Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review



Abstract
:1. Introduction
- To characterize the main parameters of the large lakes and estimate their reference conditions on the basis of a time-space analysis of dominant characteristics;
- To identify the main patterns of changes in the aquatic ecosystems of the lakes under the conditions of anthropogenic loads and their reduction, from background characteristics through degradation to recovery;
- To explain the course of these changes from the perspective of ecological theory and to assess the possibility of ecosystems returning to their natural state after toxic stress.
2. Main Features of the Natural Condition of the Lakes
3. Anthropogenic Loads in the Bay
4. Through Disturbance to Recovery
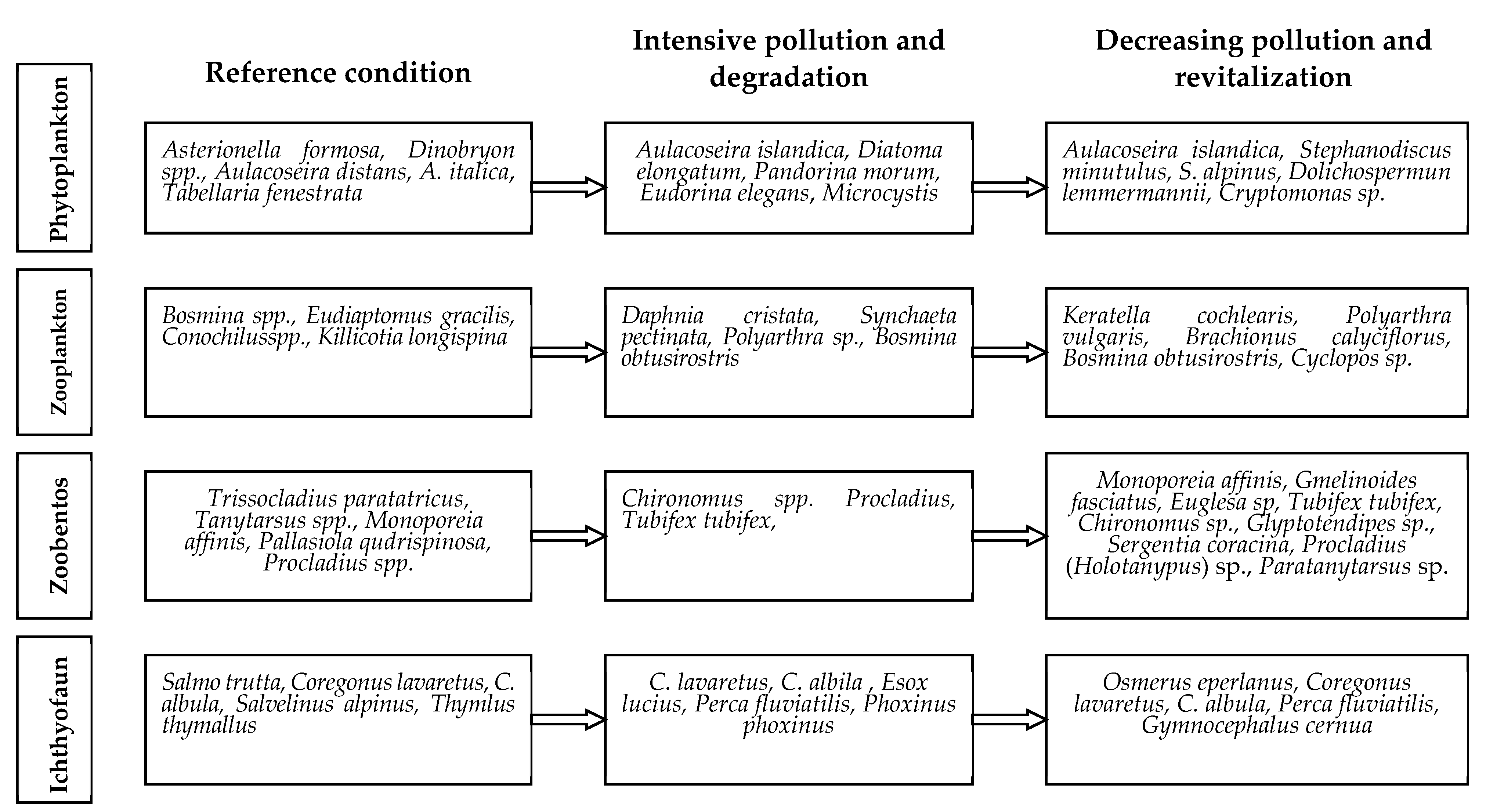
4.1. Pollution-Caused Changes
4.2. Tendency to Recovery
5. Discussion: Is it Possible to Recover the Lakes?
6. Conclusions
Funding
Conflicts of Interest
References
- Butler, G.C. Development in Ecotoxicology. Ecol. Bull. 1984, 36, 9–12. [Google Scholar]
- Cairns, J., Jr. The predict, validation, monitoring and migration of anthropogenic effect upon natural systems. Environ. Aud. 1990, 2, 19–25. [Google Scholar]
- Ecotoxicology: A Hierarchical Treatment; Newman, M.C.; Jagoe, C.H. (Eds.) Levis Publishers Ltd.: Boca Raton, FL, USA, 1996. [Google Scholar]
- Thruhaut, R. Ecotoxicology: Objectives, Principles and perspectives. Ecotoxicol. Environ. Saf. 1977, 1, 151–173. [Google Scholar] [CrossRef]
- Walker, C.H.; Hopkin, S.P.; Sibly, R.M.; Peakall, D.B. Principles of Ecotoxicology; Taylor & Francis Ltd.: London, UK, 2001; 307p. [Google Scholar]
- Chapman, M.G. Improving sampling designs for measuring restoration in aquatic habitats. J. Aquat. Ecosyst. Strees Recovery 1999, 6, 235–251. [Google Scholar] [CrossRef]
- Power, M. Recovery in aquatic ecosystem: An overview of knowledge and needs. J. Aquat. Ecosyst. Stress Recovery 1999, 6, 253–257. [Google Scholar] [CrossRef]
- Klotzi, F.; Gootjans, A.P. Restoration of natural and semi-natural wetland systems in Central Europe: Progress and predictability of developments. Restor. Ecol. 2001, 2, 209–219. [Google Scholar] [CrossRef]
- Cairns, J., Jr. Restoration Ecology and Ecotoxicology. In Handbook of Ecotoxicology; Hoffman, D.J., Rattner, B.A., Burton, G.A., Cairnce, J., Jr., Eds.; Lewis Publishers: Boca Raton, FL, USA, 2005; pp. 1015–1029. [Google Scholar]
- Falk, D.A.; Palmer, M.; Zedler, J. Foundation of Restoration Ecology: The Science and Practice of Ecological Restoration; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Hobbs, R.J. Setting effective and realistic restoration goals: Key direction for researches. Restor. Ecol. 2007, 5, 354–357. [Google Scholar] [CrossRef]
- Özkan, K.; Jeppesen, E.; Davidson, T.A.; Bjerring, R.; Johansson, L.S.; Søndergaard, M.; Lauridsen, T.L.; Svenning, J.-C. Long-Term Trends and Temporal Synchrony in Plankton Richness, Diversity and Biomass Driven by Re-Oligotrophication and Climate across 17 Danish Lakes. Water 2016, 8, 427. [Google Scholar] [CrossRef]
- Aleksandrov, B.M. About studying of structure of bottom fauna Onega Lake. In Preliminary Results of Works of Complex Expedition on Research Onega Lake; Petrozavodsk: Karelia, Russia, 1968; pp. 37–39. (In Russian) [Google Scholar]
- Berg, L.S.; Pravdin, I.F. Fishes of Kola Peninsula. Lenigrad, News Vniorkh 1948, 26, 37–45. (In Russian) [Google Scholar]
- Gerd, S.V. Benthos Biocenoses of Large Lakes of Karelia; Izd-vo Karelo-Finskogo gosuniversiteta: Petrozavodsk, Russia, 1949. (In Russian) [Google Scholar]
- Poretskij, V.S.; Zhuze, A.P.; Sheshukova, V.S. Diatoms of Kola Peninsula in collection with microscopic structure Kola Diatomite. Work. Geomorphol. Inst. Ussr 1934, 8, 96–210. (In Russian) [Google Scholar]
- Sokolov, I.I. Zoobentos of Littoral Zone of Southern Half of Ladoga Lake. In Works of Karelian Department AS SSSR; Karelian department AS SSSR: Petrozavodsk, Russia, 1956; Volume 5, pp. 76–87. (In Russian) [Google Scholar]
- Voronikhin, N.N. Seaweed and their groupings in lakes Imandra and Notozero (Kola Peninsula). In Works of Botanical Institute AN USSR; Series 2, Sporous Plants; Botanical Institute AN USSR: Moscow, Russia, 1935; pp. 107–150. (In Russian) [Google Scholar]
- Anthropogenic Eutrophycation of Ladoga Lake; Petrova, N.A. (Ed.) Nauka: Leningrad, Russia, 1982. (In Russian) [Google Scholar]
- Anthropogenic Modifications of the Lake Imandra Ecosystem; Moiseenko, T.I. (Ed.) Nauka: Moscow, Russia, 2002. (In Russian) [Google Scholar]
- Rukhovets, L.; Filatov, N. Ladoga and Onego—Great European Lakes: Observations and Modeling; Rukhovets, L., Filatov, N., Eds.; Springer: Chichester, UK, 2010. [Google Scholar]
- Ladoga; Rumyantsev, V.A.; Kondratev, S.A. (Eds.) Nestor-istoriya: St. Petersburg, Russia, 2013. (In Russian) [Google Scholar]
- Filatov, N.N.; Kalinkina, N.M.; Tekanova, E.V. Modern changes in the ecosystem of Lake Onego with climate warming. Limnol. Freshwater Biol. 2018, 1, 15–17. [Google Scholar] [CrossRef]
- Kalinkina, N.; Belkina, N. Dynamics of benthic communities state and the sediment chemical composition in Lake Onega under the influence of anthropogenic and natural factors. Principy Èkologii 2018, 2, 56–74. [Google Scholar] [CrossRef]
- Terentyev, P.M.; Kashulin, N.A.; Zubova, E.M. The role of european smelt Osmerus eperlanus in fish fauna structure of the Imandra lake basin (Murmansk province). Trudy Zool. Inst. RAN. 2017, 2, 228–243. [Google Scholar]
- Eaton, A.; Arnold, E.; Archie, A.E.; Rice, E.W.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater, 17th ed.; American Public Health Association (APHA): Washington, DC, USA, 1992. [Google Scholar]
- Abakumov, V.A. (Ed.) Guidance on Hydrobiological Monitoring of Freshwater Ecosystems; Gidrometeoizdat: St. Petersburg, Russia, 1992; p. 318. [Google Scholar]
- Onezhskoe ozero. Atlas (Lake Onega: Atlas); Filatov, N.N. (Ed.) Izd. Kar. NTs RAN: Petrozavodsk, Russia, 2010. [Google Scholar]
- Petrova, N.A. The phytoplankton of Ladoga and Onega lakes and its recent successional changes. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1987, 25, 11–18. [Google Scholar]
- Krokhin, E.M.; Semenovich, N.I. Data on Water Bodies in the Kola Peninsula. In The Collection NO 1, Manuscript. Funds of Kola Science Centre of RAN; Kola Science Centre of RAN: Apatity, Russia, 1940. [Google Scholar]
- Lake Onega. Ecological Problems; Karelian Research Centre: Petrozavodsk, Russia, 1999. (In Russian)
- Sokolova, M.F. Zooplankton of Ladoga Lake. Proc. VNIORH 1956, 38, 53–65. (In Russian) [Google Scholar]
- Slepukhina, T.D. Features of Development Macrozoobentos in Different lake Zones. Ladoga Lake—Criteria of Ecosystem Condition; Nauka: Sank-Petersburg, Russia, 1992; p. 218. (In Russian) [Google Scholar]
- Poliakova, T.N. Bottom cenosis in conditions anthropogenic eutrophication. In Lake Onega: Ecological Problems; Filatov, N.N., Ed.; Karelian Research Centre: Petrozavodsk, Russia, 1999; Volume 83, pp. 211–227. (In Russian) [Google Scholar]
- Lake Ladoga—Past, Present and Future; Rumyntsev, V.A.; Drabkova, V.G. (Eds.) Nauka: Sankt-Petersburg, Russia, 2002. (In Russian) [Google Scholar]
- Sabylina, A.V.; Lozovik, P.A.; Zobkov, M.B. Water chemistry in Onega Lake and its tributaries. Water Resour. 2010, 37, 842–853. [Google Scholar] [CrossRef]
- Vollenweider, R.A. Advances in defining critical loading levels for phosphorous in lake eutrophication. Met. Ins. Ital. Jdrobion 1979, 33, 53–83. [Google Scholar]
- Belkina, N.A.; Polyakova, T.N.; Timakova, T.M.; Kalinkina, N.M. The state of sediments as a consequence of anthropogenic influence on Lake Onego. In Proceeding of the Fourth International Lake Ladoga Symposium 2002; Publications of Karelian Institute: Joensuu, Finland, 2003; Volume 138, pp. 277–283. [Google Scholar]
- Moiseenko, T.I.; Yakovlev, V.A. Anthropogenic Transformations of Aquatic Ecosystems in the Kola North; Rumiancev, V.A., Ed.; Nauka: Leningrad, Russia, 1990. (In Russian) [Google Scholar]
- Arshanitsa, N.M.; Foresters, L.A. Patho-morphological analysis of the fish status in the field and experimental toxicology studies. Methods Ichthyol. Res. 1987, 7–9. [Google Scholar]
- Moiseenko, T.I.; Kudrjavzeva, L.P. Trace Metals Accumulation and Fish pathologies in Areas affected by Mining and Metallurgical enterprises. Environ. Pollut. 2002, 114, 285–297. [Google Scholar] [CrossRef]
- Adam, S.M.; Ryon, M.G.A. Comparison of health assessment approaches for evaluating the effects of contaminant-related stress on fish populations. J. Aquat. Ecosyst. Health 1994, 3, 15–25. [Google Scholar]
- Sandimirov, S.S.; Kudryavceva, L.P.; Petrova, O.V. The current state of hydro-chemical indicators of the southern reaches of lake Imandra. Voda Him. I Ekol. 2017, 2, 9–19. [Google Scholar]
- Terentyeva, I.A.; Kashulin, N.A.; Denisov, D.B. Estimate of the trophic status of subarctic Lake Imandra. Vestn. Mgtu. 2017, 20, 197–204. [Google Scholar] [CrossRef]
- Suding, K.N.; Gross, K.L.; Houseman, G.R. Alternative states and positive feedbacks in restoration ecology. Trends Ecol. Evol. 2004, 19, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.A.; Hobbs, R.J.; Higgs, E.; Aronson, J. Ecological restoration and global climate change. Restor. Ecol. 2006, 14, 170–176. [Google Scholar] [CrossRef]
- Falk, D.A.; Palmer, M.A.; Zedler, J.B. Foundations of Restoration Ecology; Island Press: Washington, DC, USA, 2013; p. 378. [Google Scholar]
- Duarte, C.M. Coastal eutrophication research: A new awareness. Hydrobiologia 2009, 629, 263–269. [Google Scholar] [CrossRef]
- Schindler, D.W.; Vallentyne, J.R. The Algal Bowl: Over fertilization of the World’s Freshwaters and Estuaries; University of Alberta Press: Edmonton, AB, Canada, 2008. [Google Scholar]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative stable states in ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Odum, E.P. Trends Expected in Stressed Ecosystems. Bioscience 1985, 35, 419–422. [Google Scholar] [CrossRef]
- The Second Roshydromet Assessment Report on Climate Change and its Consequences in the Russian Federation. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=2&ved=2ahUKEwicjIugxP3lAhXFy4sBHSaoC04QFjABegQIBRAC&url=http%3A%2F%2Fclimate2008.igce.ru%2Fv2008%2Fpdf%2Fresume_ob_eng.pdf&usg=AOvVaw07_6Bo22ZiqmnxMuW3TVRd (accessed on 22 November 2019).
- Grey, C.B.; Neilson, M.; Johannsson, O.; Fitzsimmons, J.; Millard, S.; Dermott, R. Lake Ontario. The Book of Canadian Lakes; Monograph Series; FAO: Quebec City, QC, Canada, 1994; Volume 3, pp. 14–36. [Google Scholar]
- Great Lake Ecosystem Report. 2001. Available online: http://www.epa.gov/glnpo/ rptcong. (accessed on 9 March 2006).
- Willen, E. Phytoplankton and reversed eutrophication in Lake Malaren, Central Sweden, 1965–1983. Br. Phycol. J. 1987, 22, 193–208. [Google Scholar] [CrossRef]
- Scavia, D.; Allan, J.D.; Arend, K.K.; Bartell, S.M.; Beletsky, D.; Bosch, N.S.; Brandt, S.B.; Briland, R.D.; Daloglu, I.; Depinto, J.; et al. Assessing and addressing the re-eutrophication of Lake Erie: Central basin hypoxia. J. Great Lakes Res. 2014, 40, 226–246. [Google Scholar] [CrossRef]
- Stewart, A.J.; Wetzel, R.G. Cryptophytes and other microflagellates as couplers in planktonic community dynamics. Arch. Hydrobiol 1986, 106, 1–19. [Google Scholar]
- Sutton, J.N.; André, L.; Cardinal, D.; Conley, D.J.; de Souza, G.F.; Dean, J.; Dodd, J.; Ehlert, C.; Ellwood, M.J.; Frings, P.J.; et al. A review of the stable isotope bio-geochemistry of the global silicon cycle and its associated trace elements. Front. Earth Sci. J. Artic. Rev. 2018. [Google Scholar] [CrossRef]
- Conley, D.J.; Frings, P.J.; Fontorbe, G.; Clymans, W.; Stadmark, J.; Hendry, K.R.; Marron, A.O.; De La Rocha, C.L. Biosilicification drives a decline of dissolved Si in the oceans through geologic time. Front. Mar. Sci. 2017, 4, 397. [Google Scholar] [CrossRef]
- Berezina, N.A.; Panov, B.E. Invasion of Baikal amphipod Gmelinoides fasciatus (Amphipoda, Crustacea) on Onega Lake. Zool. J. 2003, 6, 731–734. [Google Scholar]
- Berezina, N.A.; Maksimov, A.A. Abundance and preferences of Amphipods (Crustacea: Amphypoda) in the Eastern Gulf of Finland, Baltic Sea. J. Sib. Univ. Biol. 2016, 4, 409–426. [Google Scholar] [CrossRef]
- Vandysh, O.I.; Kashulin, N.A.; Cherepanov, A.A. Long-term changes in zooplankton communities of Lake Imandra under conditions of multi-level pollution by mining production. Vestn. Kola Sci. Cent. Russ. Acad. Sci. 2014, 2, 121–129. (In Russian) [Google Scholar]
- Filatov, N.N.; Kalinkina, N.M.; Kulikova, T.P.; Litvinenko, A.V.; Lozovik, P.A. The Largest Lakes are the Reservoirs of the North-West ETR: Current State and Changes in Ecosystems under Climatic and Anthropogenic Impacts Lozovik; KarRC RAS: Petrozavodsk, Russia, 2015; 375p. (In Russian) [Google Scholar]
- Moiseenko, T.I. A Fate of Metals in Arctic Surface Waters. Method for Defining Critical Levels. Sci. Total Environ. 1999, 236, 19–39. [Google Scholar] [CrossRef]
- O’Naill, R.V. Recovery in complex ecosystems. J. Aquat. Ecosyst. Stress Recovery 1999, 6, 181–187. [Google Scholar]




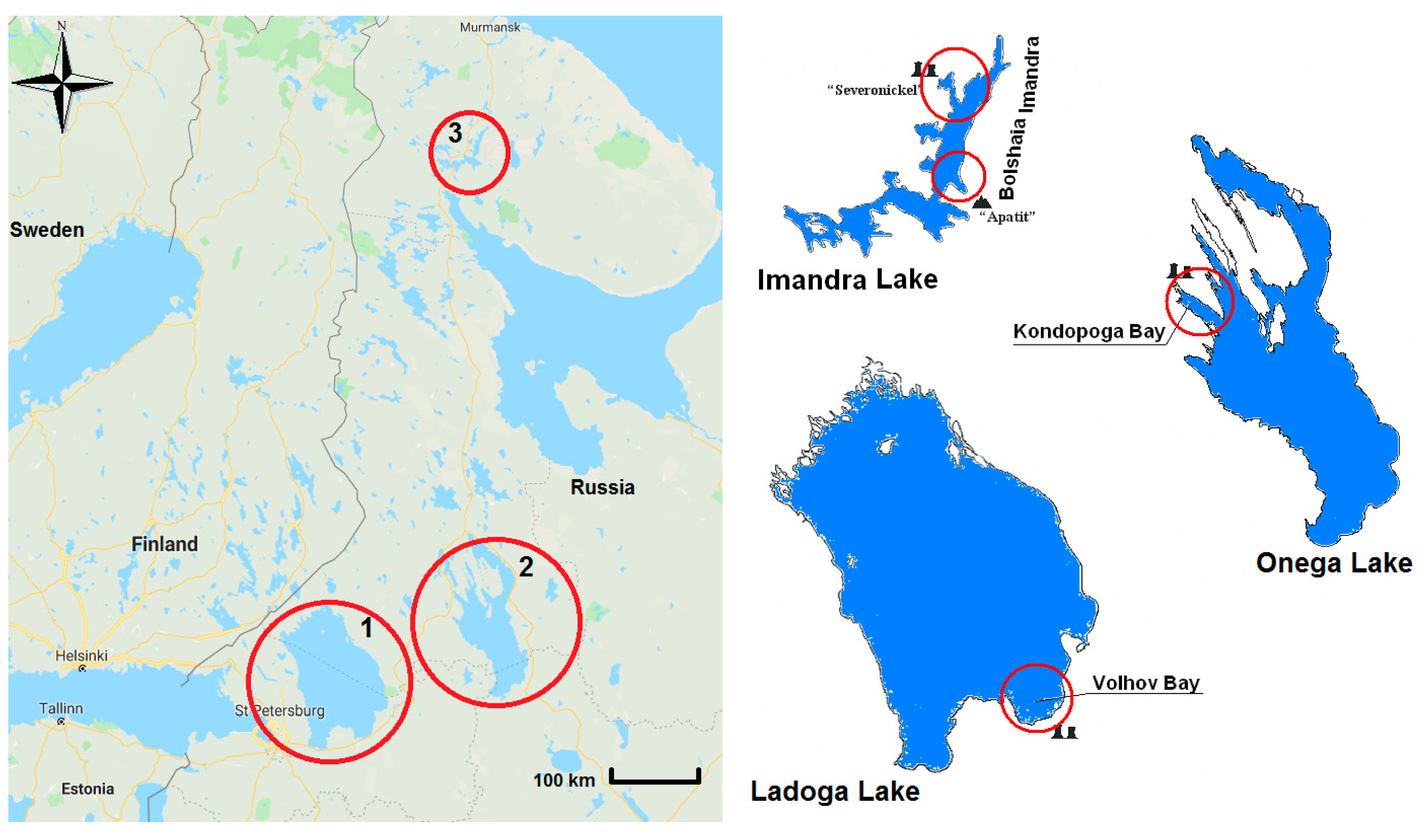{kind=link}
{kind=link}
{kind=link}
| Parameters | Lake Ladoga | Lake Onega | Lake Imandra | |||
|---|---|---|---|---|---|---|
| Whole Lake | Volkhov Bay | Whole Lake | Kondopoga Bay | Whole Lake | Bolshaya Imandra | |
| Area, km2 | 18,300 | 1464 (8%) | 9800 | 224 (2.7%) | 813 | 244 (30%) |
| Depth, m, max./average | 230/47 | 20/7 | 120/30 | 82/21 | 67/13 | 67/17 |
| Volume km3 | 838 | - | 262 | - | 11 | - |
| Water residence time, years | 11 | 3 | 15 | 5 | 6 | 4 |
| SD, m | 2.9 ± 0.9 0.5−6.6 | 0.7 ± 0.2 0.5–1.2 | 4 ± 1.2 0.5–6 | 2.5 ± 1.1 0.5–3.2 | 5 ± 1.2 1.2–7.1 | 2.5 ± 0.8 1.2–3.5 |
| Σ ions, mg/L | 63.7 ± 7.5 55–173 | 95 ± 12.0 86–173 | 36.6 ± 4.5 27–56 | 36.5 ± 2.3 30–45 | 81.3 ± 27 13–176 | 82 ± 9 67–112 |
| Lakes area | Bolshaya Imandra | Kondopoga Bay of Onega Lake | Volkhov Bay of Ladoga Lake | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Periods | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 |
| pH | 7.1 ± 0.2 6.4–7.2 | 7.3 ± 0.3 6.3–8.2 | 7.4 ± 0.2 6.9–7.7 | 7.0 ± 0.2 6.3–7.2 | 7.6 ± 0.3 6.5–8.4 | 7.4 ± 0.3 6.5–7.8 | 7.3 ± 0.2 6.5–7.5 | 7.5 ± 0.3 6.6–9.3 | 7.4 ± 0.2 6.7–9.0 |
| Ptot, µg/L | 6 ± 1 2–10 | 70 ± 30 2–176 | 26 ± 7 13–68 | 8 ± 1 3–11 | 54 ± 15 6–98 | 23 ± 9 8–36 | 10 ± 1 1–25 | 178 ± 50 180–260 | 30 ± 5 10–137 |
| PO4, µg/L | 1 ± 0.1 0–8 | 21 ± 5 2–154 | 6 ± 2 0–12 | 1 ± 0.1 0.1–9 | 30 ± 12 5–69 | 4 ± 1 0–15 | 3 ± 0.2 0.3–7 | 100 ± 11 5–9 | 8 ± 1 0.4–5 |
| Ntot, µg/L | 10 ± 3 10–100 | 436 ± 98 164–1925 | 203 ± 35 106–402 | 350 ± 50 150–420 | 750 ± 75 450–900 | 620 ± 55 400–820 | 450 ± 27 120–610 | 920 ± 97 540–1620 | 805 ± 56 730–1230 |
| NO3, µg/L | 1 ± 0.2 0–35 | 102 ± 31 1–1271 | 7 ± 1 1–158 | 110 ± 9 10–220 | 120 ± 9 44–150 | 77 ± 21 35–140 | 130 ± 12 50–150 | 240 ± 17 120–450 | 220 ± 25 96–361 |
| Si, µg/L | 0.6 ± 0.2 0.3–0.7 | 1.1 ± 1.1 0.1–5.2 | 0.6 ± 0.3 0.1–1.5 | 2.0 ± 0.2 0.2–2.5 | 1.2 ± 0.2 0.1–3.8 | 0.3 ± 0.1 0.1–1.5 | 1.0 ± 0.1 0.6–1.2 | 0.5 ± 0.1 0.3–0.9 | 0.2 0.1–0.5 |
| Chl “a”, µg/m3 | 0.3 ± 0.1 0.2–0.5 | 3.8 ± 0.7 2–7 | 3.4 ± 0.5 1.5–6 | 0.7 ± 0.1 0.2–0.9 | 8.4 ± 0.7 0.5–11 | 8.6 ± 0.5 1.5–18 | 0.7 ± 0.1 0.3–9 | 8.0 ± 0.5 2–39 | 9.6 ± 1 0.6–10.3 |
| Toxic loads (ΣCi/MPCi) ** | 0.1 ± 0.1 0.02–0.4 | 27.0 ± 7 12–35 | 2.1 ± 1 1.2–17 | 0.1 ± 0.1 0.01–0.03 | 0.7 ± 0.3 0.4–1.3 | 0.9 ± 0.2 0.4–2.3 | 0.1 ± 0.1 0.02–0.6 | 0.8 ± 0.2 0.2–2.5 | 0.8 ± 0.2 0.2–2.7 |
| Phytoplankton | |||||||||
| Biomass, g/m3 | 0.1 ± 0.1 0.01–0.5 | 3.6 ± 0.5 0.4–20.5 | 3.8 ± 0.3 0.5–9.2 | 0.1 ± 0.1 0.05–0.5 | 2.4 ± 0.9 0.5–5.9 | 2.2 ± 0.5 0.5–3.6 | 0.5 ± 0.1 0.1–0.9 | 5.5 ± 1.2 0.2–7.8 | 2.5 ± 0.7 0.2–6.7 |
| Number, cell 106/L | 0.1 ± 0.1 0.01–0.3 | 3.8 ± 0.3 0.9–7.8 | 4.4 ± 0.3 0.7–8.2 | 0.1 ± 0.1 0.02–0.5 | 3.6 ± 1.4 0.5–8.3 | 3.4 ± 1.2 0.4–8.5 | 0.4 ± 0.2 0.04–0.9 | 12.3 ± 3.8 1–17 | 3.7 ± 2.2 1–9 |
| H (Shannon’s index) | 3.2 ± 0.2 3.0–3.5 | 2.5 ± 0.2 2.0–3.5 | 2.8 ± 0.2 2.0–3.6 | 3.7 ± 0.5 3.1–3.9 | 3.3 ± 0.6 2.9–3.8 | 4.6 ± 0.3 3.0–4.8 | 3.4 ± 0.2 3.1–3.9 | 3.1 ± 0.7 2.5–3.8 | 5.4 ± 0.4 2.8–5.6 |
| Zooplankton | |||||||||
| Biomass, g/m3 | 0.3 ± 0.1 0.2–1.0 | 1.7 ± 1.1 1–3.5 | 0.8 ± 1 0.5–2.5 | 0.3 ± 0.1 0.1–0.6 | 2.9 ± 0.7 0.8–4.1 | 1.4 ± 0.5 0.9–3.2 | 0.6 ± 0.2 0.3–0.7 | 2.8 ± 0.4 0.6–3.1 | 1.9 ± 0.3 0.6–2.5 |
| Number, spec, 103/m3 | 15.0 ± 10 10–100 | 271.0 ± 139 78–320 | 445.0 ± 99 98–510 | 10.0 ± 1.2 5–20 | 110.0 ± 37 15–130 | 91.0 ± 25 20–110 | 13.0 ± 7.2 10–120 | 143.0 ± 45 48–240 | 58.0 ± 27 22–121 |
| H (Shannon’s index) | 2.8 ± 0.2 2.5–3.0 | 1.9 ± 1 1–2.5 | 2.0 ± 0.4 1.5–3 | 2.3 ± 0.4 2–3 | 1.7 ± 0.5 1–3.5 | 3.7 ± 0.4 2–4 | - | - | 4.1 ± 0.5 2.8–4.3 |
| Macrozoobenthos | |||||||||
| Biomass, g/m2 | 0.6 ± 0.1 0.3–0.8 | 49.0 ± 9 5–60 | 23.8 ± 7 7–50 | 0.6 ± 0.2 0.4–1 | 3.2 ± 1.1 1–58 | 15.2 ± 3.6 5–60 | 1.6 ± 0.3 0.2–13 | 4.8 ± 2.4 0.2–19 | 12.9 ± 7.1 1–28 |
| Number, spec, 103/m2 | 0.5 ± 0.1 0.4–0.7 | 6.2 ± 2 1–12 | 4.8 ± 3 1–9 | 0.2 ± 0.1 0.1–0.4 | 2.4 ± 1.5 0.5–41 | 8.5 ± 2.3 1–50 | 0.8 ± 0.2 0.2–5 | 1.1 ± 0.9 0.4–6.5 | 3.7 ± 1.5 0.6–10.1 |
| H (Shannon’s index) | 3.5 ± 0.2 2.1–3.8 | 1.6 ± 0.3 1.0–2.2 | 1.2 ± 0.2 1.0–2.1 | 2.6 ± 0.5 2.2–3.1 | 2.0 ± 0.7 1.1–2.9 | 2.2 ± 0.3 1.7–3.0 | - | - | 2.4 ± 0.5 1.5–3.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moiseenko, T.; Sharov, A. Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review. Geosciences 2019, 9, 492. https://doi.org/10.3390/geosciences9120492
Moiseenko T, Sharov A. Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review. Geosciences. 2019; 9(12):492. https://doi.org/10.3390/geosciences9120492
Chicago/Turabian StyleMoiseenko, Tatiana, and Andrey Sharov. 2019. "Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review" Geosciences 9, no. 12: 492. https://doi.org/10.3390/geosciences9120492
APA StyleMoiseenko, T., & Sharov, A. (2019). Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review. Geosciences, 9(12), 492. https://doi.org/10.3390/geosciences9120492