
Symptom-Dependent Changes in MEG-Derived Neuroelectric Brain Activity in Traumatic Brain Injury Patients with Chronic Symptoms

Abstract
:1. Introduction
2. Results
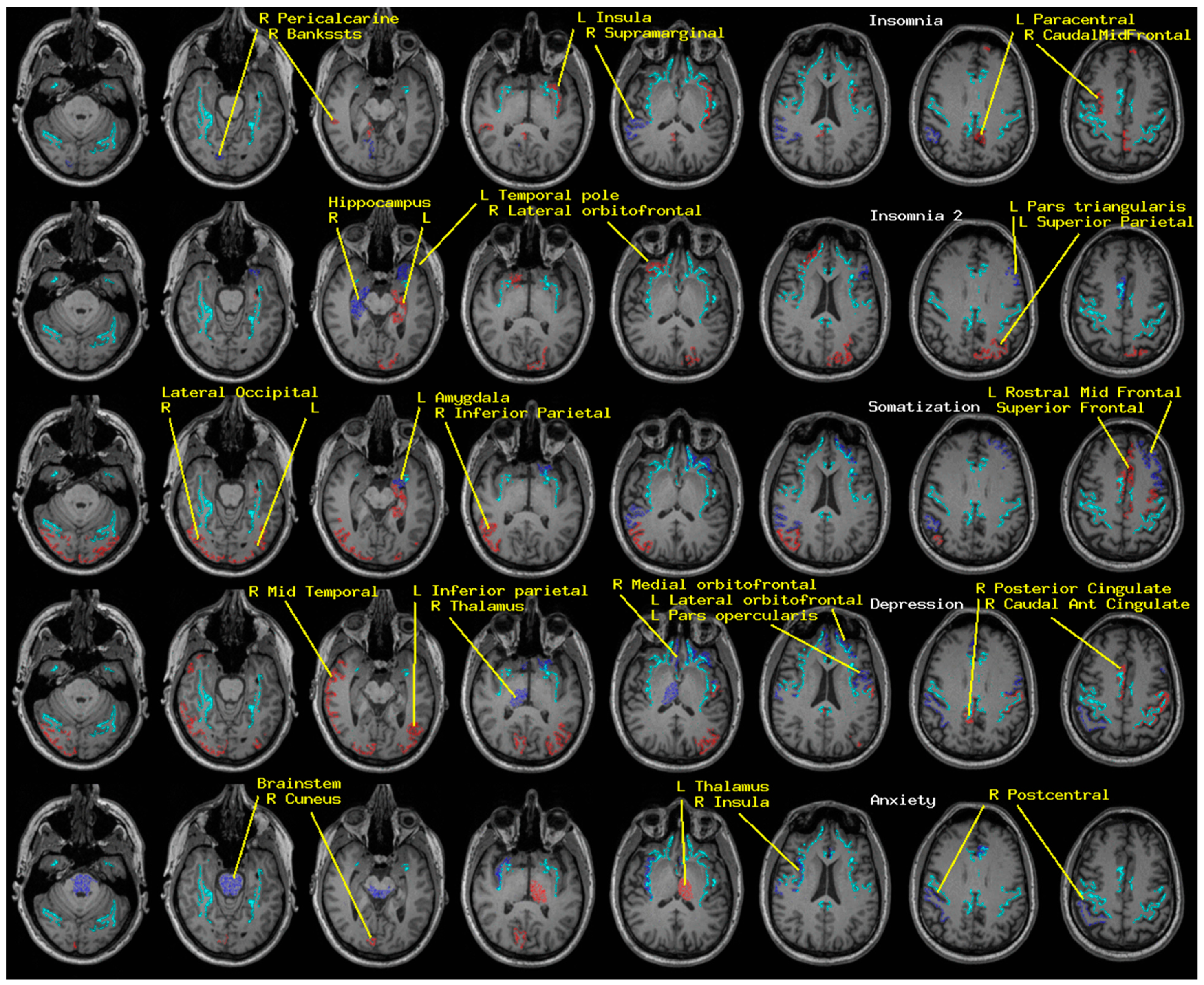
2.1. Insomnia and Psychological Distress
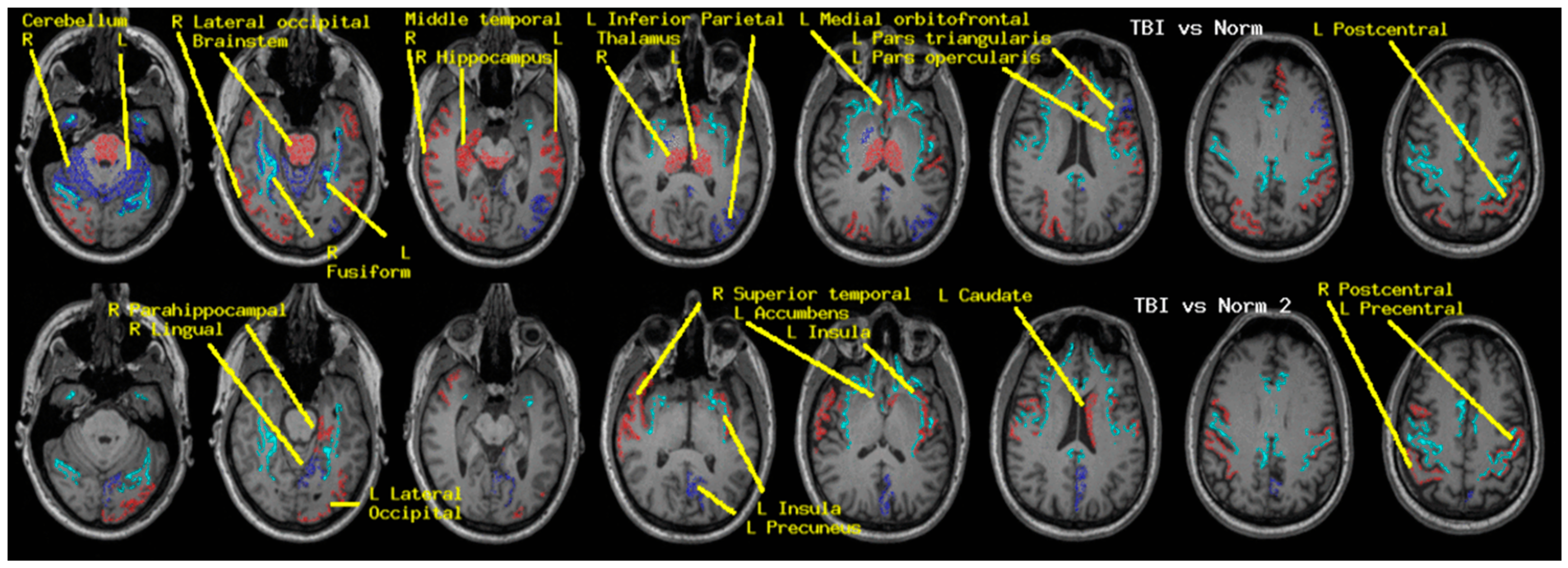
2.2. CamCAN vs. TEAM-TBI Cohort
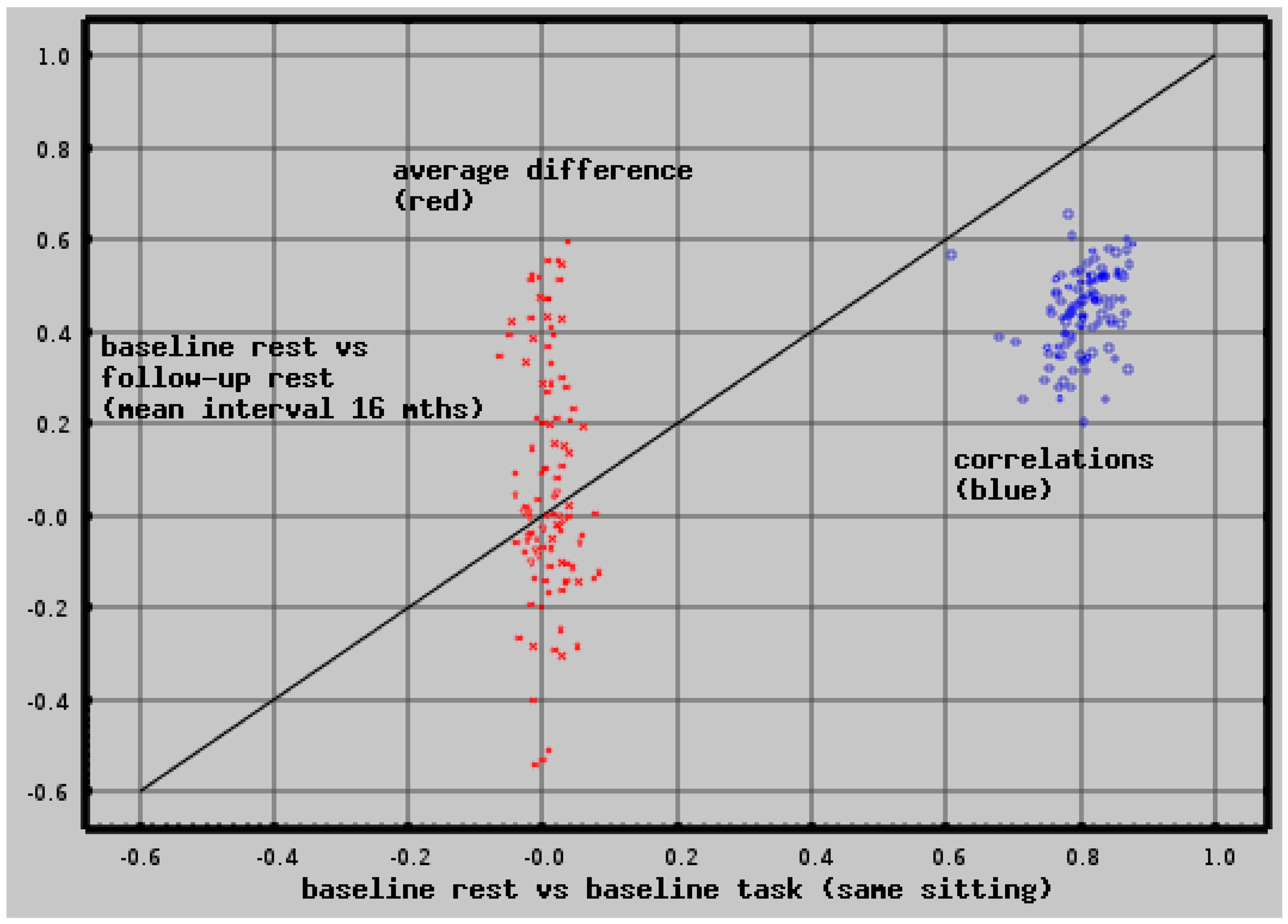
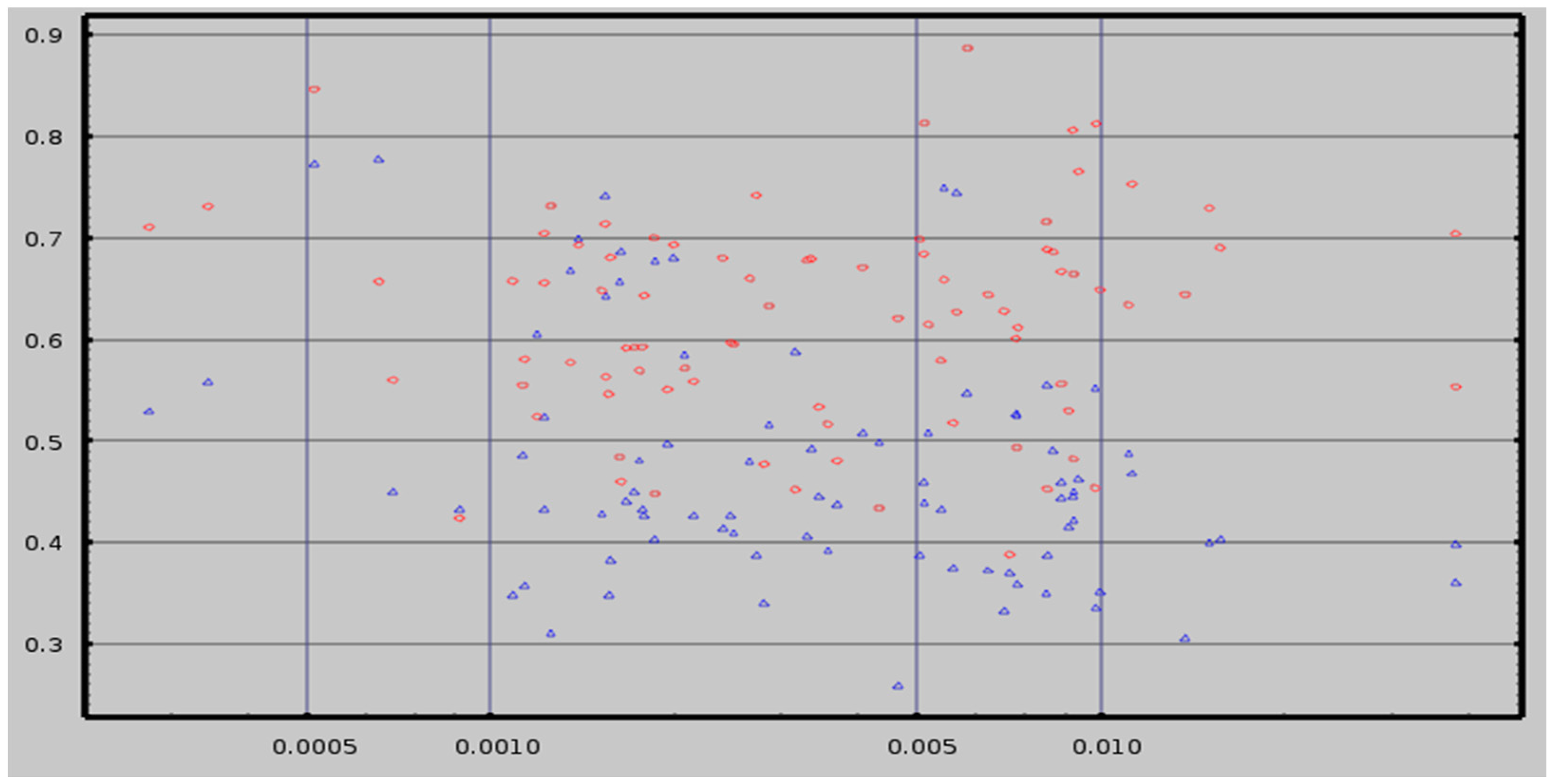
2.3. Test-Retest Reliability
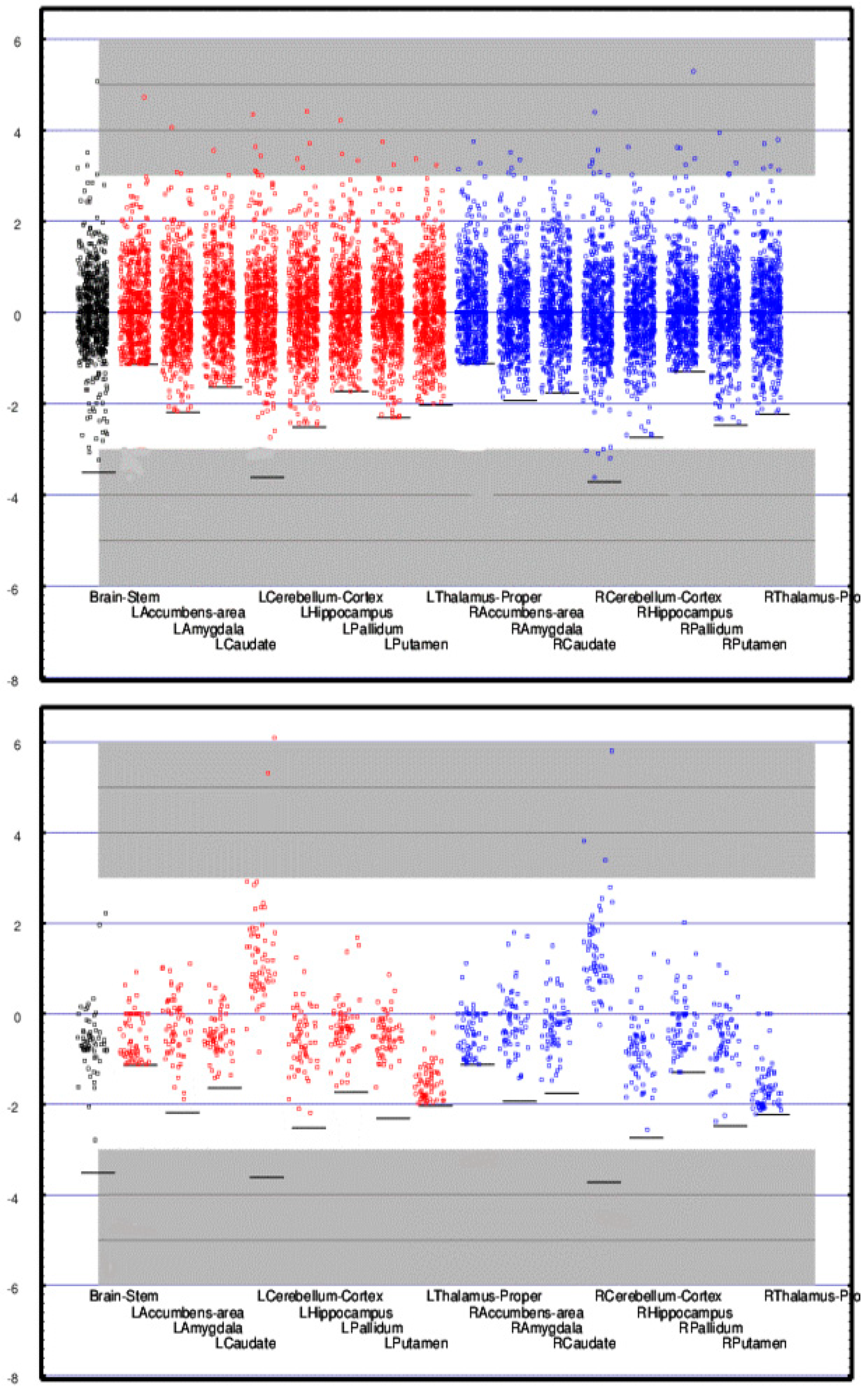
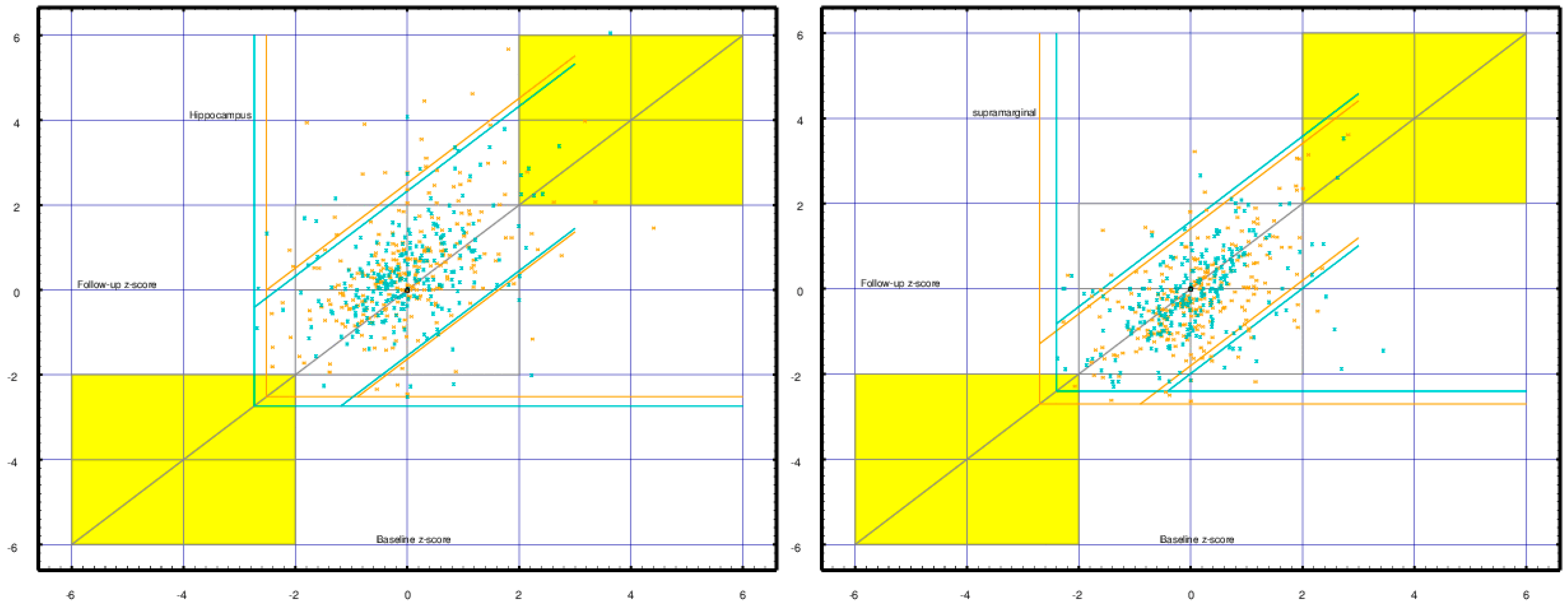
2.4. Differential Activity: Cortical vs. Adjacent White Matter Regions
3. Materials and Methods
3.1. CamCAN Dataset
3.2. TEAM-TBI Dataset
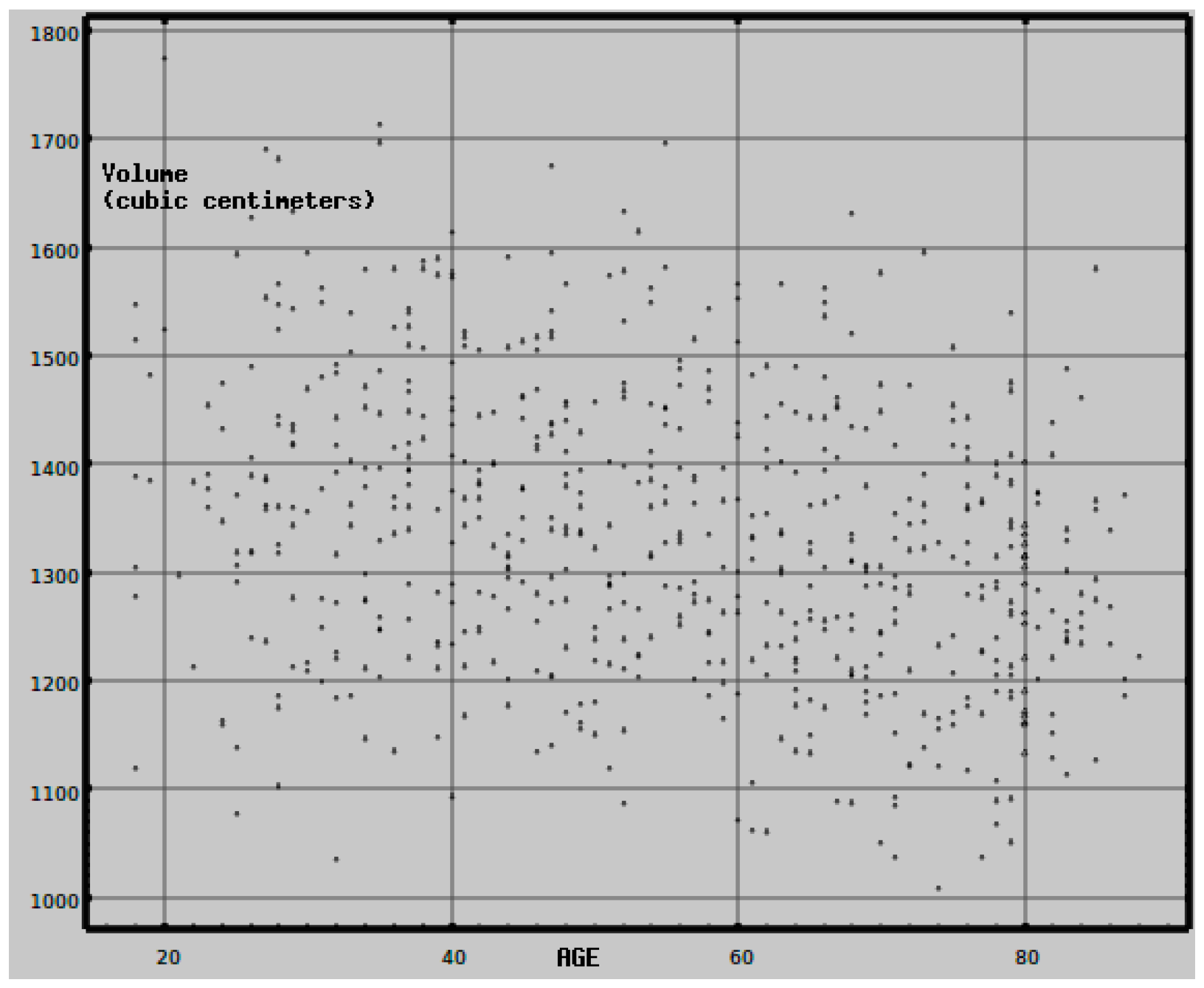
3.3. MRI Processing
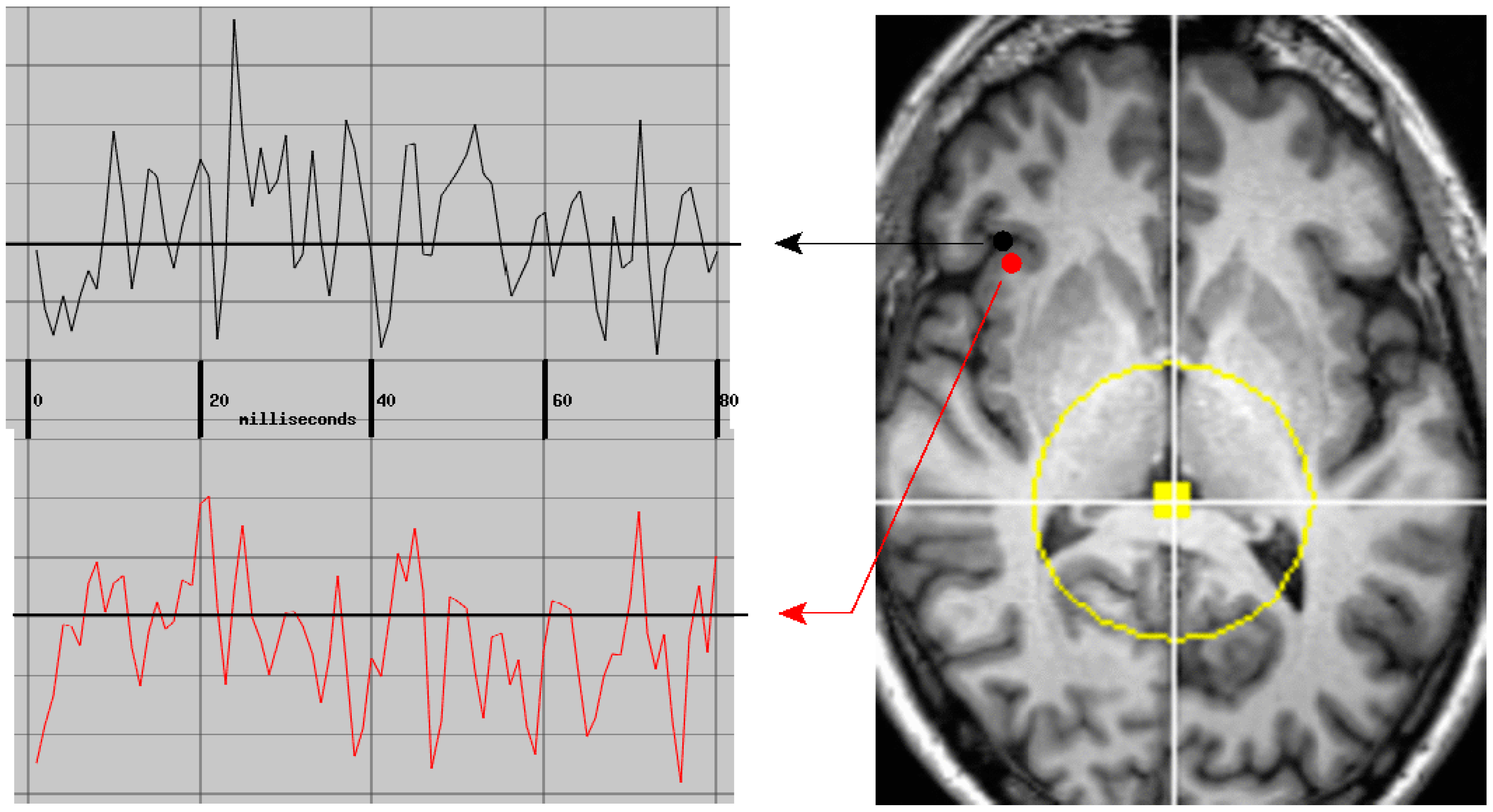
3.4. MEG Processing
4. Normative Measures
Empty Room Correction
5. Classification
6. Discussion
6.1. Potential Clinical Utility
6.2. Test-Retest Reliability
6.3. Cortical vs. Adjacent White Matter Regions
6.4. CamCAN vs. TEAM-TBI Differences
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Atlas of Normative Regional Neuroelectric Measures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Deep White Matter Tracts CamCAN Baseline (n = 589) | Baseline vs. Follow-up (n = 240) Correlation and Mean Difference | |||||||
|---|---|---|---|---|---|---|---|---|
| mean | s.d. | corr | diff | |||||
| 0.319 | 0.168 | corpus callosum splenium (fmajor) | 0.255 | −0.055 | ||||
| 0.561 | 0.234 | corpus callosum genu (fminor) | 0.345 | −0.102 | ||||
| Left | Right | Left | Right | |||||
| mean | s.d. | mean | s.d. | corr | diff | corr | diff | |
| 0.924 | 0.275 | 0.828 | 0.278 | anterior thalamic radiations (atr) | 0.427 | −0.001 | 0.348 | 0.050 |
| 0.914 | 0.304 | 1.039 | 0.325 | cingulum angular bundle (cab) | 0.468 | 0.044 | 0.507 | −0.085 |
| 0.477 | 0.245 | 0.511 | 0.259 | cingulate gyrus endings (ccg) | 0.366 | −0.059 | 0.435 | −0.021 |
| 1.004 | 0.227 | 0.878 | 0.217 | cortico-spinal tract (cst) | 0.471 | 0.041 | 0.369 | −0.052 |
| 0.958 | 0.254 | 0.796 | 0.253 | anterior thalamic radiations (atr) | 0.589 | −0.012 | 0.532 | −0.030 |
| 0.826 | 0.260 | 0.856 | 0.254 | superior longitudinal fasciculus, parietal (slfp) | 0.515 | −0.003 | 0.576 | 0.008 |
| 0.980 | 0.197 | 1.020 | 0.236 | superior longitudinal fasciculus, temporal (slft) | 0.497 | 0.006 | 0.408 | −0.073 |
| 1.087 | 0.288 | 0.947 | 0.288 | uncinate fasciculus (unc) | 0.523 | −0.000 | 0.524 | −0.046 |
| Subcortical Regions CamCAN Baseline (n = 619) | Baseline vs. Follow-up (n = 253) Correlation and Mean Difference | |||||||
|---|---|---|---|---|---|---|---|---|
| mean | s.d. | corr | diff | |||||
| 0.713 | 0.203 | brainstem | 0.434 | −0.510 | ||||
| Left | Right | Left | Right | |||||
| mean | s.d. | mean | s.d. | corr | diff | corr | diff | |
| 0.746 | 0.658 | 0.635 | 0.566 | accumbens | 0.388 | 0.157 | 0.349 | 0.136 |
| 1.057 | 0.483 | 0.921 | 0.476 | amygdala | 0.523 | 0.474 | 0.560 | 0.513 |
| 0.569 | 0.347 | 0.580 | 0.330 | caudate | 0.488 | 0.022 | 0.609 | −0.050 |
| 0.666 | 0.184 | 0.697 | 0.187 | cerebellum | 0.441 | 0.207 | 0.546 | 0.004 |
| 0.876 | 0.348 | 0.985 | 0.360 | hippocampus | 0.456 | 0.524 | 0.476 | 0.408 |
| 0.957 | 0.552 | 0.715 | 0.552 | pallidum | 0.488 | 0.287 | 0.523 | 0.333 |
| 0.944 | 0.409 | 1.062 | 0.429 | putamen | 0.509 | 0.194 | 0.656 | 0.287 |
| 0.622 | 0.306 | 0.730 | 0.328 | thalamus | 0.520 | 0.146 | 0.549 | −0.079 |
| Cortical Regions CamCAN Baseline (n = 619) | Baseline vs. Follow-up (n = 253) Correlation and Mean Difference | |||||||
|---|---|---|---|---|---|---|---|---|
| Left | Right | Left | Right | |||||
| mean | s.d. | mean | s.d. | corr | diff | corr | diff | |
| 0.961 | 0.415 | 0.903 | 0.442 | bankssts | 0.538 | 0.203 | 0.523 | 0.091 |
| 0.565 | 0.426 | 0.547 | 0.156 | caudalanteriorcingulate | 0.252 | −0.142 | 0.377 | −0.200 |
| 0.729 | 0.408 | 0.848 | 0.448 | caudalmiddlefrontal | 0.434 | 0.210 | 0.459 | 0.231 |
| 0.276 | 0.276 | 0.287 | 0.275 | cuneus | 0.253 | −0.134 | 0.355 | −0.042 |
| 1.015 | 0.405 | 0.954 | 0.419 | entorhinal | 0.393 | 0.422 | 0.532 | 0.345 |
| 0.124 | 0.310 | 0.117 | 0.313 | frontalpole | 0.567 | −0.169 | 0.418 | −0.137 |
| 0.578 | 0.229 | 0.611 | 0.243 | fusiform | 0.419 | 0.395 | 0.420 | 0.331 |
| 0.661 | 0.268 | 0.553 | 0.234 | inferiorparietal | 0.426 | −0.068 | 0.530 | −0.284 |
| 1.041 | 0.277 | 0.988 | 0.284 | inferiortemporal | 0.520 | 0.556 | 0.521 | 0.471 |
| 0.697 | 0.267 | 0.652 | 0.274 | insula | 0.581 | 0.213 | 0.508 | 0.428 |
| 0.227 | 0.286 | 0.263 | 0.298 | isthmuscingulate | 0.318 | −0.002 | 0.337 | −0.112 |
| 0.889 | 0.263 | 0.855 | 0.261 | lateraloccipital | 0.577 | −0.432 | 0.472 | 0.369 |
| 0.952 | 0.300 | 0.907 | 0.320 | lateralorbitofrontal | 0.478 | 0.517 | 0.471 | 0.386 |
| 0.470 | 0.260 | 0.497 | 0.253 | lingual | 0.473 | −0.020 | 0.520 | −0.284 |
| 0.517 | 0.263 | 0.537 | 0.269 | medialorbitofrontal | 0.337 | −0.140 | 0.384 | −0.531 |
| 0.716 | 0.256 | 0.862 | 0.252 | middletemporal | 0.4675 | 0.546 | 0.601 | 0.471 |
| 0.335 | 0.290 | 0.334 | 0.274 | paracentral | 0.317 | −0.146 | 0.422 | −0.123 |
| 0.886 | 0.378 | 0.909 | 0.386 | parahippocampal | 0.410 | 0.394 | 0.372 | 0.301 |
| 0.845 | 0.355 | 0.843 | 0.389 | parsopercularis | 0.431 | −0.161 | 0.354 | −0.112 |
| 0.506 | 0.362 | 0.462 | 0.335 | parsorbitalis | 0.525 | 0.152 | 0.342 | 0.104 |
| 0.803 | 0.362 | 0.882 | 0.381 | parstriangularis | 0.443 | −0.033 | 0.415 | −0.038 |
| 0.352 | 0.297 | 0.359 | 0.302 | pericalcarine | 0.490 | 0.109 | 0.292 | 0.036 |
| 0.784 | 0.276 | 0.712 | 0.264 | postcentral | 0.466 | −0.060 | 0.399 | 0.019 |
| 0.382 | 0.356 | 0.403 | 0.324 | posteriorcingulate | 0.316 | −0.102 | 0.319 | −0.290 |
| 0.793 | 0.281 | 0.777 | 0.287 | precentral | 0.459 | 0.092 | 0.480 | 0.198 |
| 0.150 | 0.154 | 0.160 | 0.165 | precuneus | 0.205 | −0.305 | 0.365 | −0.248 |
| 0.730 | 0.438 | 0.701 | 0.419 | rostralanteriorcingulate | 0.278 | −0.401 | 0.280 | −0.540 |
| 0.560 | 0.276 | 0.559 | 0.296 | rostralmiddlefrontal | 0.450 | 0.081 | 0.394 | 0.102 |
| 0.530 | 0.258 | 0.512 | 0.262 | superiorfrontal | 0.442 | 0.005 | 0.457 | −0.143 |
| 0.618 | 0.273 | 0.612 | 0.289 | superiorparietal | 0.442 | −0.105 | 0.447 | −0.071 |
| 0.892 | 0.232 | 0.902 | 0.255 | superiortemporal | 0.468 | 0.596 | 0.513 | 0.430 |
| 0.768 | 0.284 | 0.731 | 0.304 | supramarginal | 0.524 | −0.264 | 0.482 | −0.194 |
| 0.642 | 0.387 | 0.856 | 0.403 | temporalpole | 0.351 | 0.512 | 0.438 | 0.555 |
| 0.808 | 0.450 | 0.832 | 0.503 | transversetemporal | 0.521 | 0.267 | 0.293 | 0.280 |
Appendix B. The Referee Consensus Method
Appendix C: The Referee Consensus Method—Advantages
References
- Telkes, I.; Jimenez-Shahed, J.; Viswanathan, A.; Abosch, A.; Ince, N.F. Prediction of STN-DBS Electrode Implantation Track in Parkinson’s Disease by Using Local Field Potentials. Front. Neurosci. 2016, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Moroni, F.; Nobili, L.; Curcio, G.; De Carli, F.; Fratello, F.; Marzano, C.; De Gennaro, L.; Ferrillo, F.; Cossu, M.; Francione, S.; et al. Sleep in the Human Hippocampus: A Stereo-EEG Study. PLoS ONE 2007, 2, e867. [Google Scholar] [CrossRef] [Green Version]
- Sarvas, J. Basic mathematical and electromagnetic concepts of the biomagnetic inverse problem. Phys. Med. Biol. 1987, 32, 11–22. [Google Scholar] [CrossRef]
- Cheyne, D.; Weinberg, H. Neuromagnetic fields accompanying unilateral finger movements: Pre-movement and movement-evoked fields. Exp. Brain Res. 1989, 78, 604–612. [Google Scholar] [CrossRef]
- Harrop, R.; Weinberg, H.; Brickett, P.; Dykstra, C.; Robertson, A.; Cheyne, D.; Baff, M.; Crisp, D. The biomagnetic inverse problem: Some theoretical and practical considerations. Phys. Med. Biol. 1987, 32, 1545–1557. [Google Scholar] [CrossRef]
- Hämäläinen, M.S.; Ilmoniemi, R.J. Interpreting magnetic fields of the brain: Minimum norm estimates. Med Biol. Eng. Comput. 1994, 32, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Papadelis, C.; Leonardelli, E.; Staudt, M.; Braun, C. Can magnetoencephalography track the afferent information flow along white matter thalamo-cortical fibers? NeuroImage 2012, 60, 1092–1105. [Google Scholar] [CrossRef]
- Krieger, D.; McNeil, M.; Zhang, J.; Puccio, A.; Schneider, W.; Li, X.; Okonwko, D.O. Very high resolution neuroelectric brain imag-ing by referee consensus processing. Int. J. Adv. Comput. Sci. 2014, 1, 15–25. [Google Scholar]
- Krieger, D.; Shepard, P.; Okonkwo, D.O. Normative atlases of neuroelectric brain activity and connectivity from a large human cohort. arXiv 2018, arXiv:1805.01552. [Google Scholar]
- Fischl, B.; Sereno, M.I.; Dale, A.M. Cortical Surface-Based Analysis. NeuroImage 1999, 9, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, M.; Schmansky, N.J.; Rosas, H.D.; Fischl, B. Within-subject template estimation for unbiased longitudinal image analysis. NeuroImage 2012, 61, 1402–1418. [Google Scholar] [CrossRef] [Green Version]
- Morin, C.M.; Belleville, G.; Bélanger, L.; Ivers, H. The Insomnia Severity Index: Psychometric Indicators to Detect Insomnia Cases and Evaluate Treatment Response. Sleep 2011, 34, 601–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, C.; Bélanger, L.; Ivers, H.; Morin, C.M. Validation of the Insomnia Severity Index in Primary Care. J. Am. Board Fam. Med. 2013, 26, 701–710. [Google Scholar] [CrossRef]
- Lancaster, M.A.; McCrea, M.A.; Nelson, L.D. Psychometric properties and normative data for the Brief Symptom Inventory-18 (BSI-18) in high school and collegiate athletes. Clin. Neuropsychol. 2016, 30, 321–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, S.S.; Rosenfeld, B.; Rasmussen, A. Measurement Invariance of the Brief Symptom Inventory in Survivors of Torture and Trauma. J. Interpers. Violence 2017, 32, 1708–1729. [Google Scholar] [CrossRef] [PubMed]
- Dixon, W.J. BMDP Statistical Software Manual; Chapter on BMDP7M; University of California Press: Berkeley, CA, USA, 1990. [Google Scholar]
- Afifi, A.; Clark, V. Computer Aided Multivariate Analysis; Wadsworth Publishing: London, UK, 1984. [Google Scholar]
- Tabachnick, B.G.; Fidell, L.S. Using Multivariate Statistics; Pearson/Allyn & Boeon: Boston, MA, USA, 2007. [Google Scholar]
- Taylor, J.R.; Williams, N.; Cusack, R.; Auer, T.; Shafto, M.A.; Dixon, M.; Tyler, L.K.; Henson, R.N. The Cambridge Centre for Ageing and Neuroscience (CamCAN) data repository: Structural and functional MRI, MEG, and cognitive data from a corss-sectional adult lifespan sample. Neuroimage 2015, 144, 262–269. [Google Scholar] [CrossRef]
- Shafto, M.A.; Can, C.-; Tyler, L.K.; Dixon, M.; Taylor, J.R.; Rowe, J.B.; Cusack, R.; Calder, A.J.; Marslen-Wilson, W.D.; Duncan, J.; et al. The Cambridge Centre for Ageing and Neuroscience (Cam-CAN) study protocol: A cross-sectional, lifespan, multidisciplinary examination of healthy cognitive ageing. BMC Neurol. 2014, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.W.; Kontos, A.P.; Okonkwo, D.O.; Almquist, J.; Bailes, J.; Barisa, M.; Bazarian, J.; Bloom, O.J.; Brody, D.; Cantu, R.; et al. Concussion is treatable: Statements of agreement from the targeted Evalua-tion and Active Management (TEAM) approaches to treating concussion meeting held in Pittsburgh, Ocober 15–16, 2015. Neurosurgery 2016, 79, 912–929. [Google Scholar] [CrossRef] [Green Version]
- Yendiki, A.; Panneck, P.; Srinivasan, P.; Stevens, A.; Zöllei, L.; Augustinack, J.; Wang, R.; Salat, D.; Ehrlich, S.; Behrens, T.; et al. Automated probabilistic reconstruction of white-matter pathways in health and disease using an atlas of the underlying anatomy. Front. Neuroinform. 2011, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Scahill, R.I.; Frost, C.; Jenkins, R.; Whitwell, J.L.; Rossor, M.N.; Fox, N.C. A Longitudinal Study of Brain Volume Changes in Normal Aging Using Serial Registered Magnetic Resonance Imaging. Arch. Neurol. 2003, 60, 989–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hämäläinen, M.; Hari, R.; Ilmoniemi, R.J.; Knuutila, J.; Lounasmaa, O.V. Magnetoencephalography—theory, instrumentation, and applications to noninvasive studies of the working human brain. Rev. Mod. Phys. 1993, 65, 413–497. [Google Scholar] [CrossRef] [Green Version]
- Krieger, D.; Onodipe, S.; Charles, P.J.; Sclabassi, R.J. Real time signal processing in the clinical setting. Ann. Biomed. Eng. 1998, 26, 462–472. [Google Scholar] [CrossRef] [Green Version]
- MaxFilter User’s Guide; Software Version 2.1; Elekta Neuromag Oy: Helsinki, Finland, 2008.
- Krieger, D.; Shepard, P.; Zusman, B.; Jana, A.; Okonkwo, D. Shared high value research resources: The CamCAN human lifespan neuroimaging dataset processed on the open science grid. In Proceedings of the 2017 IEEE International Conference on Bioinformatics and Biomedicine, Kansas City, MO, USA, 13–16 November 2017; IEEE: Piscatawy, NJ, USA, 2017; pp. 1815–1822. [Google Scholar]
- Pordes, R.; Petravick, D.; Kramer, B.; Olson, D.; Livny, M.; Roy, A.; Avery, P.; Blackburn, K.; Wenaus, T.; Würthwein, F.; et al. The open science grid. J. Phys. 2008, 78. [Google Scholar] [CrossRef]
- Sfiligoi, I.; Bradley, D.C.; Holzman, B.; Mhashilkar, P.; Padhi, S.; Wurthwein, F. The Pilot Way to Grid Resources Using glideinWMS. In Proceedings of the 2009 WRI World Congress on Computer Science and Information Engineering, Los Angeles, CA, USA, 31 March–2 April 2009; Volume 2, pp. 428–432. [Google Scholar] [CrossRef] [Green Version]
- Gan, Z.S.; Stein, S.C.; Swanson, R.; Guan, S.; Garcia, L.; Mehta, D.; Smith, D.H. Blood biomarkers for traumatic brain injury: A quanti-tative assessment of diagnostic and prognostic accuracy. Front. Neurol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Peltz, C.B.; Kenney, K.; Gill, J.; Diaz-Arrastia, R.; Gardner, R.C.; Yaffe, K. Blood biomarkers of traumatic brain injury and cognitive impairment in older veterans. Neurology 2020, 95, e1126–e1133. [Google Scholar] [CrossRef] [PubMed]
- Lewine, J.D.; Davis, J.T.; Sloan, J.H.; Kodituwakku, P.W.; Orrison, W.W., Jr. Neuromagnetic assessment of pathophysiologic brain ac-tivity induced by minor head trauma. Am. J. Neuroradiol. 1999, 20, 857–866. [Google Scholar] [PubMed]
- Huang, M.; Theilmann, R.J.; Robb, A.; Angeles, A.; Nichols, S.; Drake, A.; Dandrea, J.; Levy, M.; Holland, M.; Song, T.; et al. Integrated imaging approach with MEG and DTI to Detect Mild Traumatic Brain Injury in Military and Civilian Patients. J. Neurotrauma 2009, 1213–1226. [Google Scholar] [CrossRef]
- Huang, M.; Risling, M.; Baker, D.G. The role of biomarkers and MEG-based imaging markers in the diagnosis of post-traumatic stress disorder and blast-induced mild traumatic brain injury. Psychoneuroendocrinology 2016, 63, 398–409. [Google Scholar] [CrossRef] [Green Version]
- Sack, A.T.; Kadosh, R.C.; Schuhmann, T.; Moerel, M.; Walsh, V.; Goebel, R. Optimizing Functional Accuracy of TMS in Cognitive Studies: A Comparison of Methods. J. Cogn. Neurosci. 2009, 21, 207–221. [Google Scholar] [CrossRef]
- Alexander, B.; Laycock, R.; Crewther, D.P.; Crewther, S.G. An fMRI-Neuronavigated chronometric TMS investigation of V5 and intraparietal cortex in motion driven attention. Front. Hum. Neurosci. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Inuggi, A.; Filippi, M.; Chieffo, R.; Agosta, F.; Rocca, M.A.; González-Rosa, J.J.; Cursi, M.; Comi, G.; Leocani, L. Motor area localization using fMRI-constrained cortical current density reconstruction of movement-related cortical potentials, a comparison with fMRI and TMS mapping. Brain Res. 2010, 1308, 68–78. [Google Scholar] [CrossRef]
- Trapp, N.T.; Uitermarkt, B.; Johnson, M.K.; Koscik, T.R.; Garrett, L.; Heinzerling, A.; Zanaty, M.; Holland, M.T.; Howard, M.; Boes, A.D. A new device to improve target localization for transcranial magnetic stimulation therapy. Brain Stimul. 2019, 12, 1600–1602. [Google Scholar] [CrossRef]
- Luber, B.M.; Davis, S.; Bernhardt, E.; Neacsiu, A.; Kwapil, L.; Lisanby, S.H.; Strauman, T.J. Using neuroimaging to individualize TMS treatment for depression: Toward a new paradigm for imaging-guided intervention. Neuroimage 2017, 148, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Van Dun, K.; Bodranghien, F.; Manto, M.; Mariën, P. Targeting the Cerebellum by noninvasive neurostimulation: A review. Cerebellum 2017, 16, 695–741. [Google Scholar] [CrossRef] [PubMed]
- Sonmez, A.I.; Camsari, D.D.; Nandakumar, A.L.; Voort, J.L.V.; Kung, S.; Lewis, C.P.; Croarkin, P.E. Accelerated TMS for Depression: A systematic review and meta-analysis. Psychiatry Res. 2019, 273, 770–781. [Google Scholar] [CrossRef]
- Alexopoulos, G.S. Mechanisms and treatment of late-life depression. Transl. Psychiatry 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayhew, S.D.; Hylands-White, N.; Porcaro, C.; Derbyshire, S.W.; Bagshaw, A.P. Intrinsic variability in the human response to pain is assembled from multiple, dynamic brain processes. NeuroImage 2013, 75, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.-R.; Gross, J.; Uhlhaas, P. MEG—measured auditory steady-state oscillations show high test–retest reliability: A sensor and source-space analysis. NeuroImage 2015, 122, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legget, K.T.; Hild, A.K.; Steinmetz, S.E.; Simon, S.T.; Rojas, D.C. MEG and EEG demonstrate similar test-retest reliability of the 40 Hz auditory steady-state response. Int. J. Psychophysiol. 2017, 114, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Onishi, H.; Otsuru, N.; Kojima, S.; Miyaguchi, S.; Saito, K.; Inukai, Y.; Yamashiro, K.; Sato, D.; Tamaki, H.; Shirozu, H.; et al. Vaiability and reliability of paired-pulse depression and cortical oscillation induced by median nerve stimulation. Brain To-pogr. 2018, 31, 780–794. [Google Scholar] [CrossRef] [Green Version]
- Recasens, M.; Uhlhaas, M. Test-retest reliability of the magnetic mismatch negativity response to sound duration and omission deviants. NeuroImage 2017, 157, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Dunkley, B.T.; Wong, S.M.; Jetly, R.; Wong, J.K.; Taylor, M.J. Post-traumatic stress disorder and chronic hyperconnectivity in emo-tional processing. NeuroImage Clin. 2018, 20, 197–204. [Google Scholar] [CrossRef]
- Takeuchi, N.; Sugiyama, S.; Inui, K.; Kanemoto, K.; Nishihara, M. Long-latency suppression of auditory and somatosensory change-related cortical responses. PLoS ONE 2018, 13, e0199614. [Google Scholar] [CrossRef]
- Ahonen, L.; Huotilainen, M.; Brattico, E. Within- and between-session replicability of cognitive brain processes: An MEG study with an N-back task. Physiol. Behav. 2016, 158, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Solomon, J.; Boe, S.G.; Bardouille, T. Reliability for non-invasive somatosensory cortex localization: Implications for pre-surgical mapping. Clin. Neurol. Neurosurg. 2015, 139, 224–229. [Google Scholar] [CrossRef]
- Colclough, G.L.; Woolrich, M.W.; Tewarie, P.K.; Brookes, M.J.; Quinn, A.J.; Smith, S.M. How reliable are MEG resting-state connectivi-ty metrics? NeuroImage 2016, 138, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, S.I.; Routley, B.; Linden, D.E.; Singh, K.D. Reliability of Static and Dynamic Network Metrics in the Resting-State: A MEG-Beamformed Connectivity Analysis. Front. Neurosci. 2018, 12, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Buro, M.C.; Garcés, P.; Maestú, F. Test-retest reliability of resting-state magnetoencephalography power in sensor and source space. Hum. Brain Mapp. 2016, 37, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Candelaria-Cook, F.T.; Schendel, M.E.; Ojeda, C.J.; Bustillo, J.R.; Stephen, J.M. Reduced parietal alpha power and psychotic symp-toms: Test-retest reliability of resting-state mangetoencephalography in schizophrenica and health controls. Schizophr. Res. 2019, 215, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Marquetand, J.; Vannoni, S.; Carboni, M.; Hegner, Y.L.; Stier, C.; Braun, C.; Focke, N.K. Reliability of Magnetoencephalography and High-Density Electroencephalography Resting-State Functional Connectivity Metrics. Brain Connect. 2019, 9, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Piitulainen, H.; Illman, M.; Laaksonen, K.; Jousmäki, V.; Forss, N. Reproducibility of corticokinematic coherence. NeuroImage 2018, 179, 596–603. [Google Scholar] [CrossRef]
- Garcés, P.; Martin-Buro, M.C.; Maestú, F. Quantifying the test-retest reliability of magnetoencephalography resting-state func-tional connectivity. Brain Connect. 2016, 6, 448–460. [Google Scholar] [CrossRef]
- Kimura, T.; Ozaki, I.; Hashimoto, I. Impulse Propagation along Thalamocortical Fibers Can Be Detected Magnetically outside the Human Brain. J. Neurosci. 2008, 28, 12535–12538. [Google Scholar] [CrossRef] [Green Version]
- Roth, B.; Wikswo, J. The magnetic field of a single axon. A comparison of theory and experiment. Biophys. J. 1985, 48, 93–109. [Google Scholar] [CrossRef] [Green Version]
- Krieger, D.; McNeil, M.; Zhang, J.; Schneider, W.; Li, X.; Okonkwo, D.O. Referee consensus. In Proceedings of the Conference on Extreme Science and Engineering Discovery Environment: Gateway to Discovery, XSEDE 2013, San Diego, CA, USA, July 2007; ACM: New York, NY, USA, 2007. ISBN 978-1-4503-2170-9. [Google Scholar] [CrossRef]
- Krieger, D.; Becker, J.; Fabrizio, M.; McNeil, M.; Schneider, W.; Habeych, M.; Okonkwo, D. LP35: Task relevant high frequency neuroelectric oscillations in humans. Clin. Neurophysiol. 2014, 125 (Suppl. S1). [Google Scholar] [CrossRef]
- Antonakakis, M.; Dimitriadis, S.I.; Zervakis, M.; Papanicolaou, A.C.; Zouridakis, G. Aberrant Whole-Brain Transitions and Dynamics of Spontaneous Network Microstates in Mild Traumatic Brain Injury. Front. Comput. Neurosci. 2020, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, D.; McNeil, M.; Zhang, J.; Li, X. Referee consensus: A platform technology for nonlinear optimization. In Proceedings of the Conference on Extreme Science and Engineering Discovery Environment; Association for Computing Machinery: New York, NY, USA, 2013; Article No. 12; ISBN 9781450321709. [Google Scholar]











| Classified Negative | Classified Positive | Percentage | p-Value | |
|---|---|---|---|---|
| insomnia | ||||
| clinically negative | 42 | 12 | 77.8% | 0.000007 |
| clinically positive | 11 | 37 | 77.1% | 0.000031 |
| insomnia—2nd step | ||||
| clinically negative | 43 | 11 | 79.6% | 0.0000017 |
| clinically positive | 13 | 35 | 72.9% | 0.00036 |
| somatization | ||||
| clinically negative | 46 | 12 | 79.3% | 0.000001 |
| clinically positive | 14 | 30 | 68.2% | 0.0048 |
| depression | ||||
| clinically negative | 57 | 13 | 81.4% | 0.00000001 |
| clinically positive | 9 | 23 | 71.9% | 0.0035 |
| anxiety | ||||
| clinically negative | 37 | 18 | 67.3% | 0.0032 |
| clinically positive | 12 | 35 | 74.5% | 0.00017 |
| Insomnia | Somatization | Depression | Anxiety | |
|---|---|---|---|---|
| Brain-Stem | + | |||
| L Amygdala | + | |||
| L Hippocampus | -- | - | ||
| L Thalamus | -- | |||
| R Hippocampus | + | |||
| R Thalamus | ++ | |||
| L caudal middle frontal | - | |||
| L frontal pole | - | |||
| L inferior parietal | -- | |||
| L insula | -- | |||
| L lateral occipital | - | |||
| L lateral orbito-frontal | + | + | ||
| L paracentral | - | |||
| L pars opercularis | + | |||
| L pars triangularis | + | |||
| L precentral | -- | |||
| L rostral anterior cingulate | ++ | |||
| L rostral middle frontal | + | |||
| L superior frontal | - | |||
| L superior parietal | - | |||
| L temporal pole | + | |||
| R bankssts | -- | |||
| R caudal anterior cingulate | + | - | - | |
| R caudal middle frontal | -- | |||
| R cuneus | - | - | ||
| R inferior parietal | - | |||
| R insula | ++ | |||
| R isthmus cingulate | - | |||
| R lateral occipital | - | -- | ||
| R lateral orbito-frontal | - | |||
| R medial orbito-frontal | + | |||
| R middle temporal | - | |||
| R pericalcarine | + | |||
| R posterior central | + | + | ||
| R posterior cingulate | - | |||
| R supramarginal | + | + |
| t | df (Adjusted) | p-Value | Levene | p-Value | |
|---|---|---|---|---|---|
| insomnia | 9.20 | 87.8 | 10−14 | 0.10 | 0.756 |
| insomnia—2nd step | 6.70 | 86.8 | 10−9 | 0.39 | 0.532 |
| somatization | 8.63 | 73.6 | 10−13 | 1.27 | 0.262 |
| depression | 10.15 | 56.6 | 10−16 | 0.09 | 0.763 |
| anxiety | 2.14 | 90.6 | 0.025 | 0.00 | 0.980 |
| Symptom Coincidence Rates | Classifier Score Correlations | |||||
|---|---|---|---|---|---|---|
| Somatization | Depression | Anxiety | Somatization | Depression | Anxiety | |
| 0.67 | 0.61 | 0.71 | insomnia | 0.269 | 0.258 | 0.383 |
| 0.66 | 0.70 | somatization | 0.599 | 0.666 | ||
| 0.66 | depression | 0.781 | ||||
| CamCAN | TEAM-TBI | Percentage | p-Value | |
|---|---|---|---|---|
| Cortical/Subcortical | ||||
| CamCAN baseline | 581 | 38 | 93.9% | 10−126 |
| CamCAN follow-up | 227 | 25 | 90.1% | 10−42 |
| TEAM-TBI baseline | 0 | 63 | 100.0% | 10−18 |
| TEAM-TBI follow-up | 0 | 40 | 100.0% | 10−12 |
| cortical/subcortical—2nd step | ||||
| CamCAN baseline | 504 | 115 | 81.4% | 10−126 |
| CamCAN follow-up | 228 | 24 | 90.5% | 10−42 |
| TEAM-TBI baseline | 2 | 61 | 96.8% | 10−18 |
| TEAM-TBI follow-up | 3 | 37 | 92.5% | 10−12 |
| deep white matter | ||||
| CamCAN baseline | 518 | 71 | 87.9% | 10−85 |
| CamCAN follow-up | 208 | 31 | 87.0% | 10−33 |
| TEAM-TBI baseline | 14 | 48 | 77.4% | 0.0000024 |
| TEAM-TBI follow-up | 9 | 30 | 76.9% | 0.00015 |
| Brain-Stem | ++ | |
|---|---|---|
| Left | Right | |
| Accumbens | - | |
| Cerebellum | + | ++ |
| Caudate | -- | |
| Hippocampus | -- | |
| Pallidum | + | |
| Thalamus | -- | -- |
| fusiform | ++ | + |
| inferior parietal | + | |
| insula | -- | |
| isthmus cingulate | ++ | |
| lateral occipital | - | -- |
| lingual | ++ | |
| medial orbito-frontal | - | |
| middle temporal | - | -- |
| para-hippocampal | -- | |
| pars opercularis | - | - |
| pars triangularis | + | |
| peri-calcarine | - | |
| postcentral | - | - |
| precuneus | + | |
| precentral | - | |
| superior parietal | - | |
| superior temporal | -- | |
| transverse temporal | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krieger, D.; Shepard, P.; Soose, R.; Puccio, A.M.; Beers, S.; Schneider, W.; Kontos, A.P.; Collins, M.W.; Okonkwo, D.O. Symptom-Dependent Changes in MEG-Derived Neuroelectric Brain Activity in Traumatic Brain Injury Patients with Chronic Symptoms. Med. Sci. 2021, 9, 20. https://doi.org/10.3390/medsci9020020
Krieger D, Shepard P, Soose R, Puccio AM, Beers S, Schneider W, Kontos AP, Collins MW, Okonkwo DO. Symptom-Dependent Changes in MEG-Derived Neuroelectric Brain Activity in Traumatic Brain Injury Patients with Chronic Symptoms. Medical Sciences. 2021; 9(2):20. https://doi.org/10.3390/medsci9020020
Chicago/Turabian StyleKrieger, Don, Paul Shepard, Ryan Soose, Ava M. Puccio, Sue Beers, Walter Schneider, Anthony P. Kontos, Michael W. Collins, and David O. Okonkwo. 2021. "Symptom-Dependent Changes in MEG-Derived Neuroelectric Brain Activity in Traumatic Brain Injury Patients with Chronic Symptoms" Medical Sciences 9, no. 2: 20. https://doi.org/10.3390/medsci9020020
APA StyleKrieger, D., Shepard, P., Soose, R., Puccio, A. M., Beers, S., Schneider, W., Kontos, A. P., Collins, M. W., & Okonkwo, D. O. (2021). Symptom-Dependent Changes in MEG-Derived Neuroelectric Brain Activity in Traumatic Brain Injury Patients with Chronic Symptoms. Medical Sciences, 9(2), 20. https://doi.org/10.3390/medsci9020020