brandão, S.; Brix, S.; Brökeland, W.; Cedhagen, T.; Choudhury, M.; Cornelius, N.; Danis, B.; De Mesel, I.; et al. First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 2007, 447, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Brandão, S.N.; Horne, D.V. The Platycopid Signal of oxygen depletion in the ocean: A critical evaluation of the evidence from modern ostracod biology, ecology and depth distribution. Paleogeogr. Paleoclimatol. Paleoecol. 2009, 283, 126–133. [Google Scholar] [CrossRef]
- Brandão, S.N.; Saeedi, H.; Brandt, A. Macroecology of Southern Ocean benthic Ostracoda (Crustacea) from the continental margin and abyss. Zool. J. Linn. Soc. 2022, 194, 226–255. [Google Scholar] [CrossRef]
- Chapmann, F. Ostracoda from elevated deposits on the slopes of Mount Erebus, between Cape Royds and Cape Barne. British Antarctic Expedition 1907–1909 under the command of Sir E. H. Shackleton. Rep. Sci. Investig. Geol. 1916, 2, 49–52. [Google Scholar]
- Chapmann, F. Ostracoda from up thrust mud above the Drygalski Glacier, southeast of Mount Larsen. British Antarctic Expedition 1907–1909 under the command of Sir E. H. Shackleton. Rep. Sci. Investig. Geol. 1916, 2, 37–40. [Google Scholar]
- Chapmann, F. Report on the Foraminifera and Ostracoda: Out of marine muds from soundings in the Ross Sea. British Antarctic Expedition 1907–1909 under the command of Sir E. H. Shackleton. Rep. Sci. Investig. Geol. 1916, 2, 53–80. [Google Scholar]
- Salvi, G.; Anderson, J.B.; Bertoli, M.; Castagno, P.; Falco, P.; Fernetti, M.; Montagna, P.; Taviani, M. Recent Ostracod Fauna of the Western Ross Sea (Antarctica): A Poorly Known Ingredient of Polar Carbonate Factories. Minerals 2022, 12, 937. [Google Scholar] [CrossRef]
- Ishman, S.E.; Szymcek, P. Foraminiferal distributions in the former Laresen-A ice shelf and Prince Gustav channel region, eastern Antarctic Peninsula margin: A baseline for Holocene paleoenvironmental change. Antar. Res. Ser. 2003, 79, 239–260. [Google Scholar] [CrossRef]
- Melis, R.; Salvi, G. Late Quaternary foraminiferal assemblages from western Ross Sea (Antarctica) in relation to the main glacial and marine lithofacies. Mar. Micropal. 2009, 70, 39–53. [Google Scholar] [CrossRef]
- Capotondi, L.; Bonomo, S.; Budillon, G.; Giordano, P.; Langone, L. Living and dead benthic foraminiferal distribution in two areas of the Ross Sea (Antarctica). Rend. Lincei Sci. Fis. Nat. 2020, 31, 1037–1053. [Google Scholar] [CrossRef]
- Bart, P.J.; Coquereau, L.; Warny, S.; Majewski, W. In situ foraminifera in grounding zone diamict: A working hypothesis. Antarct. Sci. 2016, 28, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Briggs, W.M. Ostracoda from the Pleistocene Taylor Formation, Ross Island, and the Recent of the Ross Sea and McMurdo Sound region, Antarctica. Antarct. J. USA 1978, 14, 27–29. [Google Scholar]
- Szczechura, J.; Blaszyk, J. Ostracods from the Pecten conglomerate (Pliocene) of Cockburn Island, Antarctic peninsula. Palaeontol. Pol. 1996, 68, 175–186. [Google Scholar]
- Gooday, A.J. Epifaunal and shallow infaunal foraminiferal communities at three abyssal NE Atlantic sites subject to differing phytodetritus input regimes. Deep-Sea Res. 1996, 43, 1395–1421. [Google Scholar] [CrossRef]
- Melis, R.; Capotondi, L.; Torricella, F.; Ferretti, P.; Geniram, A.; Hong, J.K.; Kuhn, G.; Khim, B.-K.; Kim, S.; Malinverno, E.; et al. Last Glacial Maximum to Holocene paleoceanography of the northwestern Ross Sea inferred from sediment core geochemistry and micropaleontology at Hallett Ridge. J. Micropal. 2021, 40, 15–35. [Google Scholar] [CrossRef]
- Griffiths, H.J.; Anker, P.; Linse, K.; Maxwell, J.; Post, A.L.; Stevens, C.; Tulaczyk, S.; Smith, J.A. Breaking All the Rules: The First Recorded Hard Substrate Sessile Benthic Community Far Beneath an Antarctic Ice Shelf. Front. Mar. Sci. 2021, 8, 642040. [Google Scholar] [CrossRef]
- Fabiano, M.; Danovaro, R. Meiofauna distribution and mesoscale variability in two sites of the Ross Sea (Antarctica) with contrasting food supply. Polar Biol. 1999, 22, 115–123. [Google Scholar] [CrossRef]
- Jansen, J.; Hill, N.; Dunstan, P.; McKinlay, J.; Sumner, M.; Post, A.; Eléaume, M.; Armand, L.; Warnock, J.; Galton-Fenzi, B.; et al. Abundance and richness of key Antarctic seafloor fauna correlates with modelled food availability. Nat. Ecol. Evol. 2018, 2, 71–80. [Google Scholar] [CrossRef]
- Wall-Palmer, D.; Jones, M.T.; Hart, M.B.; Fisher, J.K.; Smart, C.W.; Hembury, D.J.; Palmer, M.R.; Fones, G.R. Explosive volcanism as a cause for mass mortality of pteropods. Mar. Geol. 2011, 282, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Hunt, E.A.; Pakhomov, G.W.; Hosie, V.; Siegel, P.; Ward, K.B. Pteropods in Southern Ocean ecosystems. Prog. Oceanogr. 2008, 78, 193–221. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvi, G.; Melis, R.; Del Carlo, P.; Di Roberto, A. Ostracod and Foraminifer Responses to Late Pleistocene–Holocene Volcanic Activity in Northern Victoria Land as Recorded in Ross Sea (Antarctica) Marine Sediments. Geosciences 2023, 13, 35. https://doi.org/10.3390/geosciences13020035
Salvi G, Melis R, Del Carlo P, Di Roberto A. Ostracod and Foraminifer Responses to Late Pleistocene–Holocene Volcanic Activity in Northern Victoria Land as Recorded in Ross Sea (Antarctica) Marine Sediments. Geosciences. 2023; 13(2):35. https://doi.org/10.3390/geosciences13020035
Chicago/Turabian StyleSalvi, Gianguido, Romana Melis, Paola Del Carlo, and Alessio Di Roberto. 2023. "Ostracod and Foraminifer Responses to Late Pleistocene–Holocene Volcanic Activity in Northern Victoria Land as Recorded in Ross Sea (Antarctica) Marine Sediments" Geosciences 13, no. 2: 35. https://doi.org/10.3390/geosciences13020035
APA StyleSalvi, G., Melis, R., Del Carlo, P., & Di Roberto, A. (2023). Ostracod and Foraminifer Responses to Late Pleistocene–Holocene Volcanic Activity in Northern Victoria Land as Recorded in Ross Sea (Antarctica) Marine Sediments. Geosciences, 13(2), 35. https://doi.org/10.3390/geosciences13020035