Effects of Low-Intensity Aerobic Exercise on Neurophysiological and Behavioral Correlates of Cognitive Function


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measures
2.2.1. General Medical History
2.2.2. Heart Rate (HR) and Intensity
2.2.3. Ratings of Perceived Exertion (RPE)
2.2.4. Oddball Paradigm
2.2.5. P3 Event-Related Potential (ERP)
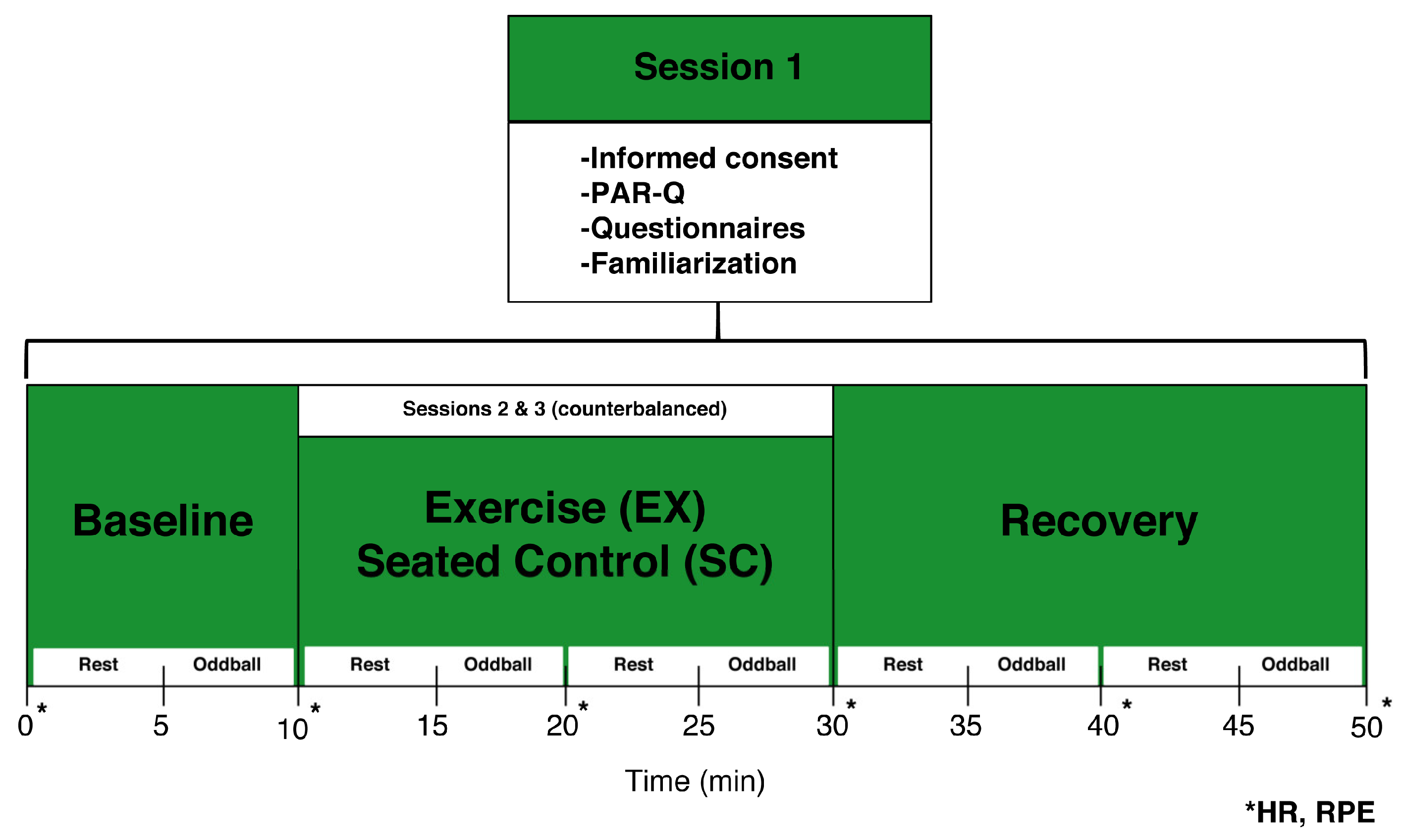
2.3. Procedures
2.4. Data Analysis
3. Results
3.1. Measures
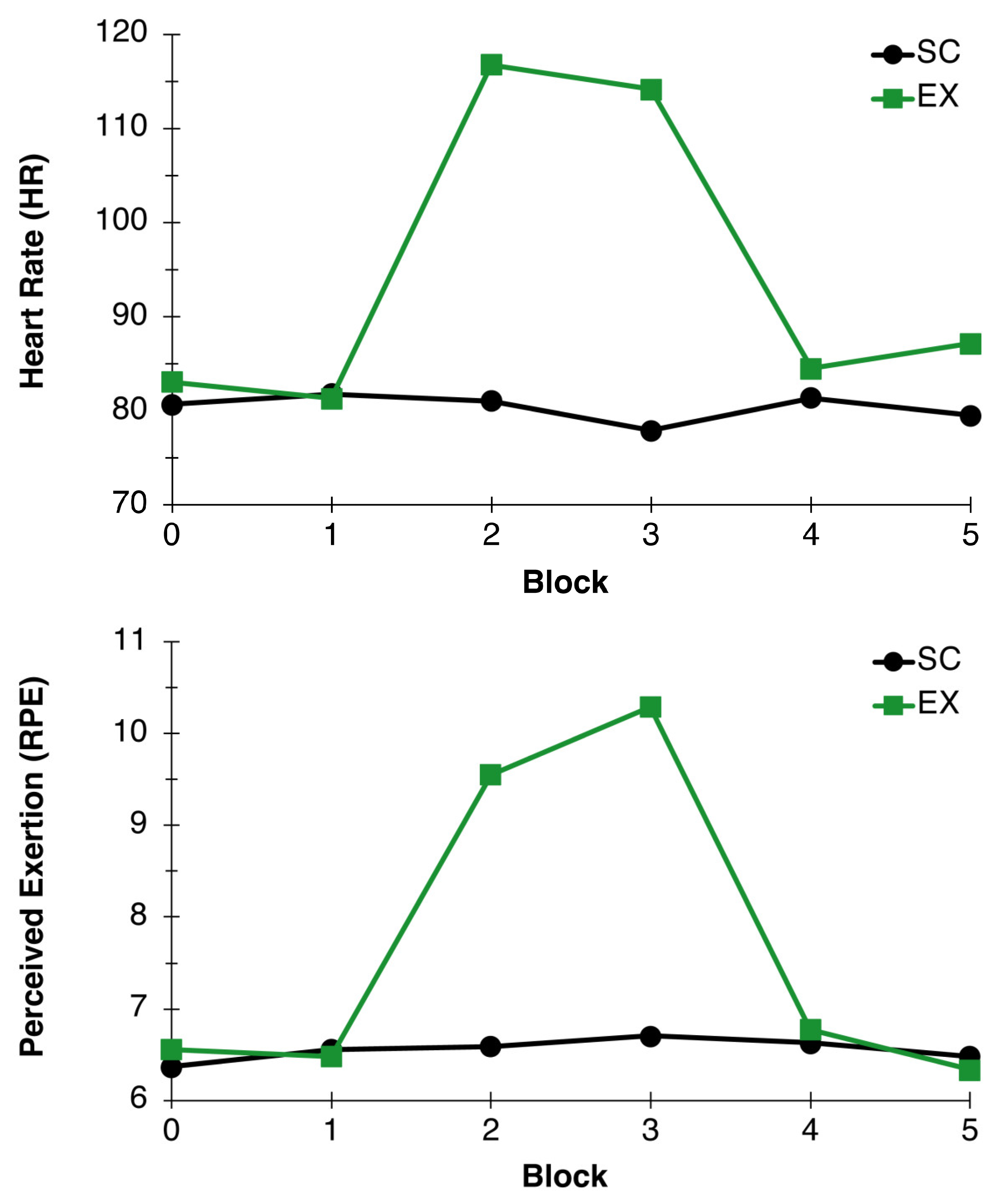
3.1.1. Heart Rate (HR)
3.1.2. Ratings of Perceived Exertion (RPE)
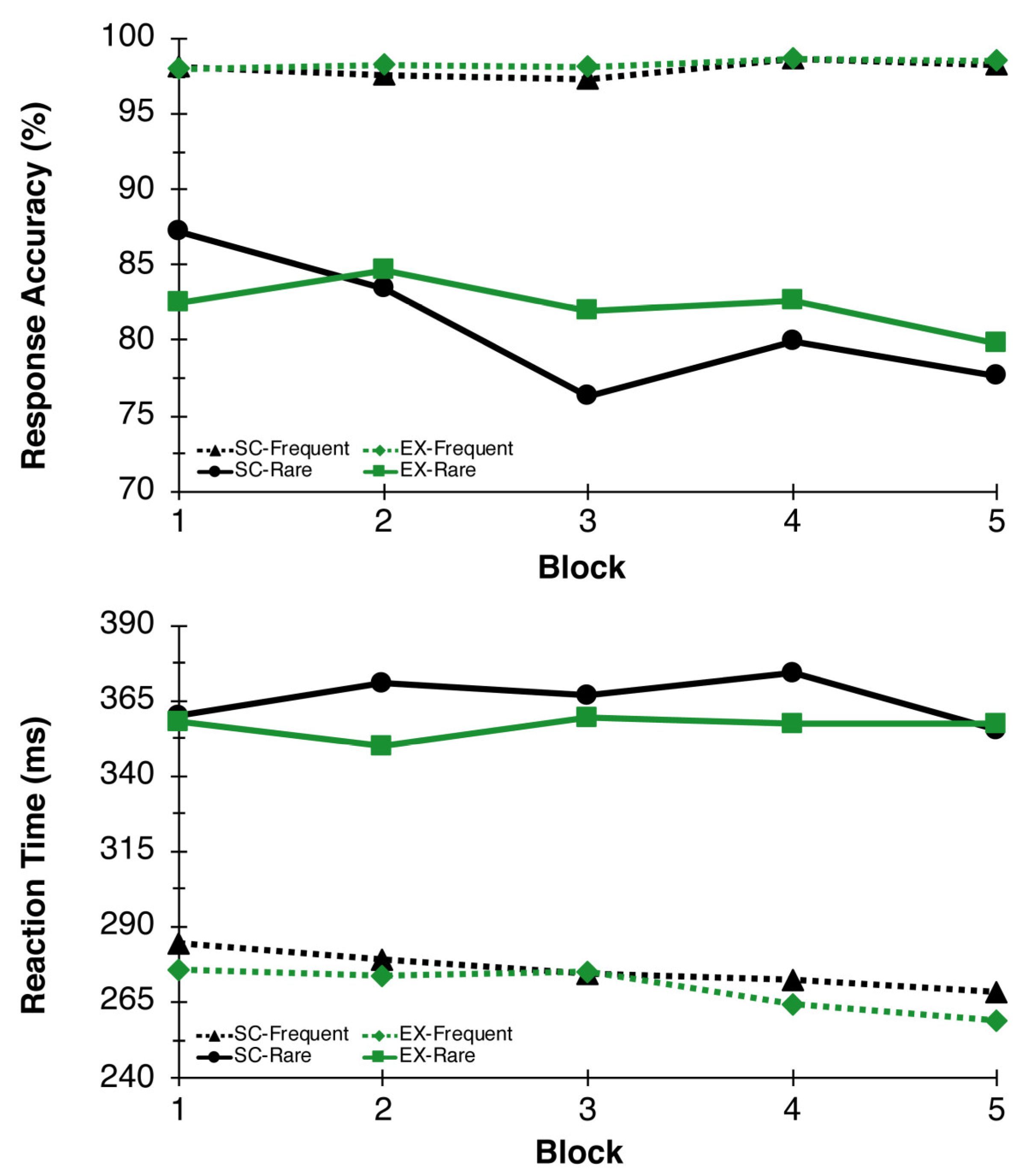
3.1.3. Response Accuracy and Reaction Time
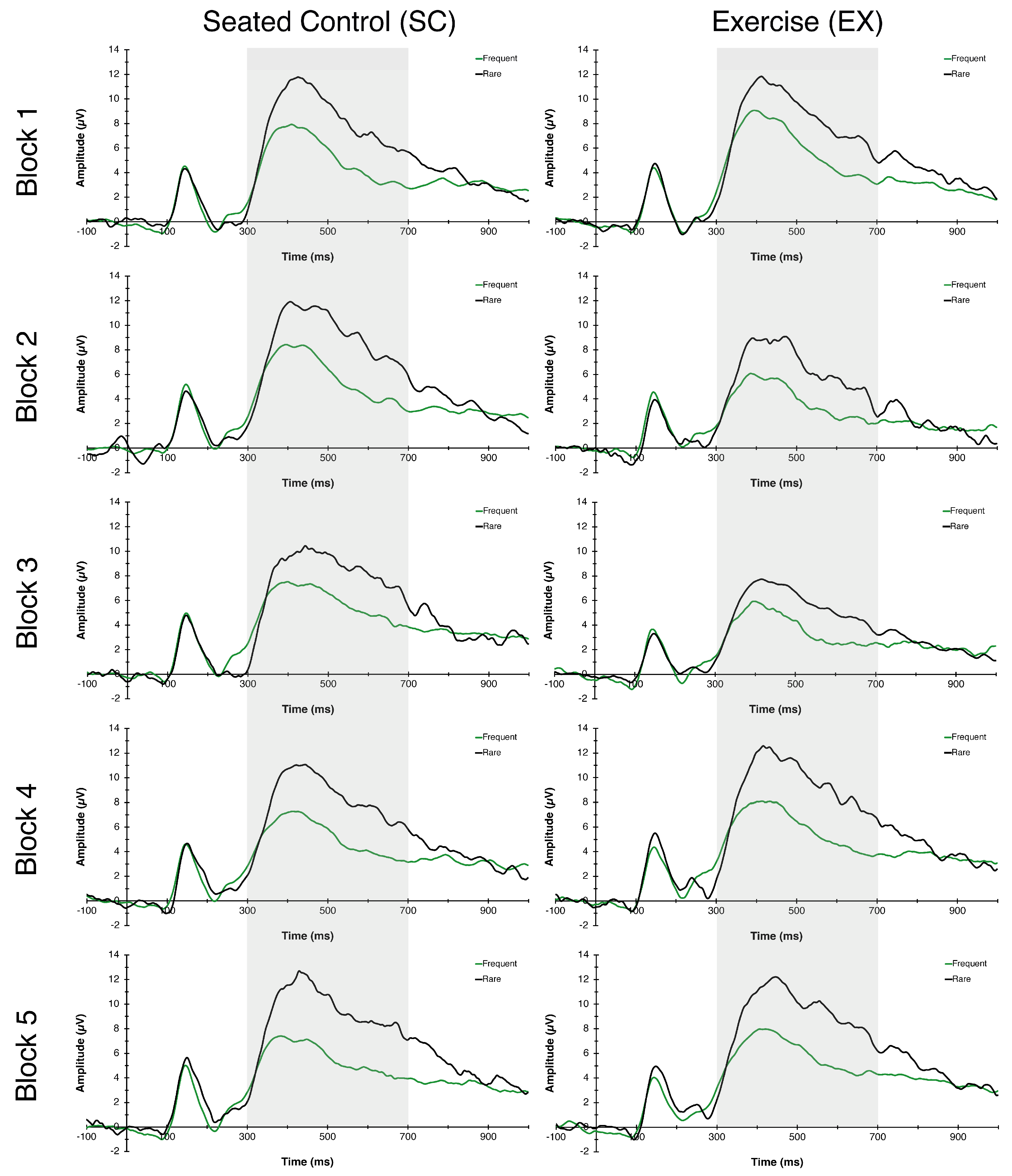
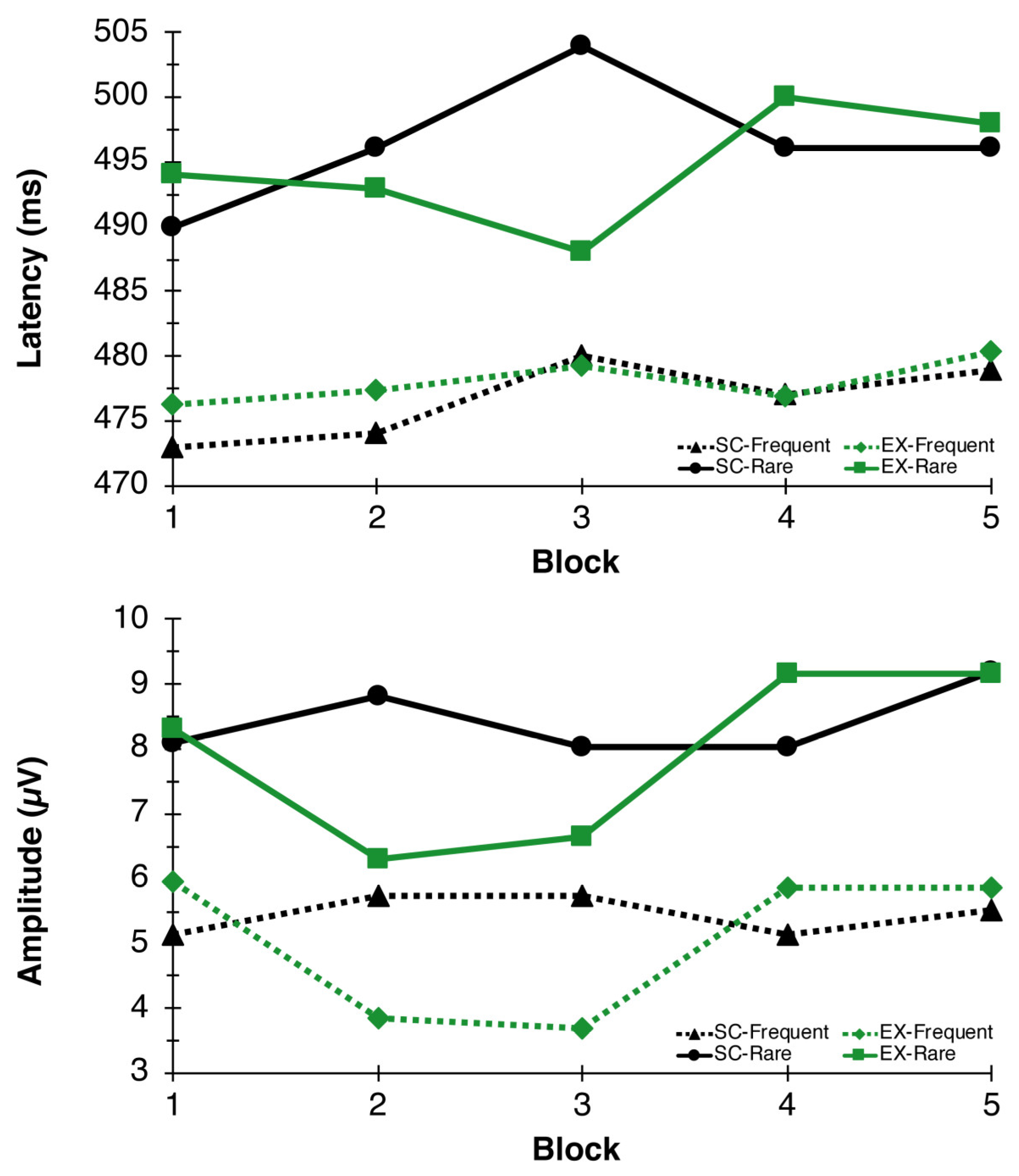
3.1.4. P3 Amplitude and Latency
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ishihara, T.; Drollette, E.S.; Ludyga, S.; Hillman, C.H.; Kamijo, K. The effects of acute aerobic exercise on executive function: A systematic review and meta-analysis of individual participant data. Neurosci. Biobehav. Rev. 2021, 128, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Lambourne, K.; Tomporowski, P. The effect of exercise-induced arousal on cognitive performance: A meta-regression analysis. Brain Res. 2010, 1341, 12–24. [Google Scholar] [CrossRef] [PubMed]
- McMorris, T.; Sproule, J.; Turner, A.; Hale, B.J. Acute, intermediate intensity exercise, and speed and accuracy in working memory tasks: A meta-analytical comparison of effects. Physiol. Behav. 2011, 102, 421–428. [Google Scholar] [CrossRef]
- Sibley, B.A.; Etnier, J.L. The relationship between physical activity and cognition in children: A meta-analysis. Pediatr. Exerc. Sci. 2003, 15, 243–256. [Google Scholar] [CrossRef]
- Oberste, M.; Javelle, F.; Sharma, S.; Joisten, N.; Walzik, D.; Bloch, W.; Zimmer, P. Effects and moderators of acute aerobic exercise on subsequent interference control: A systematic review and meta-analysis. Front. Psychol. 2019, 10, 2616. [Google Scholar] [CrossRef] [PubMed]
- Stimpson, N.J.; Davison, G.; Javadi, A.-H. Joggin’ the noggin: Towards a physiological understanding of exercise-induced cognitive benefits. Neurosci. Biobehav. Rev. 2018, 88, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Brisswalter, J.; Collardeau, M.; René, A. Effects of acute physical exercise characteristics on cognitive performance. Sports Med. 2002, 32, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.K.; Labban, J.D.; Gapin, J.I.; Etnier, J.L. The effects of acute exercise on cognitive performance: A meta-analysis. Brain Res. 2012, 1453, 87–101. [Google Scholar] [CrossRef]
- Etnier, J.L.; Salazar, W.; Landers, D.N.; Petruzzello, S.L.; Han, M.; Nowell, P. The influence of physical fitness and exercise upon cognitive functioning: A meta-analysis. J. Sport Exerc. Psychol. 1997, 19, 249–277. [Google Scholar] [CrossRef]
- Tomporowski, P.D. Effects of acute bouts of exercise on cognition. Acta Psychol. 2003, 112, 297–324. [Google Scholar] [CrossRef]
- Dodwell, G.; Müller, H.J.; Töllner, T. Electroencephalographic evidence for improved visual working memory performance during standing and exercise. Br. J. Psychol. 2019, 110, 400–427. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.L.; Chang, Y.K.; Brush, C.J.; Kwok, A.N.; Gordon, V.X.; Alderman, B.L. Neurophysiological and behavioral correlates of cognitive control during low and moderate intensity exercise. NeuroImage 2016, 131, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Sparling, P.B. Endurance exercise selectively impairs prefrontal-dependent cognition. Brain Cogn. 2004, 55, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Pontifex, M.B.; Hillman, C.H. Neuroelectric and behavioral indices of interference control during acute cycling. Clin. Neurophysiol. 2007, 118, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Scanlon, J.E.; Sieben, A.J.; Holyk, K.R.; Mathewson, K.E. Your brain on bikes: P3, MMN/N2b, and baseline noise while pedaling a stationary bike. Psychophysiology 2017, 54, 927–937. [Google Scholar] [CrossRef]
- Enders, H.; Cortese, F.; Maurer, C.; Baltich, J.; Protzner, A.B.; Nigg, B.M. Changes in cortical activity measured with EEG during a high-intensity cycling exercise. J. Neurophysiol. 2016, 115, 379–388. [Google Scholar] [CrossRef]
- Ludyga, S.; Gronwald, T.; Hottenrott, K. Effects of high vs. low cadence training on cyclists’ brain cortical activity during exercise. J. Sci. Med. Sport 2016, 19, 342–347. [Google Scholar] [CrossRef]
- Hillyard, S.A.; Anllo-Vento, L. Event-related brain potentials in the study of visual selective attention. Proc. Natl. Acad. Sci. USA 1998, 95, 781–787. [Google Scholar] [CrossRef]
- Luck, S.J.; Kappenman, E.S. (Eds.) The Oxford Handbook of Event-Related Potential Components; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Donchin, E.; Coles, M.G. Is the P300 component a manifestation of context updating? Behav. Brain Sci. 1988, 11, 357–374. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef]
- Shaw, E.P.; Rietschel, J.C.; Hendershot, B.D.; Pruziner, A.L.; Miller, M.W.; Hatfield, B.D.; Gentili, R.J. Measurement of attentional resources and mental effort for cognitive workload assessment under various task demands during dual-task walking. Biol. Psychol. 2018, 134, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Aly, M.; Ogasawara, T.; Kamijo, K.; Kojima, H. Neurophysiological Evidence of the Transient Beneficial Effects of a Brief Mindfulness Exercise on Cognitive Processing in Young Adults: An ERP Study. Mindfulness 2023, 1–11. [Google Scholar] [CrossRef]
- Dodwell, G.; Liesefeld, H.R.; Conci, M.; Müller, H.J.; Töllner, T. EEG evidence for enhanced attentional performance during moderate-intensity exercise. Psychophysiology 2021, 58, e13923. [Google Scholar] [CrossRef] [PubMed]
- Drollette, E.S.; Scudder, M.R.; Raine, L.B.; Moore, R.D.; Saliba, B.J.; Pontifex, M.B.; Hillman, C.H. Acute exercise facilitates brain function and cognition in children who need it most: An ERP study of individual differences in inhibitory control capacity. Dev. Cogn. Neurosci. 2014, 7, 53–64. [Google Scholar] [CrossRef] [PubMed]
- De Vos, M.; Debener, S. Mobile EEG: Towards brain activity monitoring during natural action and cognition. Int. J. Psychophysiol. 2014, 91, 1–2. [Google Scholar] [CrossRef]
- Gramann, K.; Gwin, J.T.; Bigdely-Shamlo, N.; Ferris, D.P.; Makeig, S. Visual evoked responses during standing and walking. Front. Hum. Neurosci. 2010, 4, 202. [Google Scholar] [CrossRef]
- Schmidt-Kassow, M.; Deusser, M.; Thiel, C.; Otterbein, S.; Montag, C.; Reuter, M.; Kaiser, J. Physical exercise during encoding improves vocabulary learning in young female adults: A neuroendocrinological study. PLoS ONE 2013, 8, e64172. [Google Scholar] [CrossRef]
- Zink, R.; Hunyadi, B.; Van Huffel, S.; De Vos, M. Mobile EEG on the bike: Disentangling attentional and physical contributions to auditory attention tasks. J. Neural Eng. 2016, 13, 046017. [Google Scholar] [CrossRef]
- Yagi, Y.; Coburn, K.L.; Estes, K.M.; Arruda, J.E. Effects of aerobic exercise and gender on visual and auditory P300, reaction time, and accuracy. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 402–408. [Google Scholar] [CrossRef]
- Grego, F.; Vallier, J.M.; Collardeau, M.; Bermon, S.; Ferrari, P.; Candito, M.; Brisswalter, J. Effects of long duration exercise on cognitive function, blood glucose, and counterregulatory hormones in male cyclists. Neurosci. Lett. 2004, 364, 76–80. [Google Scholar] [CrossRef]
- Vogt, T.; Herpers, R.; Scherfgen, D.; Strüder, H.K.; Schneider, S. Neuroelectric adaptations to cognitive processing in virtual environments: An exercise-related approach. Exp. Brain Res. 2015, 233, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Polich, J. Neuropsychology of P300. In Oxford Handbook of Event-Related Potential Components; Oxford University Press: New York, NY, USA, 2012; pp. 159–188. [Google Scholar]
- Donchin, E. Surprise!… surprise? Psychophysiology 1981, 18, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.J.; Cox, M.H.; Simper, K. An analysis of “Par-Q” responses in an office population. Can. J. Public Health 1981, 72, 37–40. [Google Scholar]
- Beck, A.T.; Steer, R.A.; Brown, G.K. Manual for the Beck Depression Inventory-II; Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Spielberger, C.D.; Gorsuch, R.L.; Lushene, R.; Vagg, P.R.; Jacobs, G.A. Manual for the State-Trait Anxiety Inventory; Consulting Psychologists Press: Palo Alto, CA, USA, 1983. [Google Scholar]
- Cohen, S.; Kamarck, T.; Mermelstein, R. A global measure of perceived stress. J. Health Soc. Behav. 1983, 24, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Garber, C.E.; Blissmer, B.; Deschenes, M.R.; Franklin, B.A.; Lamonte, M.J.; Lee, I.M.; Swain, D.P. Quantity and quality of exercise for developing and maintaining cardiorespiratory, musculoskeletal, and neuromotor fitness in apparently healthy adults: Guidance for prescribing exercise. Med. Sci. Sport. Exerc. 2011, 43, 1334–1359. [Google Scholar] [CrossRef] [PubMed]
- Borg, G. Perceived exertion as an indicator of somatic stress. Scand. J. Rehabil. Med. 1970, 2, 92–98. [Google Scholar]
- Chen, M.J.; Fan, X.; Moe, S.T. Criterion-related validity of the Borg ratings of perceived exertion scale in healthy individuals: A meta-analysis. J. Sport. Sci. 2002, 20, 873–899. [Google Scholar] [CrossRef]
- Borg, G. Borg’s Perceived Exertion and Pain Scales; Human Kinetics: Champaign, IL, USA, 1998. [Google Scholar]
- Luck, S.J.; Kappenman, E.S.; Fuller, R.L.; Robinson, B.; Summerfelt, A.; Gold, J.M. Impaired response selection in schizophrenia: Evidence from the P3 wave and lateralized readiness potential. Psychophysiology 2009, 46, 776–786. [Google Scholar] [CrossRef]
- Oostenveld, R.; Praamstra, P. The five percent electrode system for high-resolution EEG and ERP measurements. Clin. Neurophysiol. 2001, 112, 713–719. [Google Scholar] [CrossRef]
- Dien, J. The ERP PCA Toolkit: An open source program for advanced statistical analysis of event-related potential data. J. Neurosci. Methods 2010, 187, 138–145. [Google Scholar] [CrossRef]
- Bertrand, O.; Perrin, F.; Pernier, J. A theoretical justification of the average reference in topographic evoked potential studies. Electroencephalogr. Clin. Neurophysiol. 1985, 62, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Tucker, D.M.; Liotti, M.; Potts, G.F.; Russsell, G.S.; Posner, M.I. Spatiotemporal analysis of brain electrical fields. Hum. Brain Mapp. 1994, 1, 134–152. [Google Scholar] [CrossRef]
- Ritter, W.; Simson, R.; Vaughan, H.G., Jr.; Friedman, D. A brain event related to the making of a sensory discrimination. Science 1979, 203, 1358–1361. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R., Jr. On the neural generators of the P300 component of the event-related potentials. Psychophysiology 1993, 30, 90–97. [Google Scholar] [CrossRef]
- Davranche, K.; Brisswalter, J.; Radel, R. Where are the limits of the effects of exercise intensity on cognitive control? J. Sport Health Sci. 2015, 4, 56–63. [Google Scholar] [CrossRef]
- Schmit, C.; Davranche, K.; Easthope, C.S.; Colson, S.S.; Brisswalter, J.; Radel, R. Pushing to the limits: The dynamics of cognitive control during exhausting exercise. Neuropsychologia 2015, 68, 71–81. [Google Scholar] [CrossRef]
- Audiffren, M.; Tomporowski, P.D.; Zagrodnik, J. Acute aerobic exercise and information processing: Modulation of executive control in a random number generation task. Acta Psychol. 2009, 132, 85–95. [Google Scholar] [CrossRef]
- Dietrich, A. The transient hypofrontality theory and its implications for emotion and cognition. In Exercise and Cognitive Function; John Wiley and Sons, Ltd.: Chichester, UK, 2009; pp. 69–90. [Google Scholar]
- Pesce, C. An integrated approach to the effect of acute and chronic exercise on cognition: The linked role of individual and task constraints. In Exercise and Cognitive Function; John Wiley and Sons, Ltd.: Chichester, UK, 2009; pp. 213–226. [Google Scholar]
- Broadbent, D.E. Perception and Communication; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Keele, S.W. Attention and Human Performance; Goodyear Publishing Company: Warwickshire, UK, 1973. [Google Scholar]
- Arnsten, A.F. Stress signaling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009, 10, 410–422. [Google Scholar] [CrossRef]
- Ramos, B.P.; Arnsten, A.F. Adrenergic pharmacology and cognition: Focus on the prefrontal cortex. Pharmacol. Ther. 2007, 113, 523–536. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | Male (n = 17) | Female (n = 10) | Total (N = 27) |
|---|---|---|---|
| Age (years) | 23.6 ± 3.4 | 21.6 ± 1.3 | 22.9 ± 3.0 |
| BMI (kg/m2) | 25.5 ± 3.3 | 25.2 ± 4.7 | 25.4 ± 3.8 |
| Depressive Symptoms (BDI) 1 | 3.9 ± 3.5 | 6.0 ± 5.1 | 4.7 ± 4.2 |
| Anxiety levels (STAI) | 47.3 ± 3.7 | 46.6 ± 2.3 | 47.0 ± 3.2 |
| Perceived Stress (PSS) | 27.7 ± 5.7 | 31.2 ± 4.0 | 29.0 ± 5.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olson, R.L.; Cleveland, D.J.; Materia, M. Effects of Low-Intensity Aerobic Exercise on Neurophysiological and Behavioral Correlates of Cognitive Function. Behav. Sci. 2023, 13, 401. https://doi.org/10.3390/bs13050401
Olson RL, Cleveland DJ, Materia M. Effects of Low-Intensity Aerobic Exercise on Neurophysiological and Behavioral Correlates of Cognitive Function. Behavioral Sciences. 2023; 13(5):401. https://doi.org/10.3390/bs13050401
Chicago/Turabian StyleOlson, Ryan L., David J. Cleveland, and Melissa Materia. 2023. "Effects of Low-Intensity Aerobic Exercise on Neurophysiological and Behavioral Correlates of Cognitive Function" Behavioral Sciences 13, no. 5: 401. https://doi.org/10.3390/bs13050401
APA StyleOlson, R. L., Cleveland, D. J., & Materia, M. (2023). Effects of Low-Intensity Aerobic Exercise on Neurophysiological and Behavioral Correlates of Cognitive Function. Behavioral Sciences, 13(5), 401. https://doi.org/10.3390/bs13050401