
Genital Wound Repair and Scarring




Abstract
:1. Introduction
2. Differences between Genital and Non-Genital Skin
2.1. Developmental Genital Similarities between Sexes
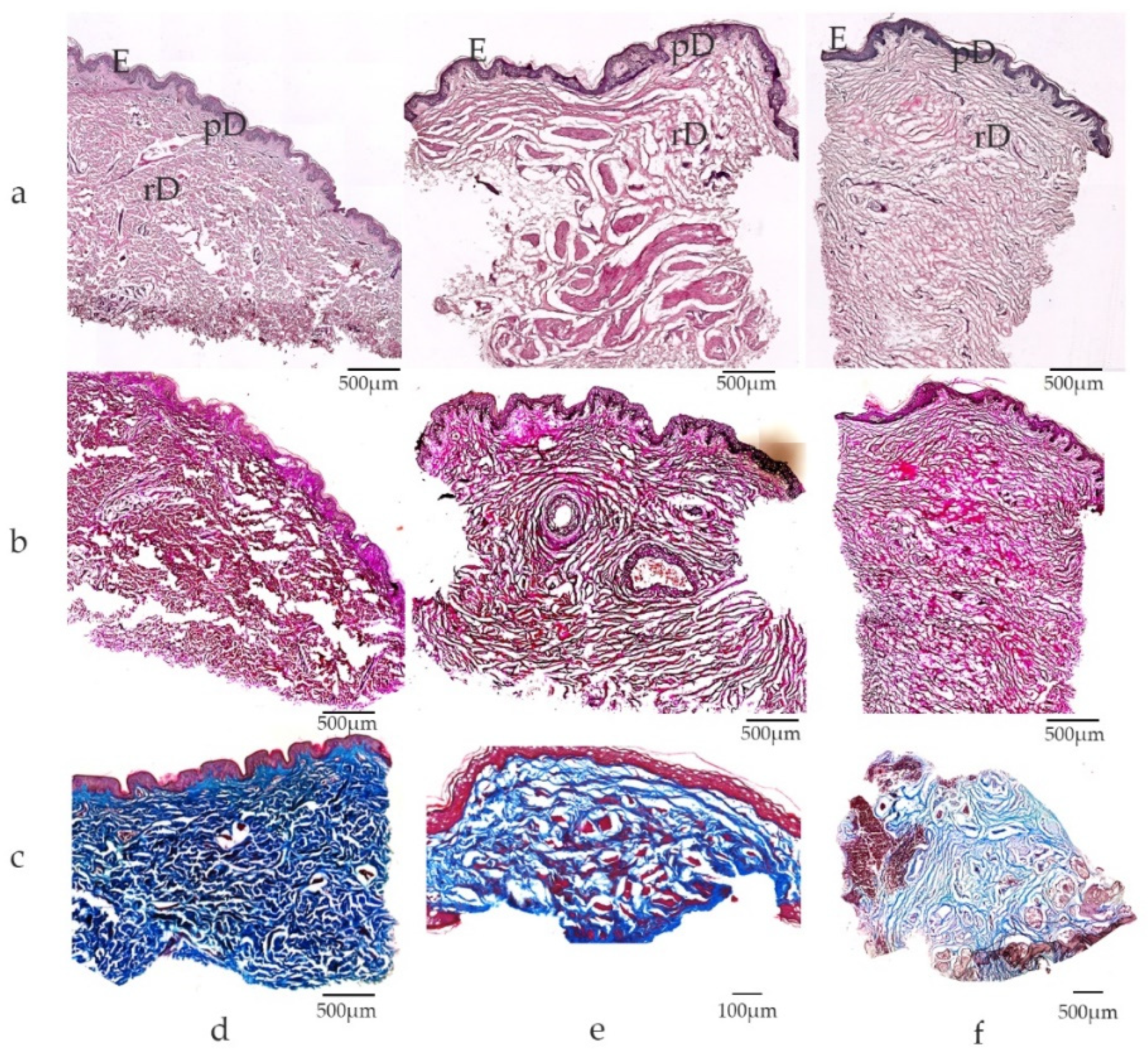
2.2. Cutaneous Microstructure, Biomechanics and Biochemical Composition
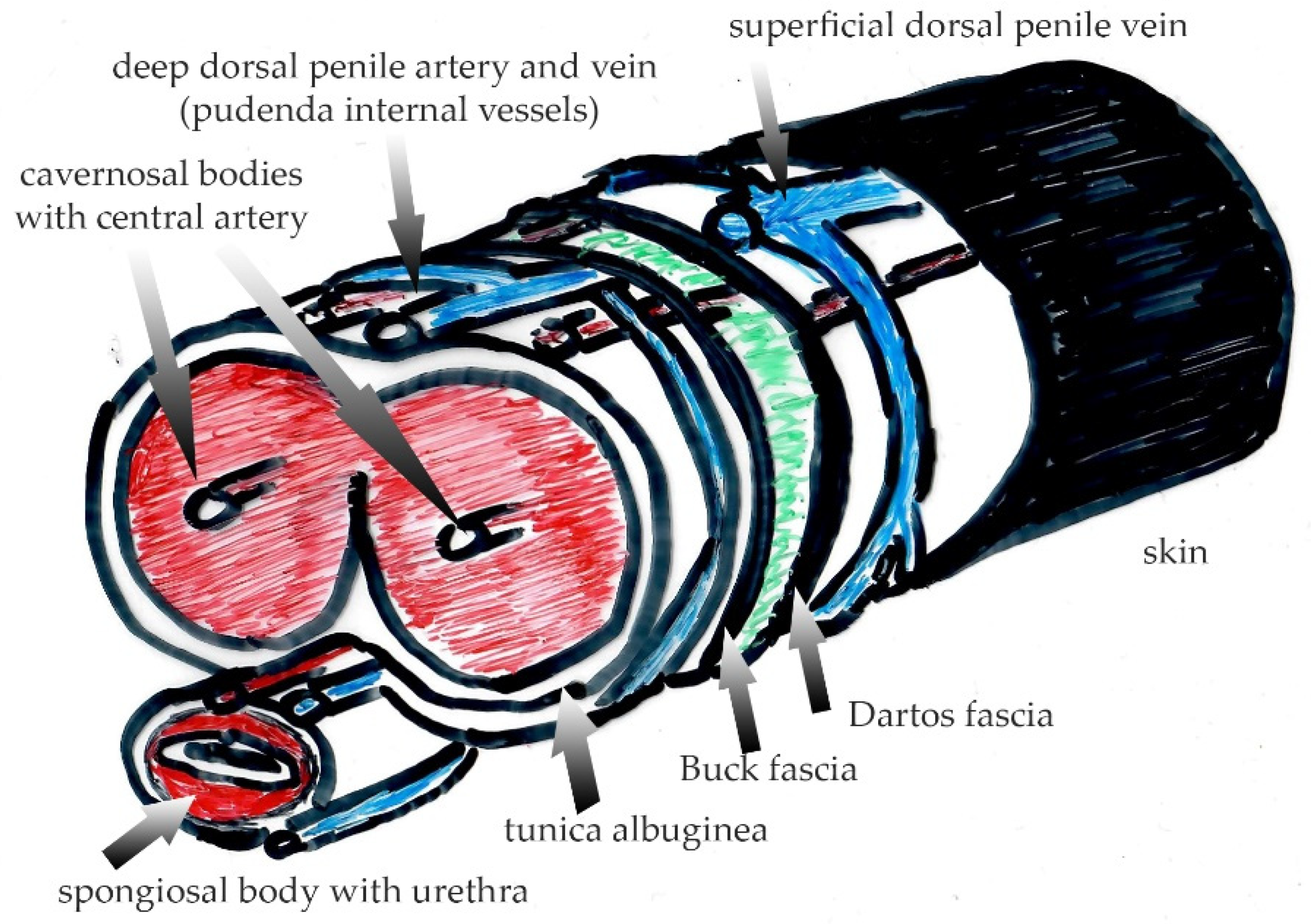
2.3. Macroscopic and Microscopic Anatomy of Genital Skin
2.4. Genital Autocrine Hormone Production and Endocrine Responsivness
2.5. Normal Genital Wound Healing
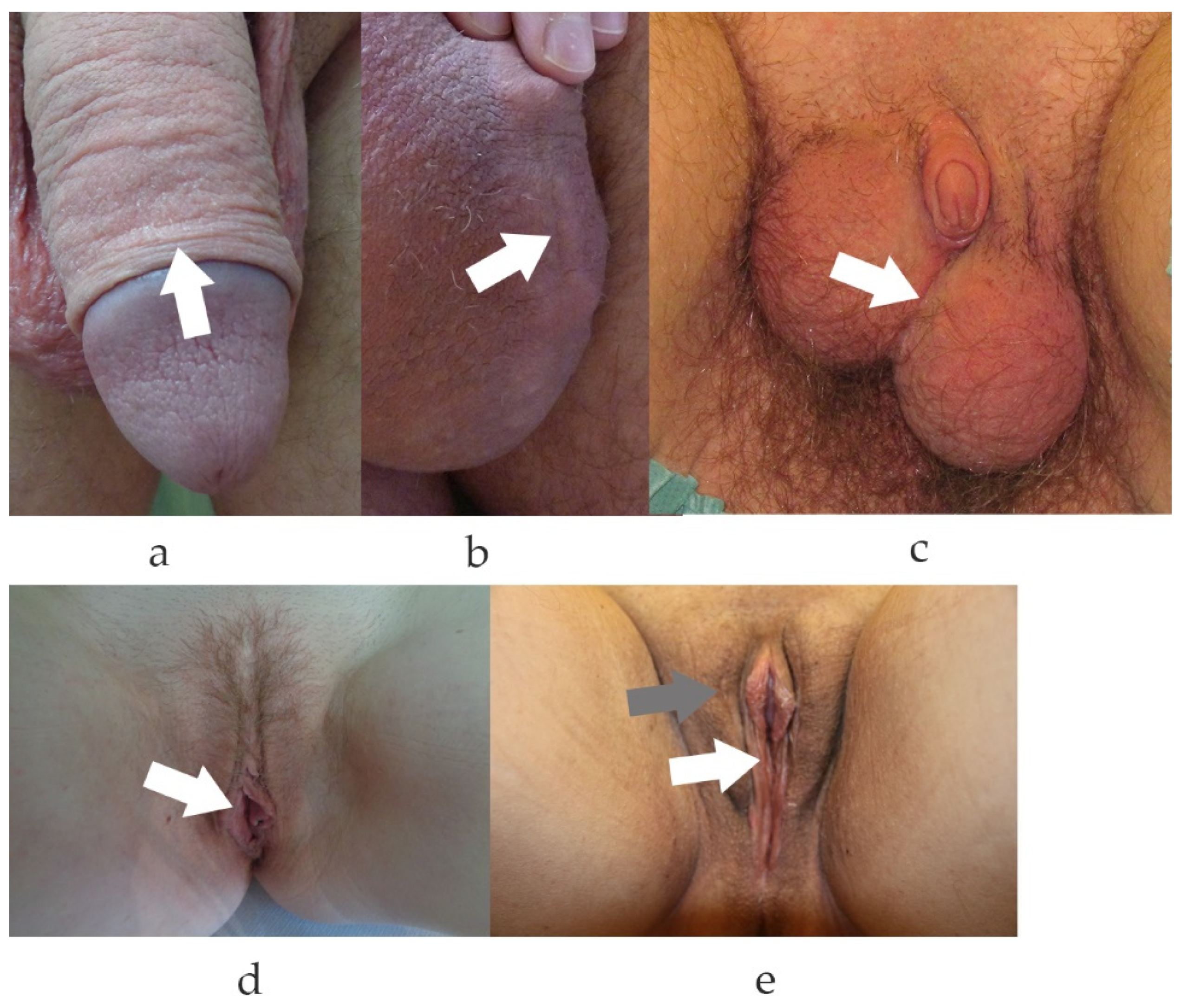
2.6. Pathological Wound Healing of the Genital Skin
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Male Circumcision. Global Trends and Determinants of Prevalence, Safety and Acceptability; WHO Press: Geneva, Switzerland, 2007. [Google Scholar]
- Borkenhagen, A.; Mirastschijski, U.; Strauss, B.; Gieler, U.; Braehler, E. Body hair removal: Prevalence, demographics, and body experience among men and women in Germany. J. Cosmet. Dermatol. 2020, 19, 2886–2892. [Google Scholar] [CrossRef] [PubMed]
- Ismail Aly, M.E.; Huang, T. Management of burn injuries of the perineum. In Total Burn Care, 5th ed.; Herndon, D., Ed.; Elsevier: London, UK, 2018; Volume 1, pp. 609–617. [Google Scholar]
- Harpole, B.G.; Wibbenmeyer, L.A.; Erickson, B.A. Genital burns in the national burn repository: Incidence, etiology, and impact on morbidity and mortality. Urology 2014, 83, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Isbir, D.; Cedidi, C. Acne inversa inguinalis. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1, pp. 243–251. [Google Scholar]
- Lehnhardt, M.; Wallner, C.; Daigeler, A. Reconstruction of the male genitals after Fournier gangrene. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1, pp. 253–263. [Google Scholar]
- Mirastschijski, U.; Remmel, E. Anatomie des weiblichen genitals. In Intimchirurgie; Springer: Berlin/Heidelberg, Germany, 2019; p. 324. [Google Scholar]
- Sharma, R.K.; Parashar, A. The management of perineal wounds. Indian J. Plast. Surg. 2012, 45, 352–363. [Google Scholar] [PubMed]
- Lee, A.; Fischer, G. Diagnosis and Treatment of Vulvar Lichen Sclerosus: An Update for Dermatologists. Am. J. Clin. Dermatol. 2018, 19, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Farage, M.; Maibach, H. Lifetime changes in the vulva and vagina. Arch. Gynecol. Obstet. 2006, 273, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Mirastschijski, U.; Jiang, D.; Rinkevich, Y.; Karim, R.B.; Sorg, H. Wound repair and scarring of genital skin. Plast. Aesthetic Res. 2020, 7, 14. [Google Scholar] [CrossRef]
- Correa-Gallegos, D.; Jiang, D.; Christ, S.; Ramesh, P.; Ye, H.; Wannemacher, J.; Kalgudde Gopal, S.; Yu, Q.; Aichler, M.; Walch, A.; et al. Patch repair of deep wounds by mobilized fascia. Nature 2019, 576, 287–292. [Google Scholar] [CrossRef]
- Jiang, D.; Christ, S.; Correa-Gallegos, D.; Ramesh, P.; Kalgudde Gopal, S.; Wannemacher, J.; Mayr, C.H.; Lupperger, V.; Yu, Q.; Ye, H.; et al. Injury triggers fascia fibroblast collective cell migration to drive scar formation through N-cadherin. Nat. Commun. 2020, 11, 5653. [Google Scholar] [CrossRef]
- Wan, L.; Jiang, D.; Correa-Gallegos, D.; Ramesh, P.; Zhao, J.; Ye, H.; Zhu, S.; Wannemacher, J.; Volz, T.; Rinkevich, Y. Connexin43 gap junction drives fascia mobilization and repair of deep skin wounds. Matrix Biol. 2021, 97, 58–71. [Google Scholar] [CrossRef]
- Mirastschijski, U. Genital scars. In Textbook on Scar Managment. State of the Art Management and Emerging Technologies; Teot, L., Mustoe, T.A., Middelkoop, E., Gauglitz, G., Eds.; Springer: Cham, Switzerland, 2020; pp. 405–425. [Google Scholar]
- Uitto, J.; Olsen, D.R.; Fazio, M.J. Extracellular matrix of the skin: 50 years of progress. J. Investig. Dermatol. 1989, 92 (Suppl. 4), 61S–77S. [Google Scholar] [CrossRef] [Green Version]
- Pfisterer, K.; Shaw, L.E.; Symmank, D.; Weninger, W. The Extracellular Matrix in Skin Inflammation and Infection. Front. Cell Dev. Biol. 2021, 9, 682414. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Brown, R.A.; Mudera, V. Collagen stiffness regulates cellular contraction and matrix remodeling gene expression. J. Biomed. Mater. Res. A 2007, 83, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Mirastschijski, U.; Haaksma, C.J.; Tomasek, J.J.; Agren, M.S. Matrix metalloproteinase inhibitor GM 6001 attenuates keratinocyte migration, contraction and myofibroblast formation in skin wounds. Exp. Cell Res. 2004, 299, 465–475. [Google Scholar] [CrossRef]
- Mirastschijski, U.; Impola, U.; Karsdal, M.A.; Saarialho-Kere, U.; Agren, M.S. Matrix metalloproteinase inhibitor BB-3103 unlike the serine proteinase inhibitor aprotinin abrogates epidermal healing of human skin wounds ex vivo. J. Investig. Dermatol. 2002, 118, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirastschijski, U.; Schnabel, R.; Claes, J.; Schneider, W.; Agren, M.S.; Haaksma, C.; Tomasek, J.J. Matrix metalloproteinase inhibition delays wound healing and blocks the latent transforming growth factor-beta1-promoted myofibroblast formation and function. Wound Repair Regen. 2010, 18, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Heinz, A.; Huertas, A.C.; Schrader, C.U.; Pankau, R.; Gosch, A.; Schmelzer, C.E. Elastins from patients with Williams-Beuren syndrome and healthy individuals differ on the molecular level. Am. J. Med. Genet. A 2016, 170, 1832–1842. [Google Scholar] [CrossRef]
- Andrade, F.; Cardoso, G.P.; Bastos, A.L.; Costa, W.; Chagas, M.; Babinski, M. Structural and stereological analysis of elastic fibers in the glans penis of young men. Rom. J. Morphol. Embryol. 2012, 53, 393–396. [Google Scholar]
- Giannandrea, M.; Parks, W.C. Diverse functions of matrix metalloproteinases during fibrosis. Dis. Models Mech. 2014, 7, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44–46, 113–121. [Google Scholar] [CrossRef]
- Mora Huertas, A.C.; Schmelzer, C.E.; Hoehenwarter, W.; Heyroth, F.; Heinz, A. Molecular-level insights into aging processes of skin elastin. Biochimie 2016, 128–129, 163–173. [Google Scholar] [CrossRef]
- Romppanen, U.; Rantala, I.; Lauslahti, K.; Reunala, T. Light- and electron-microscopic findings in lichen sclerosus of the vulva during etretinate treatment. Dermatologica 1987, 175, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Atmoko, W.; Shalmont, G.; Situmorang, G.R.; Wahyudi, I.; Tanurahardja, B.; Rodjani, A. Abnormal dartos fascia in buried penis and hypospadias: Evidence from histopathology. J. Pediatr. Urol. 2018, 14, 536.e1–536.e7. [Google Scholar] [CrossRef] [PubMed]
- Schill, S.; Panfilov, D.; Mirastschijski, U. Intimchirurgie beim Mann. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 49–68. [Google Scholar]
- Mirastschijski, U.; Schwenke, C.; Schmiedl, A. Plastisch-chirurgische Rekonstruktion des männlichen Genitales. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 189–206. [Google Scholar]
- Gilliver, S.C.; Ashworth, J.J.; Ashcroft, G.S. The hormonal regulation of cutaneous wound healing. Clin. Dermatol. 2007, 25, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Gilliver, S.C.; Ashcroft, G.S. Sex steroids and cutaneous wound healing: The contrasting influences of estrogens and androgens. Climacteric J. Int. Menopause Soc. 2007, 10, 276–288. [Google Scholar] [CrossRef]
- Pomari, E.; Dalla Valle, L.; Pertile, P.; Colombo, L.; Thornton, M.J. Intracrine sex steroid synthesis and signaling in human epidermal keratinocytes and dermal fibroblasts. FASEB J. 2015, 29, 508–524. [Google Scholar] [CrossRef]
- Jiang, H.S.; Zhu, L.L.; Zhang, Z.; Chen, H.; Chen, Y.; Dai, Y.T. Estradiol attenuates the TGF-beta1-induced conversion of primary TAFs into myofibroblasts and inhibits collagen production and myofibroblast contraction by modulating the Smad and Rho/ROCK signaling pathways. Int. J. Mol. Med. 2015, 36, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Kanda, N.; Watanabe, S. 17beta-estradiol enhances the production of granulocyte-macrophage colony-stimulating factor in human keratinocytes. J. Investig. Dermatol. 2004, 123, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Verdier-Sevrain, S.; Yaar, M.; Cantatore, J.; Traish, A.; Gilchrest, B.A. Estradiol induces proliferation of keratinocytes via a receptor mediated mechanism. FASEB J. 2004, 18, 1252–1254. [Google Scholar] [CrossRef]
- Thornton, M.J.; Taylor, A.H.; Mulligan, K.; Al-Azzawi, F.; Lyon, C.C.; O’Driscoll, J.; Messenger, A.G. Oestrogen receptor beta is the predominant oestrogen receptor in human scalp skin. Exp. Dermatol. 2003, 12, 181–190. [Google Scholar] [CrossRef]
- Stevenson, S.; Thornton, J. Effect of estrogens on skin aging and the potential role of SERMs. Clin. Interv. Aging 2007, 2, 283–297. [Google Scholar]
- Emmerson, E.; Campbell, L.; Ashcroft, G.S.; Hardman, M.J. Unique and synergistic roles for 17beta-estradiol and macrophage migration inhibitory factor during cutaneous wound closure are cell type specific. Endocrinology 2009, 150, 2749–2757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, N.; Watanabe, S. 17Beta-estradiol inhibits MCP-1 production in human keratinocytes. J. Investig. Dermatol. 2003, 120, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Watanabe, S. 17beta-estradiol inhibits the production of RANTES in human keratinocytes. J. Investig. Dermatol. 2003, 120, 420–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, N.; Watanabe, S. 17beta-estradiol inhibits the production of interferon-induced protein of 10 kDa by human keratinocytes. J. Investig. Dermatol. 2003, 120, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crompton, R.; Williams, H.; Ansell, D.; Campbell, L.; Holden, K.; Cruickshank, S.; Hardman, M.J. Oestrogen promotes healing in a bacterial LPS model of delayed cutaneous wound repair. Lab. Investig. 2016, 96, 439–449. [Google Scholar] [CrossRef]
- Wilkinson, H.N.; Hardman, M.J. The role of estrogen in cutaneous ageing and repair. Maturitas 2017, 103, 60–64. [Google Scholar] [CrossRef]
- Chabab, A.; Sultan, C. Increased peripheral aromatase activity in prepubertal children with partial androgen insensitivity syndrome. Horm. Res. 1985, 22, 83–88. [Google Scholar] [CrossRef]
- Svenstrup, B.; Brunner, N.; Dombernowsky, P.; Nohr, I.; Micic, S.; Bennett, P.; Spang-Thomsen, M. Comparison of the effect of cortisol on aromatase activity and androgen metabolism in two human fibroblast cell lines derived from the same individual. J. Steroid Biochem. 1990, 35, 679–687. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Chen, W.C.; Thornton, M.J.; Qin, K.; Rosenfield, R. Sexual hormones in human skin. Horm. Metab. Res. 2007, 39, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.R.; Bulun, S.E. Estrogen production and action. J. Am. Acad. Dermatol. 2001, 45 (Suppl. 3), S116–S124. [Google Scholar] [CrossRef]
- Chen, W.; Zouboulis, C.C.; Fritsch, M.; Blume-Peytavi, U.; Kodelja, V.; Goerdt, S.; Luu-The, V.; Orfanos, C.E. Evidence of heterogeneity and quantitative differences of the type 1 5alpha-reductase expression in cultured human skin cells—Evidence of its presence in melanocytes. J. Investig. Dermatol. 1998, 110, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinsky, L.; Finkelberg, R.; Straisfeld, C.; Zilahi, B.; Kaufman, M.; Hall, G. Testosterone metabolism by serially subcultured fibroblasts from genital and nongenital skin of individual human donors. Biochem. Biophys. Res. Commun. 1972, 46, 364–369. [Google Scholar] [CrossRef]
- Hoppe, U.; Holterhus, P.M.; Wunsch, L.; Jocham, D.; Drechsler, T.; Thiele, S.; Marschke, C.; Hiort, O. Tissue-specific transcription profiles of sex steroid biosynthesis enzymes and the androgen receptor. J. Mol. Med. 2006, 84, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zouboulis, C.C.; Orfanos, C.E. The 5 alpha-reductase system and its inhibitors. Recent development and its perspective in treating androgen-dependent skin disorders. Dermatology 1996, 193, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Gad, Y.Z.; Berkovitz, G.D.; Migeon, C.J.; Brown, T.R. Studies of up-regulation of androgen receptors in genital skin fibroblasts. Mol. Cell Endocrinol. 1988, 57, 205–213. [Google Scholar] [CrossRef]
- Sandoval, A.G.W.; Maden, M. Regeneration in the spiny mouse, Acomys, a new mammalian model. Curr. Opin. Genet. Dev. 2020, 64, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Mirastschijski, U. Classification and Treatment of the Adult Buried Penis. Ann. Plast. Surg. 2018, 80, 653–659. [Google Scholar] [CrossRef]
- Mirastschijski, U. Buried Penis. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 107–114. [Google Scholar]
- Mirastschijski, U.; Schwenke, C.; Schwab, I.; Buchhorn, A.; Schmiedl, A. Midline raphe scroti artery flap for penile shaft reconstruction. Plast. Aesthetic Res. 2020, 7, 1–13. [Google Scholar] [CrossRef]
- Schaff, J.; Morath, S.; Mirastschijski, U. Operative Techniken bei Mann-zu-Frau-Transsexualität. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 294–303. [Google Scholar]
- Terlou, A.; Santegoets, L.A.; van der Meijden, W.I.; Heijmans-Antonissen, C.; Swagemakers, S.M.; van der Spek, P.J.; Ewing, P.C.; van Beurden, M.; Helmerhorst, T.J.; Blok, L.J. An autoimmune phenotype in vulvar lichen sclerosus and lichen planus: A Th1 response and high levels of microRNA-155. J. Investig. Dermatol. 2012, 132, 658–666. [Google Scholar] [CrossRef] [Green Version]
- Corazza, M.; Schettini, N.; Zedde, P.; Borghi, A. Vulvar Lichen Sclerosus from Pathophysiology to Therapeutic Approaches: Evidence and Prospects. Biomedicines 2021, 9, 950. [Google Scholar] [CrossRef]
- Fancher, K.; Gardner, J.M.; Shalin, S.C. Elastophagocytosis and interstitial granulomatous infiltrate are more common in extragenital vs. genital lichen sclerosus. J. Cutan. Pathol. 2020, 47, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Kirtschig, G. Lichen Sclerosus-Presentation, Diagnosis and Management. Dtsch. Arztebl. Int. 2016, 113, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouston, D.; Hall, A.; Lawrentschuk, N. Penile lichen sclerosus (balanitis xerotica obliterans). BJU Int. 2011, 108 (Suppl. 2), 14–19. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Xian, D.; Liu, J.; Pan, S.; Tang, R.; Zhong, J. Regulating the Polarization of Macrophages: A Promising Approach to Vascular Dermatosis. J. Immunol. Res. 2020, 2020, 8148272. [Google Scholar] [CrossRef]
- von Fritschen, U.; Strunz, C.; Scherer, R. Female genital mutilation/cutting. In Intimchirurgie; Mirastschijski, U., Remmel, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 117–125. [Google Scholar]
- ISAPS. ISAPS International Survey on Aesthetic/Cosmetic Procedures Performed in 2020. Available online: https://www.isaps.org/wp-content/uploads/2022/01/ISAPS-Global-Survey_2020.pdf (accessed on 11 April 2022).
- Mirastschijski, U.; Schwab, I.; Coger, V.; Zier, U.; Rianna, C.; He, W.; Maedler, K.; Kelm, S.; Radtke, A.; Belge, G.; et al. Lung Surfactant Accelerates Skin Wound Healing: A Translational Study with a Randomized Clinical Phase I Study. Sci. Rep. 2020, 10, 2581. [Google Scholar] [CrossRef]
- Volk, S.W.; Bohling, M.W. Comparative wound healing—Are the small animal veterinarian’s clinical patients an improved translational model for human wound healing research? Wound Repair Regen. 2013, 21, 372–381. [Google Scholar] [CrossRef]
- Hinz, B.; Mastrangelo, D.; Iselin, C.E.; Chaponnier, C.; Gabbiani, G. Mechanical tension controls granulation tissue contractile activity and myofibroblast differentiation. Am. J. Pathol. 2001, 159, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef]
- Glim, J.E.; Beelen, R.H.; Niessen, F.B.; Everts, V.; Ulrich, M.M. The number of immune cells is lower in healthy oral mucosa compared to skin and does not increase after scarring. Arch. Oral Biol. 2015, 60, 272–281. [Google Scholar] [CrossRef]
- Szpaderska, A.M.; Zuckerman, J.D.; DiPietro, L.A. Differential injury responses in oral mucosal and cutaneous wounds. J. Dent. Res. 2003, 82, 621–626. [Google Scholar] [CrossRef]
- Peake, M.A.; Caley, M.; Giles, P.J.; Wall, I.; Enoch, S.; Davies, L.C.; Kipling, D.; Thomas, D.W.; Stephens, P. Identification of a transcriptional signature for the wound healing continuum. Wound Repair Regen. 2014, 22, 399–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, P.; Genever, P. Non-epithelial oral mucosal progenitor cell populations. Oral Dis. 2007, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Calenic, B.; Greabu, M.; Caruntu, C.; Tanase, C.; Battino, M. Oral keratinocyte stem/progenitor cells: Specific markers, molecular signaling pathways and potential uses. Periodontol 2000 2015, 69, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Board-Davies, E.; Moses, R.; Sloan, A.; Stephens, P.; Davies, L.C. Oral Mucosal Lamina Propria-Progenitor Cells Exert Antibacterial Properties via the Secretion of Osteoprotegerin and Haptoglobin. Stem Cells Transl. Med. 2015, 4, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.J.; Leibovich, S.J. Regulation of Macrophage Polarization and Wound Healing. Adv. Wound Care 2012, 1, 10–16. [Google Scholar] [CrossRef] [Green Version]
- DiPietro, L.A.; Wilgus, T.A.; Koh, T.J. Macrophages in Healing Wounds: Paradoxes and Paradigms. Int. J. Mol. Sci. 2021, 22, 950. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Su, W.R.; Shi, S.H.; Wilder-Smith, P.; Xiang, A.P.; Wong, A.; Nguyen, A.L.; Kwon, C.W.; Le, A.D. Human gingiva-derived mesenchymal stem cells elicit polarization of m2 macrophages and enhance cutaneous wound healing. Stem. Cells 2010, 28, 1856–1868. [Google Scholar] [CrossRef] [Green Version]
- Mirastschijski, U.; Caliani, A.J.; Wedekind, D.; McCawley, L.; Agren, M.S. Pro-Inflammatory Matrix Metalloproteinase-3 Is Key Effector of Tnf-Alpha-Induced Type I Collagen Degradation. Wound Repair Regen. 2015, 23, A20. [Google Scholar]
- Mirastschijski, U.; Lupse, B.; Maedler, K.; Sarma, B.; Radtke, A.; Belge, G.; Dorsch, M.; Wedekind, D.; McCawley, L.J.; Boehm, G.; et al. Matrix Metalloproteinase-3 is Key Effector of TNF-alpha-Induced Collagen Degradation in Skin. Int. J. Mol. Sci. 2019, 20, 5234. [Google Scholar] [CrossRef] [Green Version]
- Rehders, M.; Grosshauser, B.B.; Smarandache, A.; Sadhukhan, A.; Mirastschijski, U.; Kempf, J.; Dunne, M.; Slenzka, K.; Brix, K. Effects of lunar and mars dust simulants on HaCaT keratinocytes and CHO-K1 fibroblasts. Adv. Space Res. 2011, 47, 1200–1213. [Google Scholar] [CrossRef]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in wound repair: Molecular and cellular mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallant-Behm, C.L.; Du, P.; Lin, S.M.; Marucha, P.T.; DiPietro, L.A.; Mustoe, T.A. Epithelial regulation of mesenchymal tissue behavior. J. Investig. Dermatol. 2011, 131, 892–899. [Google Scholar] [CrossRef] [Green Version]
- Glim, J.E.; van Egmond, M.; Niessen, F.B.; Everts, V.; Beelen, R.H. Detrimental dermal wound healing: What can we learn from the oral mucosa? Wound Repair Regen 2013, 21, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Crescioli, C.; Maggi, M.; Vannelli, G.B.; Ferruzzi, P.; Granchi, S.; Mancina, R.; Muratori, M.; Forti, G.; Serio, M.; Luconi, M. Expression of functional estrogen receptors in human fetal male external genitalia. J. Clin. Endocrinol. Metab. 2003, 88, 1815–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowa, C.N.; Jesmin, S.; Miyauchi, T. The penis: A new target and source of estrogen in male reproduction. Histol. Histopathol. 2006, 21, 53–67. [Google Scholar]
- Hodgins, M.B.; Spike, R.C.; Mackie, R.M.; MacLean, A.B. An immunohistochemical study of androgen, oestrogen and progesterone receptors in the vulva and vagina. Br. J. Obstet. Gynaecol. 1998, 105, 216–222. [Google Scholar] [CrossRef]
- Kalloo, N.B.; Gearhart, J.P.; Barrack, E.R. Sexually dimorphic expression of estrogen receptors, but not of androgen receptors in human fetal external genitalia. J. Clin. Endocrinol. Metab. 1993, 77, 692–698. [Google Scholar]
- Morimoto, I.; Eto, S.; Inoue, S.; Izumi, M.; Nagataki, S.; Saito, Y.; Hara, T. DHT formation and degradation in cultured human skin fibroblasts: DHT accumulation in the genital skin. J. Steroid Biochem. Mol. Biol. 1991, 38, 227–232. [Google Scholar] [CrossRef]
- Silva, P.D.; Gentzschein, E.E.; Lobo, R.A. Androstenedione may be a more important precursor of tissue dihydrotestosterone than testosterone in women. Fertil. Steril. 1987, 48, 419–422. [Google Scholar] [CrossRef]
- Berkovitz, G.D.; Fujimoto, M.; Brown, T.R.; Brodie, A.M.; Migeon, C.J. Aromatase activity in cultured human genital skin fibroblasts. J. Clin. Endocrinol. Metab. 1984, 59, 665–671. [Google Scholar] [CrossRef]
- Escobar, C.; Rosenblum, N. Vaginal Estrogen-What a Urologist Should Know. Urology 2021, 151, 37–43. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male | Female | Histological Features of the Skin | |
|---|---|---|---|
| Penis/small labia | Penile shaft skin | Labia minores | Penis: keratinizing epithelium, presence of Dartos fascia (Fascia penis superficialis) Labia: outer aspect with thin keratinizing epithelium; inner aspect: mucous membrane Similar for penis and labia: absence of hair and fat; abundance of elastic fibers |
| Glans | penis | clitoridis | Multilayered, non-keratinizing epithelium |
| Foreskin | penis | clitoridis | Outside: cornified epithelium Inside: mucous membrane; no fat |
| Frenulum | Frenulum penis | Frenula clitoridis (paired) | Mucous epithelium, no subcutaneous fat |
| Scrotum/big labia | Scrotum | Labia majores | Hair bearing epidermis (labia: only outer surface), epidermal cornified layer Labia: subcutaneous fat layer and smooth muscle cells; reduction of fat amount with aging Scrotum: fat localized to scrotal wall and between scrotal septa in obese men, contractile Tunica Dartos with smooth muscle cells and myofibroblasts |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirastschijski, U.; Jiang, D.; Rinkevich, Y. Genital Wound Repair and Scarring. Med. Sci. 2022, 10, 23. https://doi.org/10.3390/medsci10020023
Mirastschijski U, Jiang D, Rinkevich Y. Genital Wound Repair and Scarring. Medical Sciences. 2022; 10(2):23. https://doi.org/10.3390/medsci10020023
Chicago/Turabian StyleMirastschijski, Ursula, Dongsheng Jiang, and Yuval Rinkevich. 2022. "Genital Wound Repair and Scarring" Medical Sciences 10, no. 2: 23. https://doi.org/10.3390/medsci10020023
APA StyleMirastschijski, U., Jiang, D., & Rinkevich, Y. (2022). Genital Wound Repair and Scarring. Medical Sciences, 10(2), 23. https://doi.org/10.3390/medsci10020023