
Effect of Yuzu (Citrus junos) Seed Limonoids and Spermine on Intestinal Microbiota and Hypothalamic Tissue in the Sandhoff Disease Mouse Model

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. SD Mouse Model and Dietary Supplementation with Limonoids and Spm
2.3. Histopathological Analysis of Brain Tissues
2.4. Extraction of DNA from Microbiota in Feces
2.5. Intestinal Microbiome Analysis: 16S rRNA Amplicon Sequencing Library Preparation
2.6. Microbiota Analysis of Sequencing Data
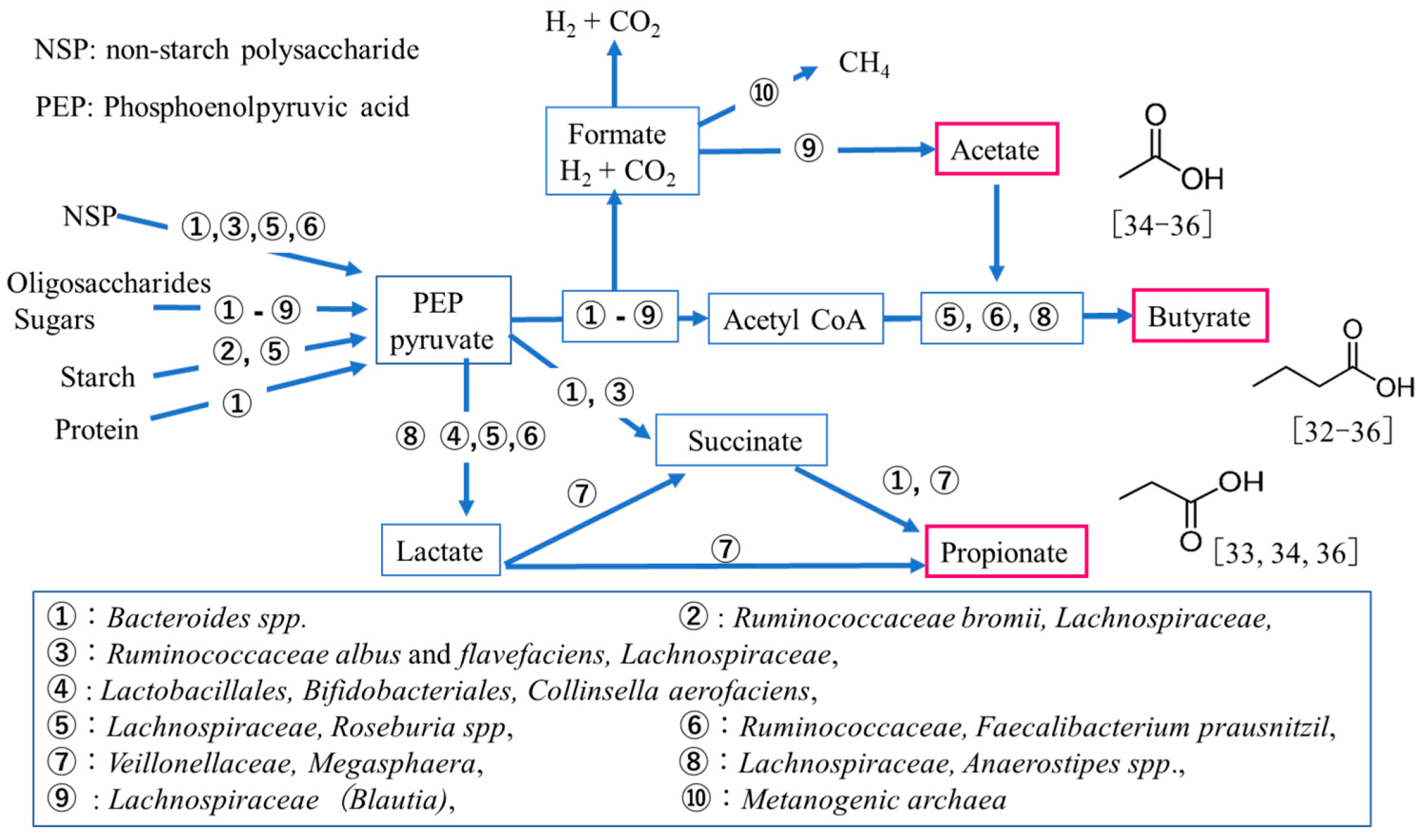
2.7. Quantitative Analysis of Short-Chain Fatty Acids (SCFAs) in Feces
2.8. Quantitative Analysis of IgA in Feces
2.9. Statistical Analysis
3. Results
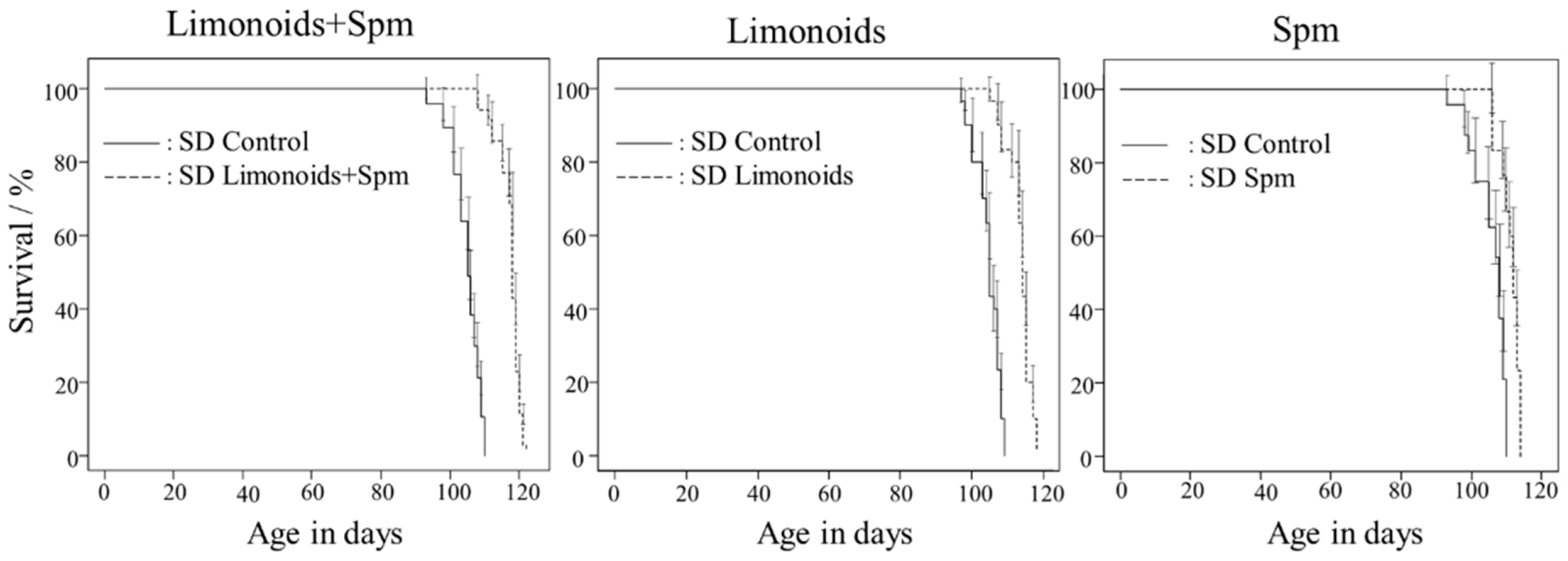
3.1. Survival Rates of SD Mice Consuming Limonoids and/or Spm
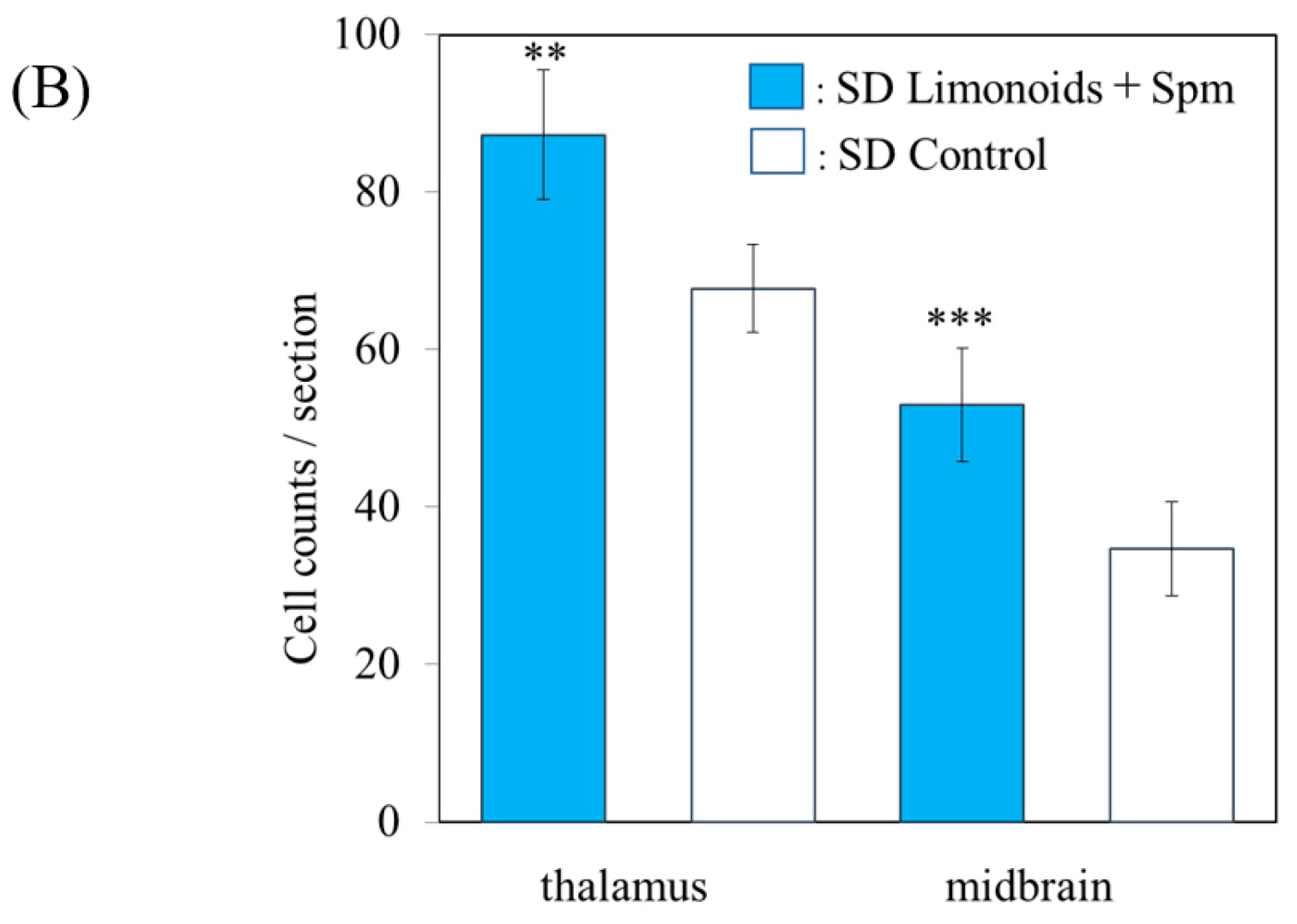
3.2. Histopathological Analysis of Brain Tissue
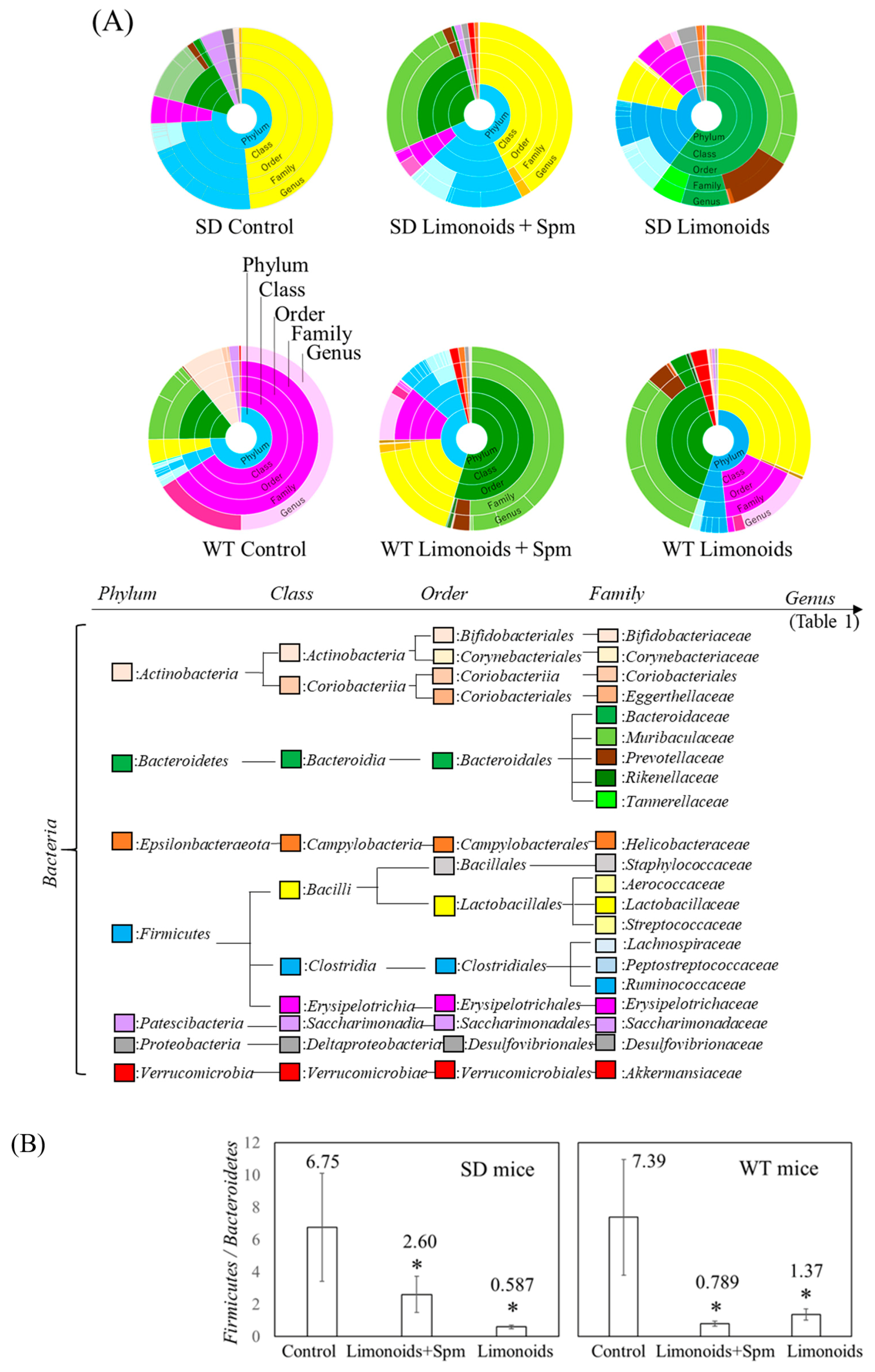
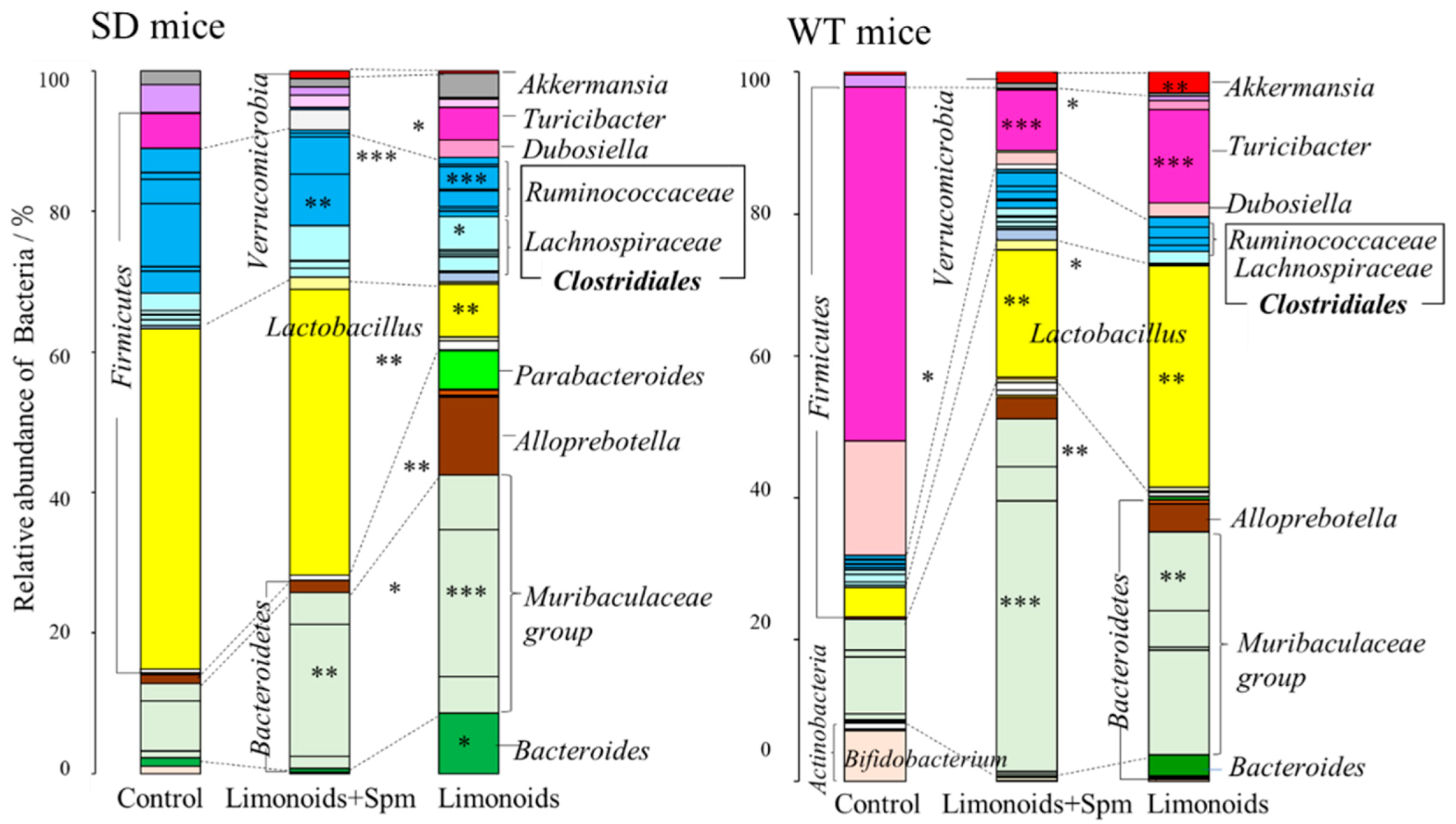
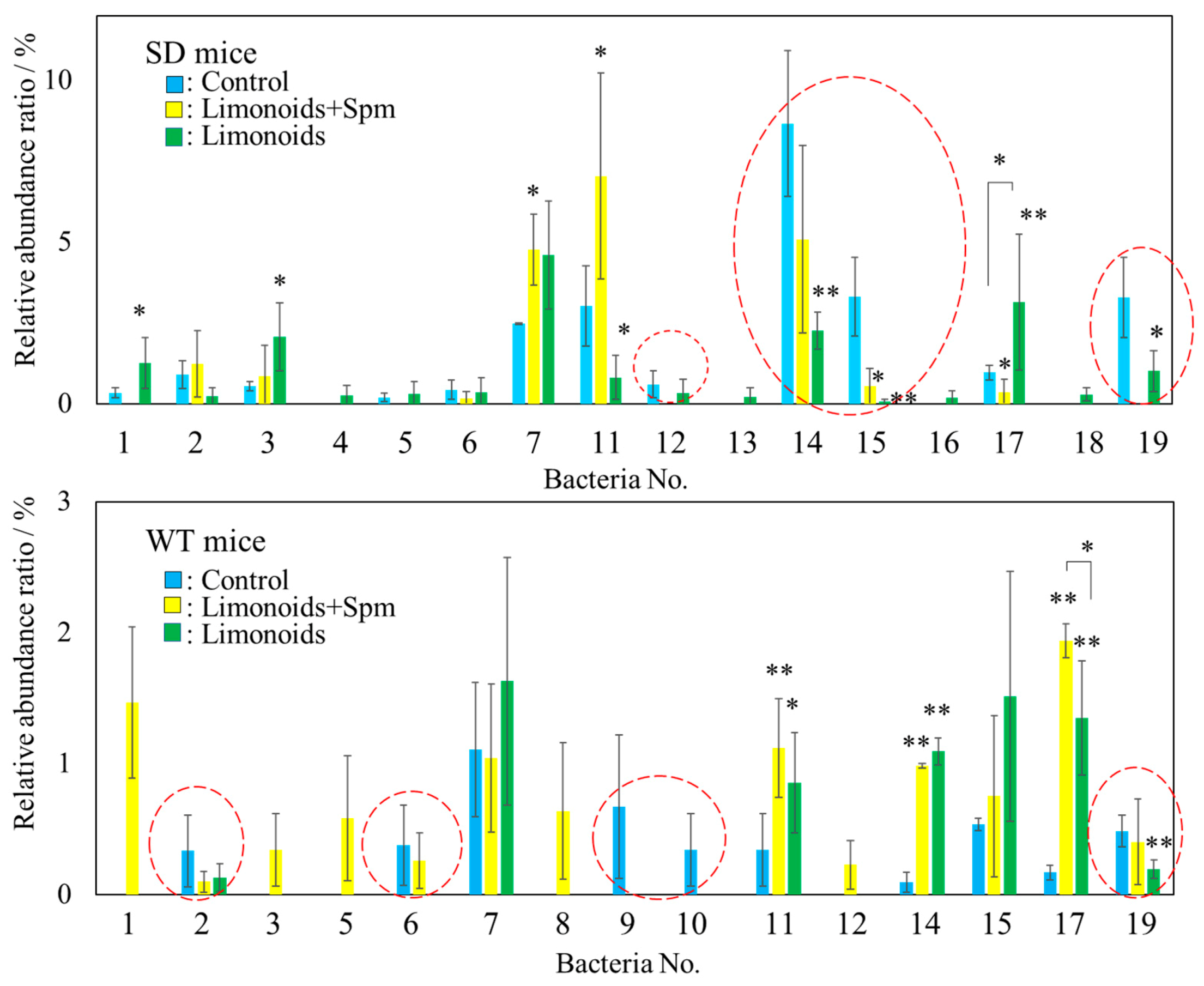
3.3. Composition of Fecal Microbiota in SD and Wild Control (WT) Mice
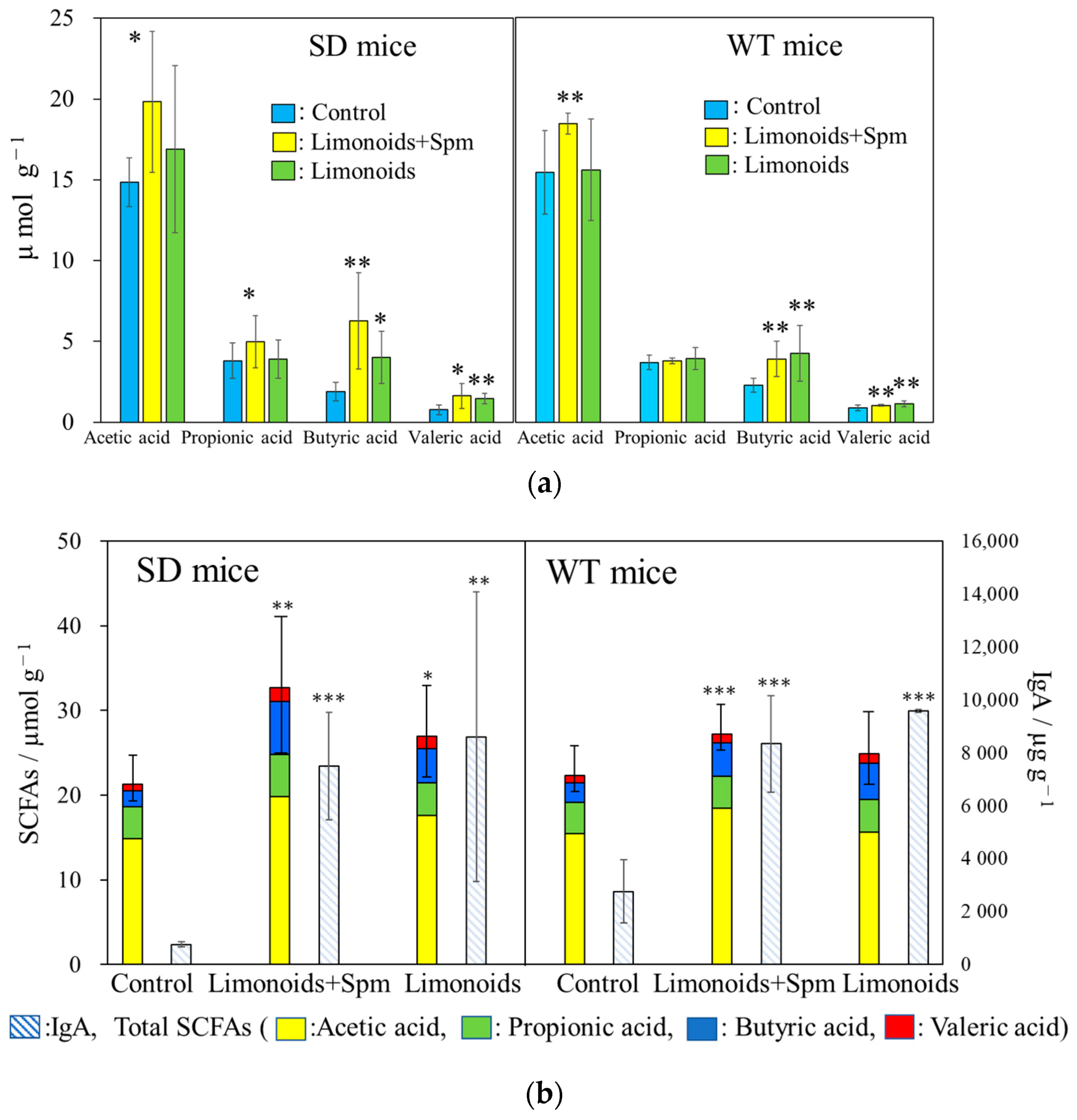
3.4. SCFAs and IgA Productions in SD and WT Mice Feces
3.5. Correlation between Limonoids and/or Spermine and the Clostridiales Group
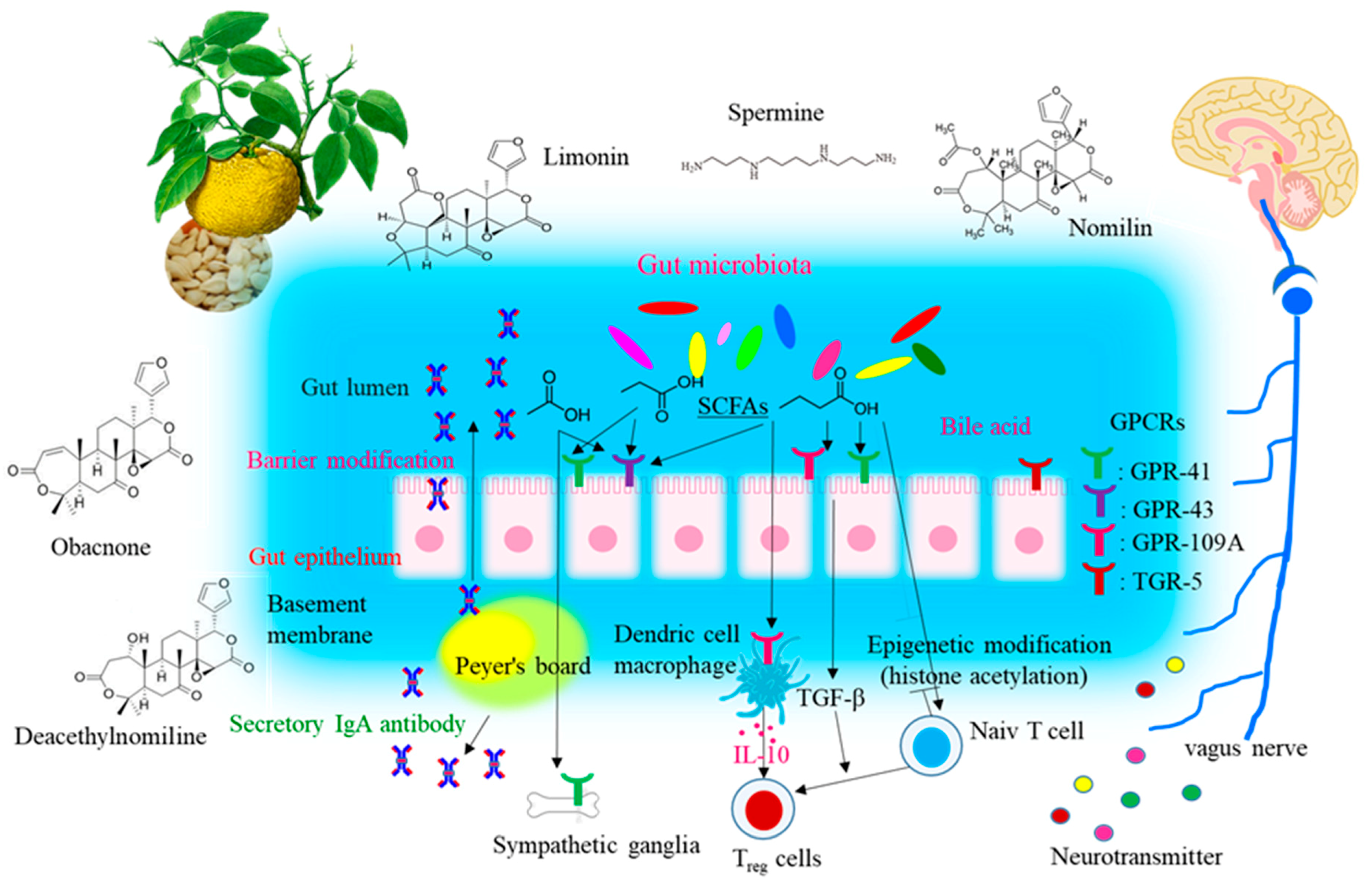
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minamisawa, M.; Suzuki, K.; Kawai, G.; Yamaguchi, A.; Yamanaka, S. Functional evaluation of yuzu (Citrus junos) extracts containing limonoids and polyamine for life extension. J. Funct. Foods 2017, 38, 591–600. [Google Scholar] [CrossRef]
- Huang, J.-Q.; Trasler, J.M.; Igdoura, S.; Michaud, J.; Hanai, N.; Gravel, R.A. Apoptotic Cell Death in Mouse Models of GM2 Gangliosidosis and Observations on Human Tay-Sachs and Sandhoff Diseases. Hum. Mol. Genet. 1997, 6, 1879–1885. [Google Scholar] [CrossRef] [Green Version]
- Yamato, O.; Hayashi, D.; Satoh, H.; Shoda, T.; Uchida, K.; Nakayama, H.; Sakai, H.; Masegi, T.; Murai, A.; Iida, T.; et al. Retrospective Diagnosis of Feline GM2 Gangliosidosis Variant 0 (Sandhoff-Like Disease) in Japan: Possible Spread of the Mutant Allele in the Japanese Domestic Cat Population. J. Vet. Med. Sci. 2008, 70, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Yamaguchi, A.; Yamanaka, S.; Kanzaki, S.; Kawashima, M.; Togo, T.; Katsuse, O.; Koumitsu, N.; Aoki, N.; Iseki, E.; et al. Accumulated α-synuclein affects the progression of GM2 gangliosidoses. Exp. Neurol. 2016, 284, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Sano, T.; Irisa, M.; Kodama, T.; Saito, T.; Furusawa, E.; Kaizu, K.; Yanagi, Y.; Tsukimura, T.; Togawa, T.; et al. FcRγ-dependent immune activation initiates astrogliosis during the asymptomatic phase of Sandhoff disease model mice. Sci. Rep. 2017, 7, 40518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, R.; Tifft, C.J.; Proia, R.L. Microglial activation precedes acute neurodegeneration in Sandhoff disease and is suppressed by bone marrow transplantation. Proc. Natl. Acad. Sci. USA 2000, 97, 10954–10959. [Google Scholar] [CrossRef] [Green Version]
- Jeyakumar, M.; Thomas, R.; Elliot-Smith, E.; Smith, D.A.; Van Der Spoel, A.C.; d’Azzo, A.; Hugh Perry, V.; Butters, T.D.; Dwek, R.A.; Platt, F.M. Central nervous system inflammation is a hallmark of pathogenesis in mouse models of GM1 and GM2 gangliosidosis. Brain 2003, 126, 974–987. [Google Scholar] [CrossRef] [Green Version]
- Horiba, T.; Katsukawa, M.; Mita, M.; Sato, R. Dietary obacunone supplementation stimulates muscle hypertrophy, and suppresses hyperglycemia and obesity through the TGR5 and PPARγ pathway. Biochem. Biophys. Res. Commun. 2015, 463, 846–852. [Google Scholar] [CrossRef]
- Ono, E.; Inoue, J.; Hashidume, T.; Shimizu, M.; Sato, R. Anti-obesity and anti-hyperglycemic effects of the dietary citrus limonoid nomilin in mice fed a high-fat diet. Biochem. Biophys. Res. Commun. 2011, 410, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Sato, R. Nomilin as an Anti-Obesity and Anti-Hyperglycemic Agent. Vitam. Horm. 2013, 91, 425–439. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kurihara, S.; Kibe, R.; Ashida, H.; Benno, Y. Longevity in Mice Is Promoted by Probiotic-Induced Suppression of Colonic Senescence Dependent on Upregulation of Gut Bacterial Polyamine Production. PLoS ONE 2011, 6, e23652. [Google Scholar] [CrossRef] [Green Version]
- Desbonnet, L.; Clarke, G.R.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. Microbiota is essential for social development in the mouse. Mol. Psychiatry 2014, 19, 146–148. [Google Scholar] [CrossRef]
- De Palma, G.; Blennerhassett, P.; Lu, J.; Deng, Y.; Park, A.J.; Green, W.; Denou, E.; Silva, M.A.; Santacruz, A.; Sanz, Y.; et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat. Commun. 2015, 6, 7735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suganya, K.; Koo, B.-S. Gut–Brain Axis: Role of Gut Microbiota on Neurological Disorders and How Probiotics/Prebiotics Beneficially Modulate Microbial and Immune Pathways to Improve Brain Functions. Int. J. Mol. Sci. 2020, 21, 7551. [Google Scholar] [CrossRef] [PubMed]
- Nobuyuki, S. Gut microbiota and brain function. J. Intest. Microbiol. 2017, 31, 23–32. [Google Scholar]
- Andoh, A. Unexpected Functions of the Gut Microbiota as a Virtual Organ. Nihon Naika Gakkai Zasshi 2015, 104, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [Green Version]
- Haghikia, A.; Li, X.S.; Liman, T.G.; Bledau, N.; Schmidt, D.; Zimmermann, F.; Landmesser, U. Gut microbiota-dependent Trimethylamine Noxide predicts risk of cardiovascular events in patients with stroke and is related to Proinflammatory monocytes. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2225–2235. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Sheng, Z.; Zhou, P.; Liu, C.; Zhao, H.; Song, L.; Li, J.; Zhou, J.; Chen, Y.; Wang, L.; et al. Plasma Trimethylamine N-Oxide as a Novel Biomarker for Plaque Rupture in Patients With ST-Segment–Elevation Myocardial Infarction. Circ. Cardiovasc. Interv. 2019, 12, e007281. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Ando, M.; Kamada, N.; Nagano, Y.; Narushima, S.; Suda, W.; Imaoka, A.; Setoyama, H.; Nagamori, T.; et al. Th17 Cell Induction by Adhesion of Microbes to Intestinal Epithelial Cells. Cell 2015, 163, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat. Med. 2015, 21, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Kurakawa, T.; Umemoto, E.; Motooka, D.; Ito, Y.; Gotoh, K.; Hirota, K.; Matsushita, M.; Furuta, Y.; Narazaki, M.; et al. Dysbiosis Contributes to Arthritis Development via Activation of Autoreactive T Cells in the Intestine. Arthritis Rheumatol. 2016, 68, 2646–2661. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Thaiss, C.A.; Zeevi, D.; Dohnalová, L.; Zilberman-Schapira, G.; Mahdi, J.A.; David, E.; Savidor, A.; Korem, T.; Herzig, Y.; et al. Microbiota-Modulated Metabolites Shape the Intestinal Microenvironment by Regulating NLRP6 Inflammasome Signaling. Cell 2015, 163, 1428–1443. [Google Scholar] [CrossRef] [Green Version]
- Hoban, A.E.; Stilling, R.M.; Moloney, G.; Shanahan, F.; Dinan, T.G.; Clarke, G.; Cryan, J.F. The microbiome regulates amygdala-dependent fear recall. Mol. Psychiatry 2018, 23, 1134–1144. [Google Scholar] [CrossRef]
- Erny, D.; De Angelis, A.L.H.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Soto, M.; Herzog, C.; Pacheco, J.A.; Fujisaka, S.; Bullock, K.; Clish, C.B.; Kahn, C.R. Gut microbiota modulate neurobehavior through changes in brain insulin sensitivity and metabolism. Mol. Psychiatry 2018, 23, 2287–2301. [Google Scholar] [CrossRef]
- Descamps, H.C.; Herrmann, B.; Wiredu, D.; Thaiss, C.A. The path toward using microbial metabolites as therapies. EBioMedicine 2019, 44, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Kettle, H.; Louis, P.G.H.; Holtrop, G.; Duncan, S.H.; Flint, H.J. Modelling the emergent dynamics and major metabolites of the human colonic microbiota. Environ. Microbiol. 2015, 17, 1615–1630. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K. Contribution of gut microbiota to the etiology of human diseases. Mod. Media. 2014, 60, 6–18. [Google Scholar]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Tang, H.; Chen, P.; Xie, H.; Tao, Y. Demystifying the manipulation of host immunity, metabolism, and extraintestinal tumors by the gut microbiome. Signal. Transduct. Target. Ther. 2019, 4, 1–34. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome–brain–β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Wu, W.; Chen, L.; Yang, W.; Huang, X.; Ma, C.; Chen, F.; Xiao, Y.; Zhao, Y.; Ma, C.; et al. Microbiota-derived short-chain fatty acids promote Th1 cell IL-10 production to maintain intestinal homeostasis. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Sun, M.; Chen, F. Microbiota metabolite short-chain fatty acid acetate promotes intestinal IgA response to mi-crobiota which is mediated by GPR. Mucosal. Immunol. 2017, 10, 946–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; Van Der Veeken, J.; DeRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell ho-meostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Yoo, D.Y.; Kim, W.; Nam, S.M.; Kim, D.W.; Chung, J.Y.; Choi, S.Y.; Yoon, Y.S.; Won, M.-H.; Hwang, I.K. Synergistic Effects of Sodium Butyrate, a Histone Deacetylase Inhibitor, on Increase of Neurogenesis Induced by Pyridoxine and Increase of Neural Proliferation in the Mouse Dentate Gyrus. Neurochem. Res. 2011, 36, 1850–1857. [Google Scholar] [CrossRef]
- Minamisawa, M.; Yoshida, S.; Uzawa, A. The functional evaluation of waste yuzu (Citrus junos) seeds. Food Funct. 2014, 5, 330–336. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.M.; Sohn, J.; Tyrrell, K.L.; Citron, D.M.; Lawson, P.A.; Patel, N.B.; Iizumi, T.; Perez-Perez, G.I.; Goldstein, E.J.C.; Blaser, M.J. Description of two novel members of the family Erysipelotrichaceae: Ileibacterium valens gen. nov., sp. nov. and Dubosiella newyorkensis, gen. nov., sp. nov., from the murine intestine, and emendation to the description of Faecalibacterium rodentium. Int. J. Syst. Evol. Microbiol. 2017, 67, 1247–1254. [Google Scholar] [CrossRef]
- Honda, K. Clostridia and the mucosal immune system. J. Intest. Microbiol. 2013, 27, 187–196. [Google Scholar]
- Mihara, H.; Suzuki, N.; Muhammad, J.S.; Nanjo, S.; Ando, T.; Fujinami, H.; Kajiura, S.; Hosokawa, A.; Sugiyama, T. Transient receptor potential vanilloid 4 (TRPV4) silencing in Helicobacter pylori-infected human gastric epithelium. Helicobacter 2016, 22, 12361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braak, H.; Del Tredici, K.; Bratzke, H.; Hamm-Clement, J.; Sandmann-Keil, D.; Rüb, U. Staging of the intracerebral inclusion body pathology associated with idiopathic Parkinson’s disease (preclinical and clinical stages). J. Neurol. 2002, 249, 1. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Goto, S.; Tsuji, H.; Okuno, T.; Asahara, T.; Nomoto, K.; Shibata, A.; Fujisawa, Y.; Minato, T.; Okamoto, A.; et al. Intestinal Dysbiosis and Lowered Serum Lipopolysaccharide-Binding Protein in Parkinson’s Disease. PLoS ONE 2015, 10, e0142164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P.; et al. Parkinson’s disease and Parkinson’s disease medications have distinct signatures of the gut microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S.; et al. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull. Exp. Biol. Med. 2017, 162, 734–737. [Google Scholar] [CrossRef]
- Kelly, L.P.; Carvey, P.M.; Keshavarzian, A.; Shannon, K.M.; Shaikh, M.; Bakay, R.A.E.; Kordower, J.H. Progression of intestinal permeability changes and alpha-synuclein expression in a mouse model of Parkinson’s disease. Mov. Disord. 2014, 29, 999–1009. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endo-toxin exposure markers in early Parkinson’s disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [Green Version]
- Forslund, K.; Hildebrand, F.; Nielsen, T.R.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Pedersen, H.K.; et al. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The gut microbiota mediates the an-ti-seizure effects of the ketogenic diet. Cell 2018, 173, 1728–1741.e13. [Google Scholar] [CrossRef] [Green Version]
- Okumura, R.; Kurakawa, T.; Nakano, T.; Kayama, H.; Kinoshita, M.; Motooka, D.; Gotoh, K.; Kimura, T.; Kamiyama, N.; Kusu, T.; et al. Lypd8 promotes the segregation of flagellated microbiota and colonic epithelia. Nat. Cell Biol. 2016, 532, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Jean-Félix, S.; Guillaume, L.B.; Philippe, V.; Mario, J.; Josée, H. Interactions of Intestinal Bacteria with Components of the Intestinal Mucus. Front. Cell Infect Microbiol. 2017, 7, 387. [Google Scholar] [CrossRef]
- Turroni, F.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Viappiani, A.; Lugli, G.A.; Ferrario, C.; Gioiosa, L.; Ferrarini, A.; et al. Deciphering bifidobacterial-mediated metabolic interactions and their impact on gut microbiota by a multi-omics approach. ISME J. 2016, 10, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M.; et al. TGR5-Mediated Bile Acid Sensing Controls Glucose Homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nature Reviews. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar]
- Sakamoto, M. The taxonomy of the genus Bacteroides and related taxa. J. Intest. Microbiol. 2016, 30, 119–127. [Google Scholar]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, D.; Kimura, I.; Wakabayashi, M.; Tsumoto, H.; Ozawa, K.; Hara, T.; Takei, Y.; Hirasawa, A.; Ishihama, Y.; Tsujimoto, G. Short-chain fatty acid receptor GPR41-mediated activation of sympathetic neurons involves synapsin 2b phosphorylation. FEBS Lett. 2012, 586, 1547–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNelis, J.C.; Lee, Y.S.; Mayoral, R.; Van Der Kant, R.; Johnson, A.M.F.; Wollam, J.; Olefsky, J.M. GPR43 Potentiates β-Cell Function in Obesity. Diabetes 2015, 64, 3203–3217. [Google Scholar] [CrossRef] [Green Version]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.I.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fiber-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbondio, M.; Palomba, A.; Tanca, A.; Fraumene, C.; Pagnozzi, D.; Serra, M.; Marongiu, F.; Laconi, E.; Uzzau, S. Fecal Metaproteomic Analysis Reveals Unique Changes of the Gut Microbiome Functions After Consumption of Sourdough Carasau Bread. Front. Microbiol. 2019, 10, 1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Tsuji, M.; Komatsu, N.; Kawamoto, S.; Suzuki, K.; Kanagawa, O.; Honjo, T.; Hori, S.; Fagarasan, S. Preferential Generation of Follicular B Helper T Cells from Foxp3+ T Cells in Gut Peyer’s Patches. Science 2009, 323, 1488–1492. [Google Scholar] [CrossRef] [PubMed]
- Littman, D.R.; Rudensky, A.Y. Th17 and regulatory T cells in mediating and restraining inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaldson, G.P.; Ladinsky, M.S.; Yu, K.B.; Sanders, J.G.; Yoo, B.B.; Chou, W.-C.; Conner, M.E.; Earl, A.M.; Knight, R.; Bjorkman, P.J.; et al. Gut microbiota utilize immunoglobulin A for mucosal colonization. Science 2018, 360, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Magri, G.; Grasset, E.K.; Cerutti, A. Rethinking mucosal antibody responses: IgM, IgG and IgD join IgA. Nat. Rev. Immunol. 2020, 20, 427–441. [Google Scholar] [CrossRef]
- Yanagibashi, T.; Hosono, A.; Oyama, A.; Tsuda, M.; Suzuki, A.; Hachimura, S.; Takahashi, Y.; Momose, Y.; Itoh, K.; Hirayama, K.; et al. IgA production in the large intestine is modulated by a different mechanism than in the small intestine: Bacteroides acidifaciens promotes IgA production in the large intestine by inducing germinal center formation and increasing the number of IgA+ B cells. Immunobiology 2013, 218, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Vogelzang, A.; Maruya, M.; Miyajima, M.; Murata, M.; Son, A.; Kuwahara, T.; Tsuruyama, T.; Yamada, S.; Matsuura, M.; et al. IgA regulates the composition and metabolic function of gut microbiota by promoting symbiosis between bacteria. J. Exp. Med. 2018, 215, 2019–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, O.L.; Pröbstel, A.K.; Porfilio, E.A.; Wang, A.A.; Charabati, M.; Sun, T.; Gommerman, J.L. Recirculating Intestinal IgA-Producing Cells Regulate Neuroinflammation via IL-10. Cell 2019, 176, 610–624.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus Classification | Relative Abundance of Bacterial (%) | |||||
|---|---|---|---|---|---|---|
| SD Mice | WT Mice | |||||
| Control | Limonoids + Spm | Limonoids | Control | Limonoids + Spm | Limonoids | |
| No. Firmicutes–Clostridiales–Lachnospiraceae family | ||||||
| 1. Lachnoclostridium | 0.33 ± 0.15 | nd. | 1.3 ± 0.79 * | nd. | 1.5 ± 0.58 | nd. |
| 2. Lachnospiraceae FCS020 group | 0.91 ± 0.42 | 1.2 ± 1.0 | 0.22 ± 0.27 | 0.34 ± 0.27 | 0.10 ± 0.08 | 0.13 ± 0.10 |
| 3. Lachnospiraceae NK4A136 group | 0.54 ± 0.15 | 0.84 ± 0.96 | 2.1 ± 1.1 * | nd. | 0.34 ± 0.20 | nd. |
| 4. Lachnospiraceae UCG-004 | nd. | nd. | 0.26 ± 0.31 | nd. | nd. | nd. |
| 5. Lachnospiraceae UCG-006 | 0.19 ± 0.14 | nd. | 0.31 ± 0.38 | nd. | 0.58 ± 0.48 | nd. |
| 6. Uncultured (Lachnospiraceae) | 0.43 ± 0.31 | 0.16 ± 0.22 | 0.36 ± 0.44 | 0.37 ± 0.30 | 0.26 ± 0.21 | nd. |
| 7. Lachnospiraceae | 2.5 ± 0.018 | 4.8 ± 1.1 * | 4.6 ± 1.7 * | 1.1 ± 0.51 | 1.0 ± 0.56 | 1.6 ± 0.94 |
| 8. Eubacterium fissicatena group | nd. | nd. | nd. | nd. | 0.64 ± 0.52 | nd. |
| Firmicutes–Clostridiales–Peptostreptococcaceae family | ||||||
| 9. Romboutsia | nd. | nd. | nd. | 0.67 ± 0.55 | nd. | nd. |
| 10. Peptostrepto- coccaceae | nd. | nd. | nd. | 0.34 ± 0.28 | nd. | nd. |
| Firmicutes–Clostridiales–Ruminococcaceae family | ||||||
| 11. Ruminiclostridia- um 5 | 3.0 ± 1.2 | 7.0 ± 3.2 ** | 0.81 ± 0.68 | 0.34 ± 0.28 | 1.1 ± 0.34 | 0.85 ± 0.38 |
| 12. Rumini- clostridium 9 | 0.60 ± 0.42 | 0.34 ± 0.41 | nd. | nd. | 0.23 ± 0.19 | nd. |
| 13. Ruminococcaceae UCG-009 | nd. | nd. | 0.22 ± 0.27 | nd. | nd. | nd. |
| 14. Ruminococcaceae UCG-014 | 8.7 ± 2.2 | 5.1 ± 2.9 | 2.3 ± 0.58 | 0.09 ± 0.08 | 0.98 ± 0.02 | 1.1 ± 0.10 |
| 15. Ruminococcus 1 | 3.3 ± 1.2 | 0.54 ± 0.56 | 0.062 ± 0.07 | 0.53 ± 0.05 | 0.75 ± 0.61 | 1.5 ± 0.95 |
| 16. UBA1819 (Ruminococcaceae) | nd. | nd. | 0.18 ± 0.22 | nd. | nd. | nd. |
| 17. Eubacteriumcopro-stanoligenes group | 0.96 ± 0.23 | 0.35 ± 0.39 | 3.1 ± 2.1 | 0.17 ± 0.05 | 1.9 ± 0.13 | 1.3 ± 0.43 |
| 18. Uncultured (Ruminococcaceae) | nd. | nd. | 0.29 ± 0.20 | nd. | nd. | 0.29 ± 0.20 |
| 19. Ruminococcaceae | 3.3 ± 1.2 | nd. | 1.0 ± 0.62 | 0.49 ± 0.12 | 0.40 ± 0.33 | 0.19 ± 0.07 |
| Firmicutes–Lactobacillales–Lactobacillaceae family | ||||||
| Lactobacillus | 47 ± 11 | 39 ± 12 | 7.4 ± 1.8 ** | 4.2 ± 1.5 | 18 ± 5.7 ** | 31 ± 9.8 ** |
| Lactococcus | nd. | 1.7 ± 2.0 | 0.30 ± 0.37 | nd. | 1.5 ± 1.2 | 0.19 ± 0.16 |
| Firmicutes–Erysipelotrichales–Erysipelotrichaceae family | ||||||
| Candidatus Stoquefichus | nd. | 2.9 ± 3.4 | nd. | nd. | 0.78 ± 0.33 | nd. |
| Dubosiella | nd. | nd. | 2.4 ± 1.4 | 16 ± 1.9 | 1.7 ± 1.4 *** | 1.9 ± 1.5 *** |
| Erysipelatoclostridium | nd. | 0.28 ± 0.39 | nd. | nd. | 0.29 ± 0.23 | nd. |
| Turicibacter | 4.8 ± 1.9 | nd. | 4.5 ± 2.1 | 50 ± 6.3 | 8.5 ± 4.0 *** | 13 ± 6.2 *** |
| Erysipelotrichaceae | 0.06 ± 0.04 | 1.7 ± 2.4 | 1.2 ± 0.92 | nd. | 0.23 ± 0.18 | 1.3 ± 0.55 |
| Patescibacteria–Saccharimonadales–Saccharimonadaceae family | ||||||
| Candidatus Saccharimonas | 3.8 ± 2.7 | 1.2 ± 1.6 | 0.22 ± 0.27 | 1.8 ± 1.4 | nd. | 0.68 ± 0.16 |
| Proteobacteria–Desulfovibrionales–Desulfovibrionaceae family | ||||||
| Desulfovibrio | 1.9 ± 0.53 | 1.1 ± 1.5 | 3.4 ± 3.9 | nd. | 0.75 ± 0.60 | 0.38 ± 0.03 |
| Verrucomicrobia–Verrucomicrobiales–Akkermansiaceae family | ||||||
| a Akkermansia | nd. | 1.1 ± 1.3 | 0.31 ± 0.27 | 0.37 ± 0.30 | 1.6 ± 1.3 * | 3.0 ± 2.0 *** |
| Bacteroidetes–Bacteroidales–Bacteroidaceae family | ||||||
| a Bacteroides | 1.2 ± 0.076 | 0.51 ± 0.36 | 8.4 ± 4.5 * | 0.14 ± 0.11 | 0.21 ± 0.13 | 3.0 ± 0.02 |
| Bacteroidetes–Bacteroidales–Muribaculaceae family | ||||||
| Muribaculum | 0.90 ± 0.64 | 1.6 ± 2.3 ** | 5.1 ± 5.8 *** | 0.90 ± 0.73 | 0.35 ± 0.26 | 15 ± 3.4 *** |
| Uncultured Bacteroidales bacterium | nd. | nd. | nd. | nd. | 0.26 ± 0.09 | nd. |
| Uncultured Barnesiella sp. | nd. | nd. | nd. | nd. | nd. | 0.41 ± 0.34 |
| Uncultured Bacterium | 6.9 ± 2.7 | 26 ± 3.8 ** | 28 ± 9.5 *** | 8.0 ± 5.5 | 38 ± 2.9 *** | 5.1 ± 0.53 |
| Uncultured organism | nd. | nd. | nd. | 1.0 ± 0.82 | 4.8 ± 0.76 | nd. |
| Muribaculaceae | 2.4 ± 0.097 | 6.6 ± 0.27 | 7.5 ± 2.3 | 4.4 ± 1.6 | 6.8 ± 0.22 | 11 ± 2.7 ** |
| Bacteroidetes–Bacteroidales–Prevotellaceae family | ||||||
| a Alloprevotella | 1.2 ± 0.37 | 1.6 ± 0.63 | 11 ± 2.7 | 0.30 ± 0.24 | 3.0 ± 0.56 | 4.0 ± 1.6 |
| a Paraprevotella | nd. | nd. | 0.18 ± 0.11 | nd. | 0.10 ± 0.08 | 0.53 ± 0.34 |
| a Prevotellaceae NK3B31 group | nd. | nd. | 0.089 ± 0.11 | nd. | 0.17 ± 0.14 | nd. |
| a Prevotellaceae | nd. | nd. | 0.65 ± 0.80 | nd. | nd. | 0.28 ± 0.23 |
| Bacteroidetes–Bacteroidales–Tannerellaceae family | ||||||
| a Parabacteroides | nd. | nd. | 5.4 ± 1.5 | nd. | 0.73 ± 0.57 | 0.58 ± 0.21 |
| Actinobacteria–Bifidobacteriales–Bifidobacteriaceae family | ||||||
| a Bifidobacterium | nd. | nd. | nd. | 7.2 ± 0.03 | nd. | nd. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minamisawa, M.; Suzumura, T.; Bose, S.; Taniai, T.; Kawai, G.; Suzuki, K.; Yamaguchi, A.; Yamanaka, S. Effect of Yuzu (Citrus junos) Seed Limonoids and Spermine on Intestinal Microbiota and Hypothalamic Tissue in the Sandhoff Disease Mouse Model. Med. Sci. 2021, 9, 17. https://doi.org/10.3390/medsci9010017
Minamisawa M, Suzumura T, Bose S, Taniai T, Kawai G, Suzuki K, Yamaguchi A, Yamanaka S. Effect of Yuzu (Citrus junos) Seed Limonoids and Spermine on Intestinal Microbiota and Hypothalamic Tissue in the Sandhoff Disease Mouse Model. Medical Sciences. 2021; 9(1):17. https://doi.org/10.3390/medsci9010017
Chicago/Turabian StyleMinamisawa, Mayumi, Takuma Suzumura, Sudeep Bose, Tetsuyuki Taniai, Gota Kawai, Kyoko Suzuki, Akira Yamaguchi, and Shoji Yamanaka. 2021. "Effect of Yuzu (Citrus junos) Seed Limonoids and Spermine on Intestinal Microbiota and Hypothalamic Tissue in the Sandhoff Disease Mouse Model" Medical Sciences 9, no. 1: 17. https://doi.org/10.3390/medsci9010017
APA StyleMinamisawa, M., Suzumura, T., Bose, S., Taniai, T., Kawai, G., Suzuki, K., Yamaguchi, A., & Yamanaka, S. (2021). Effect of Yuzu (Citrus junos) Seed Limonoids and Spermine on Intestinal Microbiota and Hypothalamic Tissue in the Sandhoff Disease Mouse Model. Medical Sciences, 9(1), 17. https://doi.org/10.3390/medsci9010017