Plastic Accumulation in the Sea Surface Microlayer: An Experiment-Based Perspective for Future Studies


Abstract
:1. Introduction
1.1. The Structure of the Sea Surface Microlayer, and the Air–Sea Interface
1.2. Plastic Accumulation and Other Pollutants in the Sea Surface Microlayer
2. Materials and Methods
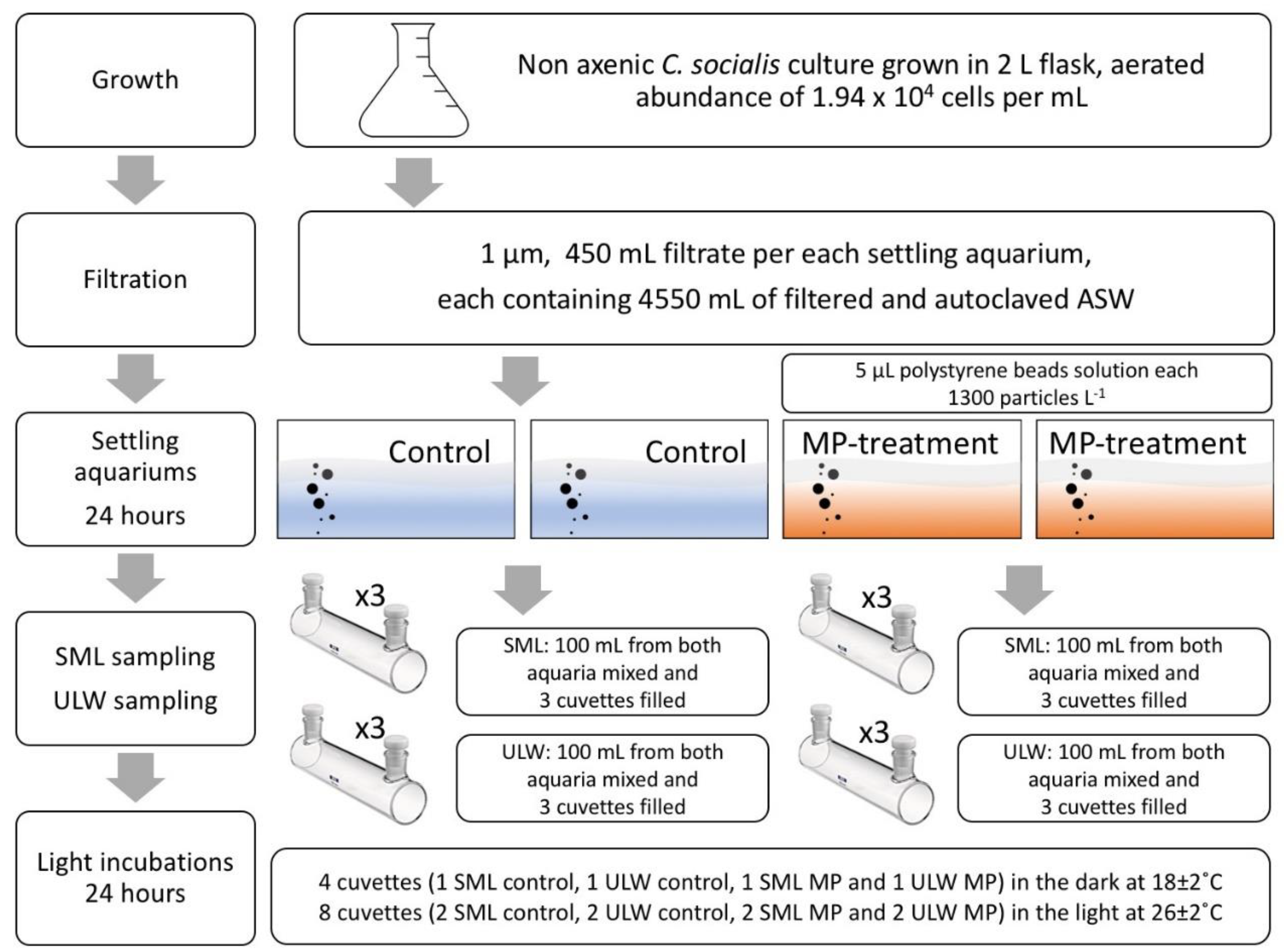
2.1. Experimental Set-Up
2.2. Parameters
2.3. Analysis and Statistics
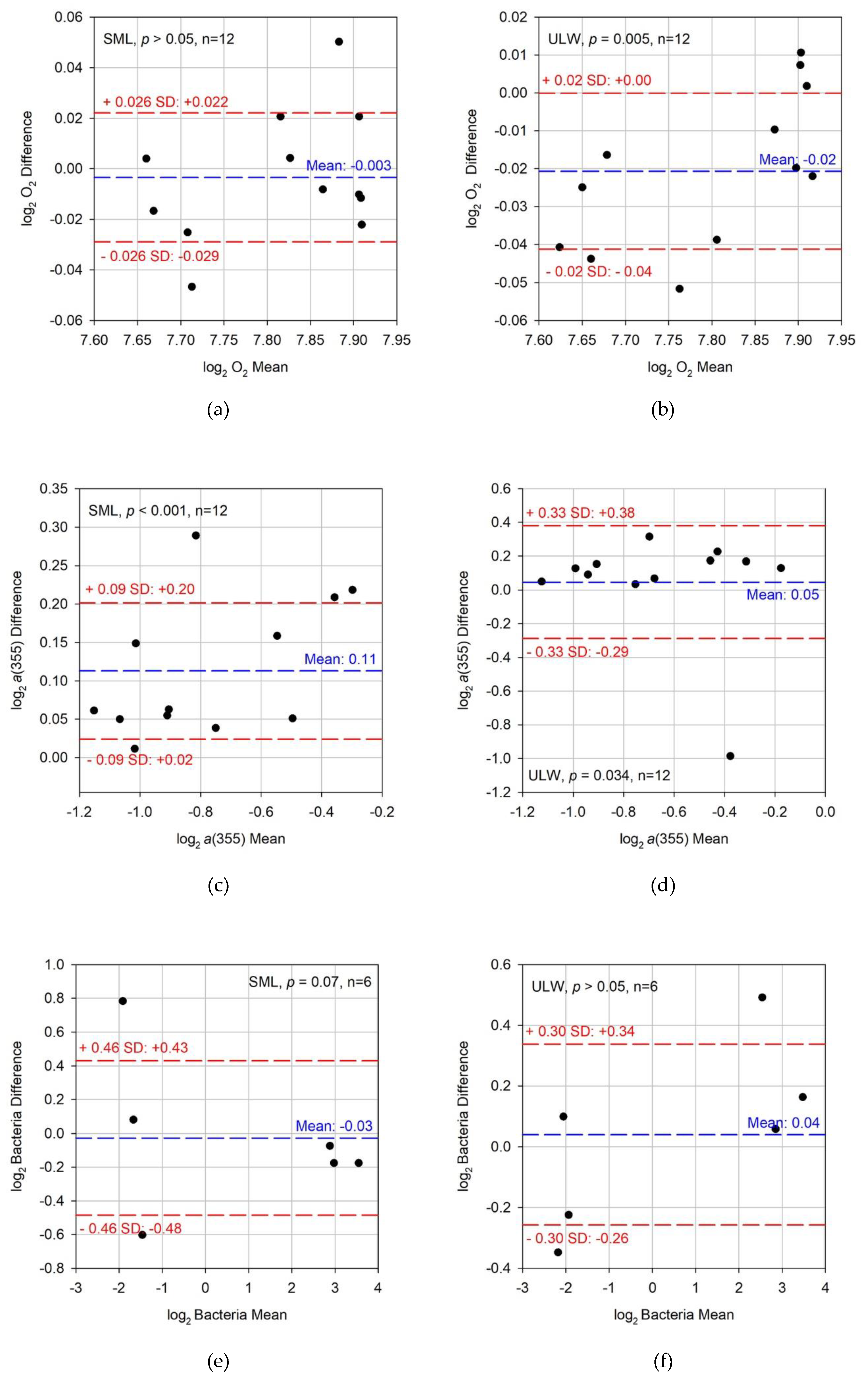
3. Results
3.1. Differences between Dark and Light Conditions
3.2. Differences between Controls and MP Treatments
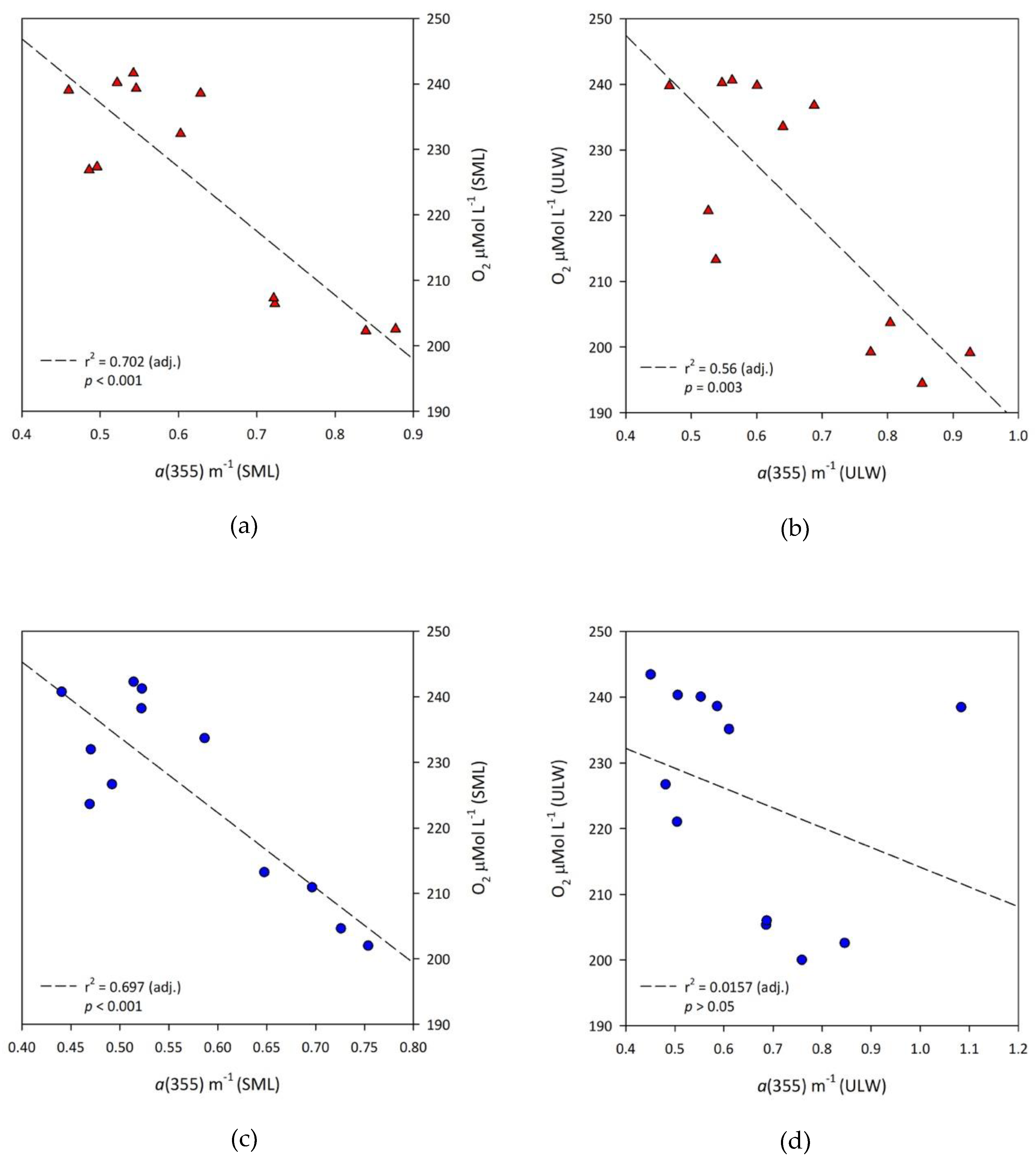
3.3. Correlations between Parameters
- Controls-ULW: O2 = 244.30 µmol·L−1−(30.15 × a(355));
- MP treatments-ULW: O2 = 286.96 µmol·L−1−(98.69 × a(355)).
3.4. Enrichment Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hardy, J.T. The sea surface microlayer: Biology, chemistry and anthropogenic enrichment. Progr. Oceanogr. 1982, 11, 307–328. [Google Scholar] [CrossRef]
- Sieburth, J.M. Microbiological and organic-chemical processes in the surface and mixed layers. In Air-Sea Exchange of Gases and Particles; D. Reidel Publishing Company: Dordrecht, The Netherlands, 1983. [Google Scholar]
- Williams, P.M. Sea surface chemistry: Organic carbon and organic and inorganic nitrogen and phosphorus in surface films and subsurface waters. Deep-Sea Res. Oceanogr. Abstr. 1967, 14, 791–800. [Google Scholar] [CrossRef]
- Wurl, O.; Wurl, E.; Miller, L.; Johnson, K.; Vagle, S. Formation and global distribution of sea-surface microlayers. Biogeosciences 2011, 8, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J.T.; Apts, C.W.; Crecelius, E.A.; Fellingham, G.W. The sea-surface microlayer: Fate and residence times of atmospheric metals. Limnol. Oceanogr. 1985, 30, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Liss, P.S.; Duce, R.A. The Sea Surface and Global Change; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Sieburth, J.M.; Conover, J.T. Slicks associated with trichodesmium blooms in the sargasso sea. Nature 1965, 205, 830. [Google Scholar] [CrossRef]
- Cunliffe, M.; Engel, A.; Frka, S.; Gašparović, B.; Guitart, C.; Murrell, J.C.; Salter, M.; Stolle, C.; Upstill-Goddard, R.; Wurl, O. Sea surface microlayers: A unified physicochemical and biological perspective of the air-ocean interface. Progr. Oceanogr. 2013, 109, 104–116. [Google Scholar] [CrossRef]
- Engel, A.; Bange, H.W.; Cunliffe, M.; Burrows, S.M.; Friedrichs, G.; Galgani, L.; Herrmann, H.; Hertkorn, N.; Johnson, M.; Liss, P.S.; et al. The ocean’s vital skin: Toward an integrated understanding of the sea surface microlayer. Front. Mar. Sci. 2017, 4, 165. [Google Scholar] [CrossRef]
- Wurl, O.; Ekau, W.; Landing, W.M.; Zappa, C.J. Sea surface microlayer in a changing ocean—A perspective. Elem. Sci. Anthr. 2017, 5. [Google Scholar] [CrossRef]
- Henrichs, S.M.; Williams, P.M. Dissolved and particulate amino acids and carbohydrates in the sea surface microlayer. Mar. Chem. 1985, 17, 141–163. [Google Scholar] [CrossRef]
- Verdugo, P. Marine microgels. Annu. Rev. Mar. Sci. 2012, 4, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Wurl, O.; Stolle, C.; Van Thuoc, C.; The Thu, P.; Mari, X. Biofilm-like properties of the sea surface and predicted effects on air–sea CO2 exchange. Progr. Oceanogr. 2016, 144, 15–24. [Google Scholar] [CrossRef]
- Wurl, O.; Holmes, M. The gelatinous nature of the sea-surface microlayer. Mar. Chem. 2008, 110, 89–97. [Google Scholar] [CrossRef]
- Cunliffe, M.; Murrell, J.C. The sea-surface microlayer is a gelatinous biofilm. ISME J. 2009, 3, 1001–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galgani, L.; Engel, A. Changes in optical characteristics of surface microlayers hint to photochemically and microbially mediated Dom turnover in the upwelling region off the coast of Peru. Biogeosciences 2016, 13, 2453–2473. [Google Scholar] [CrossRef]
- Tilstone, G.H.; Airs, R.L.; Vicente, V.M.; Widdicombe, C.; Llewellyn, C. High concentrations of mycosporine-like amino acids and colored dissolved organic matter in the sea surface microlayer off the iberian peninsula. Limnol. Oceanogr. 2010, 55, 1835–1850. [Google Scholar] [CrossRef]
- Mustaffa, N.I.H.; Ribas-Ribas, M.; Wurl, O. High-resolution variability of the enrichment of fluorescence dissolved organic matter in the sea surface microlayer of an upwelling region. Elem. Sci. Anthr. 2017, 5. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, G. Chemical properties of colored dissolved organic matter in the sea-surface microlayer and subsurface water of Jiaozhou bay, China in autumn and winter. Acta Oceanol. Sin. 2013, 32, 26–39. [Google Scholar] [CrossRef]
- Coble, P.G. Marine optical biogeochemistry: The chemistry of ocean color. Chem. Rev. 2007, 107, 402–418. [Google Scholar] [CrossRef] [PubMed]
- Bracchini, L.; Dattilo, A.M.; Falcucci, M.; Hull, V.; Tognazzi, A.; Rossi, C.; Loiselle, S.A. Competition for spectral irradiance between epilimnetic optically active dissolved and suspended matter and phytoplankton in the metalimnion. Consequences for limnology and chemistry. Photochem. Photobiol. Sci. 2011, 10, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Orellana, M.V.; Verdugo, P. Ultraviolet radiation blocks the organic carbon exchange between the dissolved phase and the gel phase in the ocean. Limnol. Oceanogr. 2003, 48, 1618–1623. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Ortega-Retuerta, E.; Passow, U.; Duarte, C.M.; Reche, I. Effects of ultraviolet b radiation on (not so) transparent exopolymer particles. Biogeosciences 2009, 6, 3071–3080. [Google Scholar] [CrossRef]
- Kieber, D.J.; McDaniel, J.; Mopper, K. Photochemical source of biological substrates in sea water: Implications for carbon cycling. Nature 1989, 341, 637–639. [Google Scholar] [CrossRef]
- Loiselle, S.; Vione, D.; Minero, C.; Maurino, V.; Tognazzi, A.; Dattilo, A.M.; Rossi, C.; Bracchini, L. Chemical and optical phototransformation of dissolved organic matter. Water Res. 2012, 46, 3197–3207. [Google Scholar] [CrossRef] [PubMed]
- Del Giorgio, P.A.; Duarte, C.M. Respiration in the open ocean. Nature 2002, 420, 379–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garabétian, F. Production de CO2 à l’interface air-mer. Une approche par l’étude des phénomènes respiratoires dans la microcouche de surface. CO2 production at the sea-air interface. An approach by the study of respiratory processes in surface microlayer. Int. Rev. Gesamten Hydrobiol. 1990, 75, 219–229. [Google Scholar] [CrossRef]
- Andrady, A.L. Microplastics in the marine environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Wurl, O.; Obbard, J.P. A review of pollutants in the sea-surface microlayer (SML): A unique habitat for marine organisms. Mar. Pollut. Bull. 2004, 48, 1016–1030. [Google Scholar] [CrossRef]
- Eriksen, M.; Lebreton, L.C.M.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G.; Reisser, J. Plastic pollution in the world’s oceans: More than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS ONE 2014, 9, e111913. [Google Scholar] [CrossRef]
- Worm, B.; Lotze, H.K.; Jubinville, I.; Wilcox, C.; Jambeck, J. Plastic as a persistent marine pollutant. Annu. Rev. Environ. Resour. 2017, 42, 1–26. [Google Scholar] [CrossRef]
- Villarrubia-Gómez, P.; Cornell, S.E.; Fabres, J. Marine plastic pollution as a planetary boundary threat—The drifting piece in the sustainability puzzle. Mar. Policy 2018, 96, 213–220. [Google Scholar] [CrossRef]
- Lavender Law, K. Plastics in the marine environment. Annu. Rev. Mar. Sci. 2017, 9, 205–229. [Google Scholar]
- Vegter, A.C.; Barletta, M.; Beck, C.; Borrero, J.; Burton, H.; Campbell, M.L.; Costa, M.F.; Eriksen, M.; Eriksson, C.; Estrades, A.; et al. Global research priorities to mitigate plastic pollution impacts on marine wildlife. Endanger. Species Res. 2014, 25, 225–247. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, C.; Puckridge, M.; Schuyler, Q.A.; Townsend, K.; Hardesty, B.D. A quantitative analysis linking sea turtle mortality and plastic debris ingestion. Sci. Rep. 2018, 8, 12536. [Google Scholar] [CrossRef] [PubMed]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.J.; Anderson, S.J.; Harvey, G.R.; Miklas, H.P.; Peck, B.B. Polystyrene spherules in coastal waters. Science 1972, 178, 749–750. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.J.; Smith, K.L. Plastics on the sargasso sea surface. Science 1972, 175, 1240–1241. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Hong, S.H.; Jang, M.; Kang, J.-H.; Kwon, O.Y.; Han, G.M.; Shim, W.J. Large accumulation of micro-sized synthetic polymer particles in the sea surface microlayer. Environ. Sci. Technol. 2014, 48, 9014–9021. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Hong, S.H.; Jang, M.; Han, G.M.; Shim, W.J. Occurrence and distribution of microplastics in the sea surface microlayer in Jinhae bay, South Korea. Arch. Environ. Contam. Toxicol. 2015, 69, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Chae, D.-H.; Kim, I.-S.; Kim, S.-K.; Song, Y.K.; Shim, W.J. Abundance and distribution characteristics of microplastics in surface seawaters of the Incheon/Kyeonggi coastal region. Arch. Environ. Contam. Toxicol. 2015, 69, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Besseling, E.; Wegner, A.; Foekema, E.M. Plastic as a carrier of pops to aquatic organisms: A model analysis. Environ. Sci. Technol. 2013, 47, 7812–7820. [Google Scholar] [CrossRef] [PubMed]
- Teuten, E.L.; Rowland, S.J.; Galloway, T.S.; Thompson, R.C. Potential for plastics to transport hydrophobic contaminants. Environ. Sci. Technol. 2007, 41, 7759–7764. [Google Scholar] [CrossRef] [PubMed]
- Garabetian, F.; Romano, J.-C.; Paul, R.; Sigoillot, J.-C. Organic matter composition and pollutant enrichment of sea surface microlayer inside and outside slicks. Mar. Environ. Res. 1993, 35, 323–339. [Google Scholar] [CrossRef]
- Wells, M.L.; Kozelka, P.B.; Bruland, K.W. The complexation of ‘dissolved’ Cu, Zn, Cd and Pb by soluble and colloidal organic matter in Narragansett bay, Ri. Mar. Chem. 1998, 62, 203–217. [Google Scholar] [CrossRef]
- Benner, R. Chemical composition and reactivity. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, D.J., Eds.; Academic Press—Elsevier: Cambridge, MA, USA, 2002; pp. 59–90. [Google Scholar]
- Benner, R.; Pakulski, J.D.; McCarthy, M.; Hedges, J.I.; Hatcher, P.G. Bulk chemical characteristics of dissolved organic matter in the ocean. Science 1992, 255, 1561–1564. [Google Scholar] [CrossRef] [PubMed]
- Galgani, L.; Engel, A.; Rossi, C.; Donati, A.; Loiselle, S.A. Polystyrene microplastics increase microbial release of marine chromophoric dissolved organic matter in microcosm experiments. Sci. Rep. 2018, 8, 14635. [Google Scholar] [CrossRef] [PubMed]
- Romera-Castillo, C.; Pinto, M.; Langer, T.M.; Álvarez-Salgado, X.A.; Herndl, G.J. Dissolved organic carbon leaching from plastics stimulates microbial activity in the ocean. Nat. Commun. 2018, 9, 1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GESAMP. The Sea-Surface Microlayer and Its Role in Global Change; WMO: Geneva, Switzerland, 1995; Volume 52, p. 92. [Google Scholar]
- Galgani, L.; Piontek, J.; Engel, A. Biopolymers form a gelatinous microlayer at the air-sea interface when arctic sea ice melts. Sci. Rep. 2016, 6, 29465. [Google Scholar] [CrossRef] [PubMed]
- Galgani, L.; Stolle, C.; Endres, S.; Schulz, K.G.; Engel, A. Effects of ocean acidification on the biogenic composition of the sea-surface microlayer: Results from a mesocosm study. J. Geophys. Res.-Oceans 2014, 119, 7911–7924. [Google Scholar] [CrossRef] [Green Version]
- Rahlff, J.; Stolle, C.; Giebel, H.-A.; Brinkhoff, T.; Ribas-Ribas, M.; Hodapp, D.; Wurl, O. High wind speeds prevent formation of a distinct bacterioneuston community in the sea-surface microlayer. FEMS Microbiol. Ecol. 2017, 93, fix041. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, G.M. Formulae and Methods IV [i.e., 4th ed.] of the Marine Biological Laboratory Chemical Room; The Marine Biological Laboratory: Woods Hole, MA, USA, 1975; Volume 2016. [Google Scholar]
- Bricaud, A.; Morel, A.; Prieur, L. Absorption by dissolved organic matter of the sea (yellow substance) in the uv and visible domains. Limnol. Oceanogr. 1981, 26, 43–53. [Google Scholar] [CrossRef]
- Engel, A.; Galgani, L. The organic sea-surface microlayer in the upwelling region off the coast of Peru and potential implications for air–sea exchange processes. Biogeosciences 2016, 13, 989–1007. [Google Scholar] [CrossRef] [Green Version]
- Dreshchinskii, A.; Engel, A. Seasonal variations of the sea surface microlayer at the Boknis Eck Times Series Station (Baltic Sea). J. Plankton Res. 2017, 39, 943–961. [Google Scholar] [CrossRef] [Green Version]
- Mustaffa, N.I.H.; Badewien, T.H.; Ribas-Ribas, M.; Wurl, O. High-resolution observations on enrichment processes in the sea-surface microlayer. Sci. Rep. 2018, 8, 13122. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Microbial attachment to particles in marine and freshwater ecosystems. Microb. Ecol. 1975, 2, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Rummel, C.D.; Jahnke, A.; Gorokhova, E.; Kühnel, D.; Schmitt-Jansen, M. Impacts of biofilm formation on the fate and potential effects of microplastic in the aquatic environment. Environ. Sci. Technol. Let. 2017, 4, 258–267. [Google Scholar] [CrossRef]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “plastisphere”: Microbial communities on plastic marine debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef]
- Reisser, J.; Shaw, J.; Hallegraeff, G.; Proietti, M.; Barnes, D.K.A.; Thums, M.; Wilcox, C.; Hardesty, B.D.; Pattiaratchi, C. Millimeter-sized marine plastics: A new pelagic habitat for microorganisms and invertebrates. PLoS ONE 2014, 9, e100289. [Google Scholar] [CrossRef]
- Dussud, C.; Meistertzheim, A.L.; Conan, P.; Pujo-Pay, M.; George, M.; Fabre, P.; Coudane, J.; Higgs, P.; Elineau, A.; Pedrotti, M.L.; et al. Evidence of niche partitioning among bacteria living on plastics, organic particles and surrounding seawaters. Environ. Pollut. 2018, 236, 807–816. [Google Scholar] [CrossRef]
- Kooi, M.; Reisser, J.; Slat, B.; Ferrari, F.F.; Schmid, M.S.; Cunsolo, S.; Brambini, R.; Noble, K.; Sirks, L.-A.; Linders, T.E.W.; et al. The effect of particle properties on the depth profile of buoyant plastics in the ocean. Sci. Rep. 2016, 6, 33882. [Google Scholar] [CrossRef] [Green Version]
- Kooi, M.; Nes, E.H.V.; Scheffer, M.; Koelmans, A.A. Ups and downs in the ocean: Effects of biofouling on vertical transport of microplastics. Environ. Sci. Technol. 2017, 51, 7963–7971. [Google Scholar] [CrossRef] [PubMed]
- Andrady, A.L. Persistence of plastic litter in the oceans. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 57–72. [Google Scholar]
- Sikandar, S.S.; Imtiaz, A.; Muhammad, I. Degradation study of used polystyrene with UV irradiation. Adv. Mater. Sci. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Royer, S.-J.; Ferrón, S.; Wilson, S.T.; Karl, D.M. Production of methane and ethylene from plastic in the environment. PLoS ONE 2018, 13, e0200574. [Google Scholar] [CrossRef] [PubMed]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 0116. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1a | Controls | ||
| Dark D Light L | O2± SE (µMol O2·L−1) | a(355)± SE(m−1) | Bacteria± SE (105 cells·mL−1) |
| SML (D) | 238.0 ± 1.7 (*) p < 0.001 | 0.51 ± 0.02 (*) p = 0.048 (**) p = 0.03 | 3.3 ± 3.0 |
| SML (L) | 213.5 ± 4.1 | 0.63 ± 0.05 | 8.0 ± 0.4 |
| ULW (D) | 239.3 ± 1.1 (*) p < 0.001 | 0.63 ± 0.09 | 2.8 ± 2.6 |
| ULW (L) | 210.3 ± 4.4 | 0.66 ± 0.06 | 6.0 ± 1.1 |
| 1b | MP Treatments | ||
| Dark D Light L | O2 ± SE (µMol O2·L−1) | a(355) ± SE (m−1) | Bacteria ± SE (105 cells·mL−1) |
| SML (D) | 238.5 ± 1.3 (*) p < 0.001 | 0.55 ± 0.02 (*) p = 0.040 (**) p = 0.02 | 3.0 ± 2.7 |
| SML (L) | 212.1 ± 4.8 (**) p = 0.006 | 0.69 ± 0.07 (**) p = 0.004 | 7.3 ± 0.1 |
| ULW (D) | 238.5 ± 1.1 (*) p < 0.001 | 0.58 ± 0.03 | 3.1 ± 2.9 |
| ULW (L) | 205.1 ± 4.1 | 0.74 ± 0.07 | 7.1 ± 0.2 |
| SML | ULW, Controls | ULW, MP Treatments |
|---|---|---|
| O2 | C = 0.881 p < 0.001 n = 12 | C = 0.993 p < 0.001 n = 12 |
| a(355) | C = 0.832 p < 0.001 n = 12 | C = 0.979 p < 0.001 n = 12 |
| Bacteria | C = 0.943 p = 0.0167 n = 6 | C = 1.0 p = 0.0028 n = 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galgani, L.; Loiselle, S.A. Plastic Accumulation in the Sea Surface Microlayer: An Experiment-Based Perspective for Future Studies. Geosciences 2019, 9, 66. https://doi.org/10.3390/geosciences9020066
Galgani L, Loiselle SA. Plastic Accumulation in the Sea Surface Microlayer: An Experiment-Based Perspective for Future Studies. Geosciences. 2019; 9(2):66. https://doi.org/10.3390/geosciences9020066
Chicago/Turabian StyleGalgani, Luisa, and Steven A. Loiselle. 2019. "Plastic Accumulation in the Sea Surface Microlayer: An Experiment-Based Perspective for Future Studies" Geosciences 9, no. 2: 66. https://doi.org/10.3390/geosciences9020066
APA StyleGalgani, L., & Loiselle, S. A. (2019). Plastic Accumulation in the Sea Surface Microlayer: An Experiment-Based Perspective for Future Studies. Geosciences, 9(2), 66. https://doi.org/10.3390/geosciences9020066