
The Relative Stability of Planktic Foraminifer Thermal Preferences over the Past 3 Million Years

 ,
,  , , , , and
, , , , and 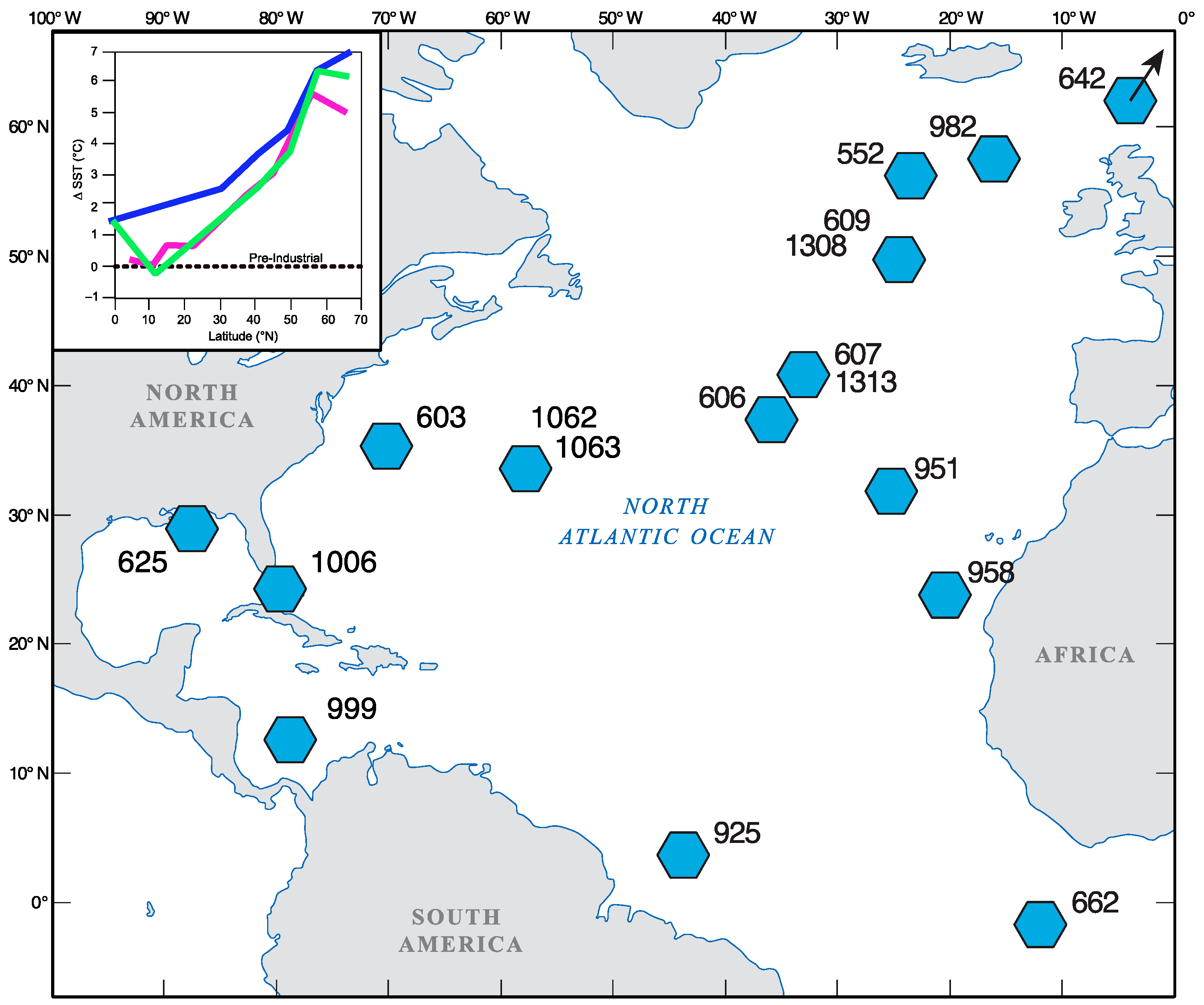{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Geographic Distribution of Pliocene Samples
2.2. Faunal Census Data
2.3. Sea Surface Temperature
2.4. Chronology
3. Results
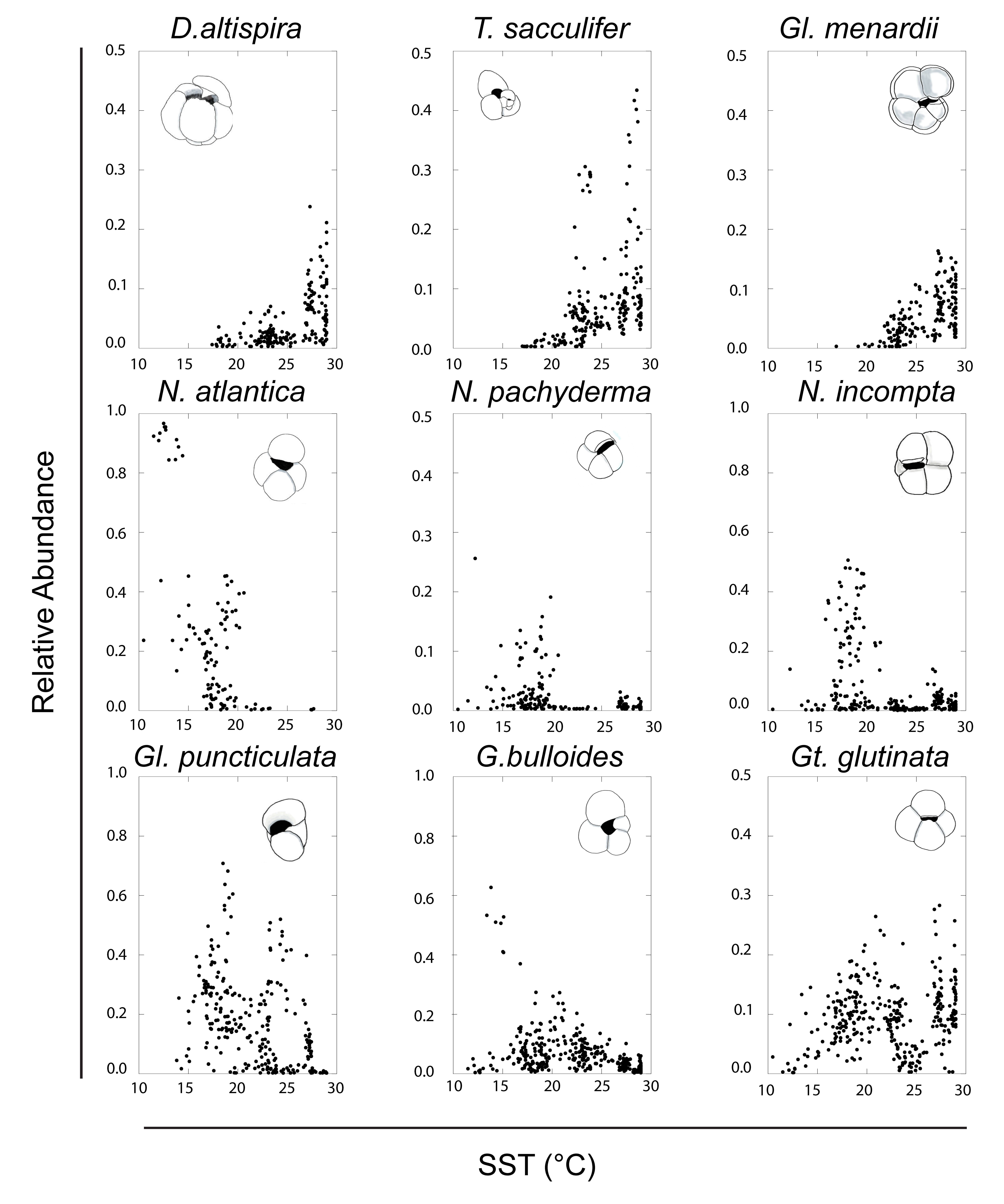
3.1. Temperature Response of Pliocene Species
3.2. Temperature Response of Pre-Industrial Core-Top Species
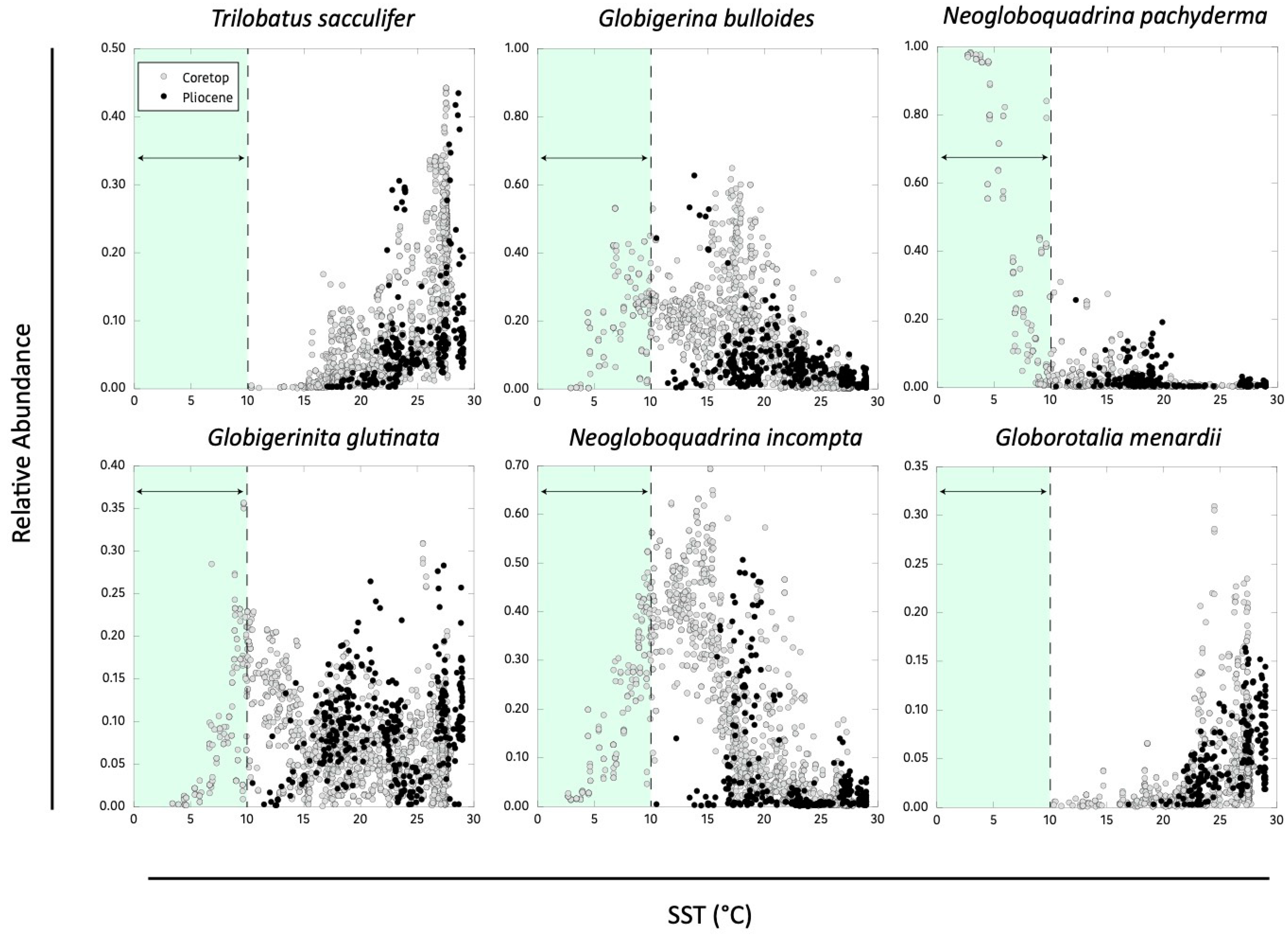
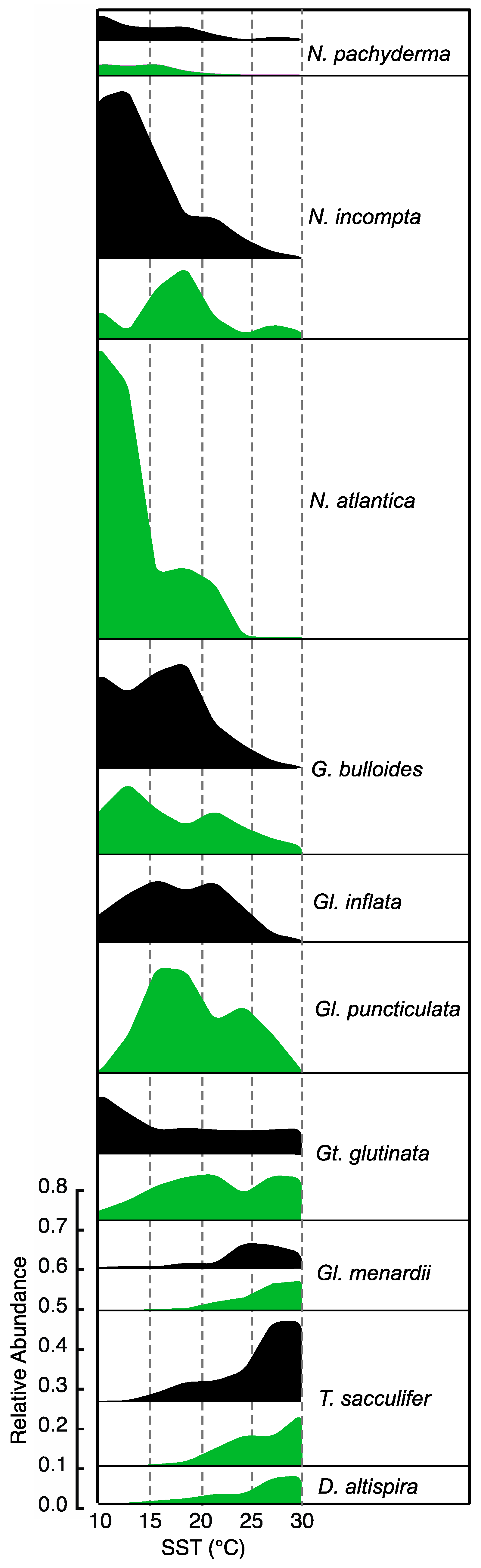
3.3. Comparison of Pliocene and PI Temperature Responses
3.3.1. Extant Taxa
3.3.2. Extinct Taxa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hutton, J. Theory of the Earth; or an investigation of the laws observable in the composition, dissolution, and restoration of land upon the Globe. Trans. R. Soc. Edinb. 1788, 1, 209–304. [Google Scholar] [CrossRef] [Green Version]
- Lyell, C. Principles of Geology, Being an Attempt to Explain the Former Changes of the Earth’s Surface, by Reference to Causes Now in Operation; John Murray: London, UK, 1830; Volume 1. [Google Scholar]
- Giamali, C.; Kontakiotis, G.; Koskeridou, E.; Ioakim, C.; Antonarakou, A. Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean). J. Mar. Sci. Eng. 2020, 8, 709. [Google Scholar] [CrossRef]
- Bé, A.W.H. An ecological, zoogeographic and taxonomic review of recent planktonic foraminifera. In Oceanic Micropaleontology; Ramsay, A.T.S., Ed.; Academic Press: London, UK, 1977; Volume 1, pp. 1–100. [Google Scholar]
- Schiebel, R.; Hemleben, C. Planktic Foraminifers in the Modern Ocean; Springer: Berlin, Germanny, 2017; p. 358. [Google Scholar]
- Morey, A.E.; Mix, A.C.; Pisias, N.G. Planktonic foraminiferal assemblages preserved in surface sediments correspond to multiple environment variables. Quat. Sci. Rev. 2005, 24, 925–950. [Google Scholar] [CrossRef]
- Bé, A.W.H.; Tolderlund, D.S. Distribution and ecology of living planktonic foraminifera in surface waters of the Atlantic and Indian Oceans. In The Micropaleontology of Oceans; Funnel, B.M., Riedel, W.R., Eds.; Cambridge University Press: London, UK, 1971; pp. 105–149. [Google Scholar]
- Tolderlund, D.S.; Bé, A.W.H. Seasonal distribution of planktonic foraminifera in the Western North Atlantic. Micropaleontology 1971, 17, 297–329. [Google Scholar]
- Imbrie, J.; Kipp, N.G. A New Micropaleontological method for paleoclimatology: Application to a Late Pleistocene Caribbean core. In The Late Cenozoic Glacial Ages; Turekian, K.K., Ed.; Yale University Press: New Haven, CT, USA, 1971; pp. 71–181. [Google Scholar]
- Cline, R.M.; Hays, J.D. Investigation of Late Quaternary Paleoceanography and Paleoclimatology; Geological Society of America: Boulder, CO, USA, 1976; Volume 123, p. 464. [Google Scholar]
- Cline, R.M.L.; Hays, J.D.; Prell, W.L.; Ruddiman, W.F.; Moore, T.C.; Kipp, N.G.; Molfino, B.E.; Denton, G.H.; Hughes, T.J.; Balsam, W.L.; et al. The last interglacial ocean. Quat. Res. 1984, 21, 123–224. [Google Scholar] [CrossRef]
- Hutson, W.H. The Agulhas Current during the Late Pleistocene: Analysis of Modern Faunal Analogs. Science 1980, 207, 64–66. [Google Scholar] [CrossRef]
- Malmgren, B.A.; Nordlund, U. Application of artificial neural networks to paleoceanographic data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 136, 359–373. [Google Scholar] [CrossRef]
- Malmgren, B.A.; Kucera, M.; Nyberg, J.; Waelbroeck, C. Comparison of statistical and artificial neural network techniques for estimating past sea surface temperatures from planktonic foraminifer census data. Paleoceanography 2001, 16, 520–530. [Google Scholar] [CrossRef]
- MARGO Project Members. Constraints on the magnitude and patterns of ocean cooling at the Last Glacial Maximum. Nat. Geosci. 2009, 2, 127–132. [Google Scholar] [CrossRef]
- CLIMAP. Seasonal Reconstructions of the Earth’s Surface at the Last Glacial Maximum; Geological Society of America: Boulder, CO, USA, 1981; p. 18. [Google Scholar]
- Sarnthein, M.; Gersonde, R.; Niebler, S.; Pflaumann, U.; Spielhagen, R.; Thiede, J.; Wefer, G.; Weinelt, M. Overview of Glacial Atlantic Ocean Mapping (GLAMAP 2000). Paleoceanography 2003, 18, 1030. [Google Scholar] [CrossRef] [Green Version]
- Antell, G.S.; Fenton, I.S.; Valdes, P.J.; Saupe, E.E. Thermal niches of planktonic foraminifera are static throughout glacial–interglacial climate change. Proc. Natl. Acad. Sci. USA 2021, 118, e2017105118. [Google Scholar] [CrossRef]
- Kucera, M.; Schonfeld, J. The origin of modern oceanic formainiferal faunas and Neogene climate change. In Deep-Time Perspectives on Climate Change: Marrying the Signal from Computer Models and Biological Proxies; The Micropalaeontological Society: London, UK, 2007; pp. 409–425. [Google Scholar]
- Keigwin, L.D. Late Cenozoic planktonic foraminiferal biostratigraphy and paleoceanography of the Panama Basin. Micropaleontology 1976, 22, 419–442. [Google Scholar]
- Thunell, R.C. Pliocene—Pleistocene paleotemperature and paleosalinity history of the Mediterranean Sea: Results from DSDP Sites 125 and 132. Mar. Micropaleontol. 1979, 4, 173–187. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Poore, R.Z. Pliocene sea surface temperatures of the North Atlantic Ocean at 3.0 Ma. Quat. Sci. Rev. 1991, 10, 189–204. [Google Scholar] [CrossRef]
- Sabaa, A.T.; Sikes, E.L.; Hayward, B.W.; Howard, W.R. Pliocene sea surface temperature changes in ODP Site 1125, Chatham Rise, east of New Zealand. Mar. Geol. 2004, 205, 113–125. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Robinson, M.M. Application of the modern analogue technique (MAT) of sea surface temperature estimation to middle Pliocene North Pacific planktic foraminifer assemblages. Palaeontol. Electron. 1998, 1, 22. [Google Scholar]
- Andersson, C. Transfer function vs. modern analog technique for estimating Pliocene sea-surface temperatures based on planktic foraminiferal data, western Equatorial Pacific Ocean. J. Foraminifer. Res. 1997, 27, 123–132. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Poore, R.Z. A new planktic foraminifer transfer function for estimating Pliocene—Holocene paleoceanographic conditions in the North Atlantic. Mar. Micropaleontol. 1990, 16, 1–23. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Foley, K.M.; Stoll, D.K.; Chandler, M.A.; Sohl, L.E.; Bentsen, M.; Otto-Bliesner, B.L.; Bragg, F.J.; Chan, W.-L.; Contoux, C.; et al. Sea Surface Temperature of the mid-Piacenzian Ocean: A Data-Model Comparison. Sci. Rep. 2013, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.; Barron, J.; Poore, R. Middle Pliocene sea surface temperatures: A global reconstruction. Mar. Micropaleontol. 1996, 27, 13–25. [Google Scholar]
- Dowsett, H.J. The PRISM palaeoclimate reconstruction and Pliocene sea-surface temperature. In Deep-Time Perspectives on Climate Change: Marrying the Signal from Computer Models and Biological Proxies; Williams, M., Haywood, A.M., Gregory, J., Schmidt, D.N., Eds.; Micropalaeontological Society (Special Publication), Geological Society of London: London, UK, 2007; pp. 459–480. [Google Scholar]
- Dowsett, H.; Robinson, M.; Haywood, A.; Salzmann, U.; Hill, D.; Sohl, L.; Chandler, M.; Williams, M.; Foley, K.; Stoll, D. The PRISM3D paleoenvironmental reconstruction. Stratigraphy 2010, 7, 123–139. [Google Scholar]
- Dowsett, H.J.; Chandler, M.A.; Cronin, T.M.; Dwyer, G.S. Middle Pliocene sea surface temperature variability. Paleoceanography 2005, 20, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.; Dolan, A.; Rowley, D.; Moucha, R.; Forte, A.M.; Mitrovica, J.X.; Pound, M.; Salzmann, U.; Robinson, M.; Chandler, M.; et al. The PRISM4 (mid-Piacenzian) paleoenvironmental reconstruction. Clim. Past 2016, 12, 1519–1538. [Google Scholar] [CrossRef] [Green Version]
- Chandler, M.A.; Rind, D.; Thompson, R. Joint investigations of the middle Pliocene climate II: GISS GCM Northern Hemisphere results. Glob. Planet. Change 1994, 9, 197–219. [Google Scholar] [CrossRef]
- Sloan, L.C.; Crowley, T.J.; Pollard, D. Modeling of middle Pliocene climate with the NCAR GENESIS general circulation model. Mar. Micropaleontol. 1996, 27, 51–61. [Google Scholar] [CrossRef]
- Haywood, A.M.; Valdes, P.J.; Sellwood, B.W. Global scale palaeoclimate reconstruction of the middle Pliocene climate using the UKMO GCM: Initial results. Glob. Planet. Chang. 2000, 25, 239–256. [Google Scholar] [CrossRef]
- Haywood, A.; Valdes, P. Modelling Pliocene warmth: Contribution of atmosphere, oceans and cryosphere. Earth Planet. Sci. Lett. 2004, 218, 363–377. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Robinson, M.M.; Haywood, A.M.; Hill, D.J.; Dolan, A.M.; Stoll, D.K.; Chan, W.-L.; Abe-Ouchi, A.; Chandler, M.A.; Rosenbloom, N.A.; et al. Assessing confidence in Pliocene sea surface temperatures to evaluate predictive models. Nat. Clim. Chang. 2012, 2, 365–371. [Google Scholar] [CrossRef]
- Haywood, A.M.; Hill, D.J.; Dolan, A.M.; Otto-Bliesner, B.L.; Bragg, F.; Chan, W.L.; Chandler, M.A.; Contoux, C.; Dowsett, H.J.; Jost, A.; et al. Large-scale features of Pliocene climate: Results from the Pliocene Model Intercomparison Project. Clim. Past 2013, 9, 191–209. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.; Robinson, M.; Foley, K. A global planktic foraminifer census data set for the Pliocene ocean. Sci. Data 2015, 2, 150076. [Google Scholar] [CrossRef]
- Kipp, N.G. New transfer function for estimating past sea-surface conditions from sea-bed distribution of planktonic formainiferal asemblages in the North Atlantic: Investigations of late Quaternary Paleoceanography and Paleoclimatology. Geol. Soc. Am. Mem. 1976, 145, 3–41. [Google Scholar]
- Dowsett, H.J.; Cronin, T.M.; Poore, R.Z.; Thompson, R.S.; Whatley, R.C.; Wood, A.M. Micropaleontological Evidence for Increased Meridional Heat Transport in the North Atlantic Ocean During the Pliocene. Science 1992, 258, 1133–1135. [Google Scholar] [CrossRef] [PubMed]
- Dowsett, H.J.; Robinson, M.M.; Foley, K.M.; Herbert, T.D.; Otto-Bliesner, B.L.; Spivey, W. The mid-Piacenzian of the North Atlantic Ocean. Stratigraphy 2019, 16, 119–144. [Google Scholar] [CrossRef]
- McClymont, E.L.; Ford, H.L.; Ho, S.L.; Tindall, J.C.; Haywood, A.M.; Alonso-Garcia, M.; Bailey, I.; Berke, M.A.; Littler, K.; Patterson, M.O.; et al. Lessons from a high-CO2 world: An ocean view from ~3 million years ago. Clim. Past 2020, 16, 1599–1615. [Google Scholar] [CrossRef]
- Müller, P.J.; Kirst, G.; Ruhland, G.; von Storch, I.; Rosell-MelÈ, A. Calibration of the alkenone paleotemperature index U37K’ based on core-tops from the eastern South Atlantic and the global ocean (60°N–60°S). Geochim. Et Cosmochim. Acta 1998, 62, 1757–1772. [Google Scholar] [CrossRef]
- Siccha, M.; Kucera, M. ForCenS, a curated database of planktonic foraminifera census counts in marine surface sediment samples. Sci. Data 2017, 4, 170109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shea, D.; Hurrell, J.; Phillips, A. Merged Hadley-OI Sea Surface Temperature and Sea Ice Concentration Data Set; Version 5.0; NCAR: Boulder, CO, USA, 2022. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Foley, K.M.; Robinson, M.M.; Herbert, T.D. PRISM late Pliocene (Piacenzian) alkenone—derived SST data. In U.S. Geological Survey Data Release; USGS: Reston, VA, USA, 2017. [Google Scholar] [CrossRef]
- Bachem, P.E.; Risebrobakken, B.; McClymont, E.L. Sea surface temperature variability in the Norwegian Sea during the late Pliocene linked to subpolar gyre strength and radiative forcing. Earth Planet. Sci. Lett. 2016, 446, 113–122. [Google Scholar]
- Herbert, T.D. 6.15—Alkenone Paleotemperature Determinations. In Treatise on Geochemistry; Holland, H.D., Turekian, K.K., Eds.; Pergamon: Oxford, UK, 2003; pp. 391–432. [Google Scholar]
- Lawrence, K.T.; Herbert, T.D.; Dekens, P.S.; Ravelo, A.C. The application of the alkenone organic proxy to the study of Plio-Pleistocene climate. In Deep-Time Perspectives on Climate Change: Marrying the Signal from Computer Models and Biological Proxies; Williams, M., Haywood, A.M., Gregory, F.J., Schmidt, D.N., Eds.; Micropalaeontological Society (Special Publication), Geological Society of London: London, UK, 2007; pp. 539–562. [Google Scholar]
- Foley, K.M.; Dowsett, H.J. Community sourced mid-Piacenzian sea surface temperature (SST) data. U.S. Geol. Surv. Data Release 2019. [Google Scholar] [CrossRef]
- Berggren, W.A. Cenozoic biostratigraphy and paleobiogeography of the North Atlantic. Initial Rep. Deep Sea Drill. Proj. 1972, 12, 965–1001. [Google Scholar]
- Poore, R.Z. Oligocene through Quaternary planktonic foraminiferal biostratigraphy of the North Atlantic: DSDP Leg 49. Initial Rep. Deep Sea Drill. Proj. 1978, 49, 447–517. [Google Scholar] [CrossRef]
- Weaver, P.P.E. Late Miocene-Recent planktonic foraminifers from the North Atlantic: Deep Sea Drilling Project Leg 94. Initial Rep. Deep Sea Drill. Proj. 1987, 94, 703–727. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Robinson, M.M. Mid-Pliocene planktic foraminifer assemblage of the North Atlantic Ocean. Micropaleontology 2007, 53, 105–126. [Google Scholar] [CrossRef]
- Flower, B.P. Data Report: Planktonic foraminifers from the subpolar North Atlantic and Nordic Seas: Sites 980–987 and 907. Proc. Ocean Drill. Program 1999, 162, 19–34. [Google Scholar] [CrossRef]
- Rillo, M.C.; Woolley, S.; Hillebrand, H. Drivers of global pre-industrial patterns of species turnover in planktonic foraminifera. Ecography 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Poore, R.Z.; Steinmetz, J.C.; Schrader, H.J. Biostratigraphic summary of DSDP Leg 49. Initial Rep. Deep Sea Drill. Proj. 1978, 49, 851–858. [Google Scholar] [CrossRef]
- Weaver, P.P.E.; Clement, B.M. Magnetobiostratigraphy of planktonic foraminiferal diatoms. Initial Rep. Deep Sea Drill. Proj. 1986, 94, 815–829. [Google Scholar] [CrossRef]
- Dowsett, H.J. Application of the Graphic Correlation method to Pliocene marine sequences. Mar. Micropaleontol. 1989, 14, 3–32. [Google Scholar] [CrossRef]
- Brombacher, A.; Wilson, P.A.; Bailey, I.; Ezard, T.H.G. The Dynamics of Diachronous Extinction Associated with Climatic Deterioration Near the Neogene/Quaternary Boundary. Paleoceanogr. Paleoclimatology 2021, 36, e2020PA004205. [Google Scholar] [CrossRef]
- Malmgren, B.; Kennett, J. Phyletic gradualism in a Late Cenozoic planktonic foraminiferal lineage; DSDP Site 284, southwest Pacific. Paleobiology 1981, 7, 230–240. [Google Scholar] [CrossRef]
- Norris, R.D.; Corfield, R.M.; Cartlidge, J.E. Evolution of depth ecology in the planktic foraminifera lineage Globorotalia (Fohsella). Geology 1993, 21, 975–978. [Google Scholar] [CrossRef]
- Prentice, M.L.; Bockheim, J.G.; Wilson, S.C.; Burckle, L.H.; Hodell, D.A.; Schluchter, C.; Kellogg, D.E. Late Neogene Antarctic glacial history: Evidence from Central Wright Valley. In The Antarctic Paleoenvironment: A Perspective on Global Change; Kennett, J.P., Warnke, D.A., Eds.; Antarctic Research Series American Geophysical Union: Washington, DC, USA, 1993; Volume 60, pp. 207–250. [Google Scholar]
- Savin, S.M.; Abel, L.; Barrera, E.; Hodell, D.; Kennett, J.P.; Murohy, M.; Keller, G.; Killingley, J.; Vincent, E. The evolution of Miocene surface and near-surface marine temperatures: Oxygen isotope evidence. In The Miocene Ocean: Paleoceanography and Biogeography, Geological Society of America Memoir 163; Kennett, J.P., Ed.; Geological Society of America: Boulder, CO, USA, 1985; pp. 49–82. [Google Scholar]
- Birch, H.; Coxall, H.K.; Pearson, P.N.; Kroon, D.; O’Regan, M. Planktonic foraminifera stable isotopes and water column structure: Disentangling ecological signals. Mar. Micropaleontol. 2013, 101, 127–145. [Google Scholar] [CrossRef]
- Spiegler, D.; Jansen, E. Planktonic Foraminifer Biostratigraphy of Norwegian Sea Sediments: ODP Leg 104. Ocean Drill. Program Sci. Results 1989, 104, 681–696. [Google Scholar] [CrossRef]
- Poore, R.Z. Temporal and spatial distribution of ice-rafted mineral grains in Pliocene sediments of the North Atlantic: Implications for Late Cenozoic climatic history. In The Deep Sea Drilling Project: A Decade of Progress; Warme, J.E., Douglas, R.G., Winterer, E.L., Eds.; Society of Economic Paleontologists and Minerologists Special Publication: Tulsa, OK, USA, 1981; Volume 32, pp. 505–515. [Google Scholar]
- Poore, R.Z.; Berggren, W.A. The morphology and classification of Neogloboquadrina atlantica (Berggren). J. Foraminifer. Res. 1975, 5, 76–84. [Google Scholar] [CrossRef]
- Levitus, S.; Boyer, T.P. World ocean atlas 1994 (vol. 4): Temperature. In NOAA Atlas NESDIS; US Department of Commerce: Washington, DC, USA, 1994; Volume 4. [Google Scholar]
- Bova, S.; Rosenthal, Y.; Liu, Z.; Godad, S.P.; Yan, M. Seasonal origin of the thermal maxima at the Holocene and the last interglacial. Nature 2021, 589, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.M.; Dowsett, H.J.; Dwyer, G.S.; Lawrence, K.T. Reevaluation of mid-Pliocene North Atlantic sea surface temperatures. Paleoceanography 2008, 23, PA3213. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, K.T.; Sigman, D.M.; Herbert, T.D.; Riihimaki, C.A.; Bolton, C.T.; Martinez-Garcia, A.; Rosell-Mele, A.; Haug, G.H. Time-transgressive North Atlantic productivity changes upon Northern Hemisphere glaciation. Paleoceanography 2013, 28, 740–751. [Google Scholar] [CrossRef] [Green Version]
- Naafs, B.D.A.; Voelker, A.H.L.; Karas, C.; Andersen, N.; Sierro, F.J. Repeated Near-Collapse of the Pliocene Sea Surface Temperature Gradient in the North Atlantic. Paleoceanogr. Paleoclimatol. 2020, 35, e2020PA003905. [Google Scholar] [CrossRef]
- Soltwedel, T.; Hasemann, C.; Vedenin, A.; Bergmann, M.; Taylor, J.; Krauß, F. Bioturbation rates in the deep Fram Strait: Results from in situ experiments at the arctic LTER observatory HAUSGARTEN. J. Exp. Mar. Biol. Ecol. 2019, 511, 1–9. [Google Scholar] [CrossRef]
- Hülse, D.; Vervoort, P.; van de Velde, S.J.; Kanzaki, Y.; Boudreau, B.; Arndt, S.; Bottjer, D.J.; Hoogakker, B.; Kuderer, M.; Middelburg, J.J.; et al. Assessing the impact of bioturbation on sedimentary isotopic records through numerical models. Earth-Sci. Rev. 2022, 234, 104213. [Google Scholar] [CrossRef]
- Hemleben, C.; Spindler, M.; Anderson, O.R. Modern Planktonic Foraminifera; Springer: New York, NY, USA, 1989; p. 363. [Google Scholar]
- Strack, A.; Jonkers, L.C.; Rillo, M.; Hillebrand, H.; Kucera, M. Plankton response to global warming is characterized by non-uniform shifts in assemblage composition since the last ice age. Nat. Ecol. Evol. 2022, 6, 1871–1880. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dowsett, H.; Robinson, M.; Foley, K.; Herbert, T.; Hunter, S.; Andersson, C.; Spivey, W. The Relative Stability of Planktic Foraminifer Thermal Preferences over the Past 3 Million Years. Geosciences 2023, 13, 71. https://doi.org/10.3390/geosciences13030071
Dowsett H, Robinson M, Foley K, Herbert T, Hunter S, Andersson C, Spivey W. The Relative Stability of Planktic Foraminifer Thermal Preferences over the Past 3 Million Years. Geosciences. 2023; 13(3):71. https://doi.org/10.3390/geosciences13030071
Chicago/Turabian StyleDowsett, Harry, Marci Robinson, Kevin Foley, Timothy Herbert, Stephen Hunter, Carin Andersson, and Whittney Spivey. 2023. "The Relative Stability of Planktic Foraminifer Thermal Preferences over the Past 3 Million Years" Geosciences 13, no. 3: 71. https://doi.org/10.3390/geosciences13030071
APA StyleDowsett, H., Robinson, M., Foley, K., Herbert, T., Hunter, S., Andersson, C., & Spivey, W. (2023). The Relative Stability of Planktic Foraminifer Thermal Preferences over the Past 3 Million Years. Geosciences, 13(3), 71. https://doi.org/10.3390/geosciences13030071