
A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
“In undisturbed rivers, each salmon population is composed of a bundle of several life histories, or several alternative survival strategies. Unlike the salmon raised in a hatchery environment with its feedlot regime, the salmon in a natural population in a healthy river do not all do the same thing in the same place at the same time. They follow different pathways through the time and space dimensions of their habitat. As the riverscape changes due to natural disturbances (fires, floods, droughts, and so on) some of the salmon’s life histories are survival peaks while others drop into troughs. This diverse array of life histories diminishes the risk of catastrophic mortality and loss of an entire population in a naturally changing environment.”
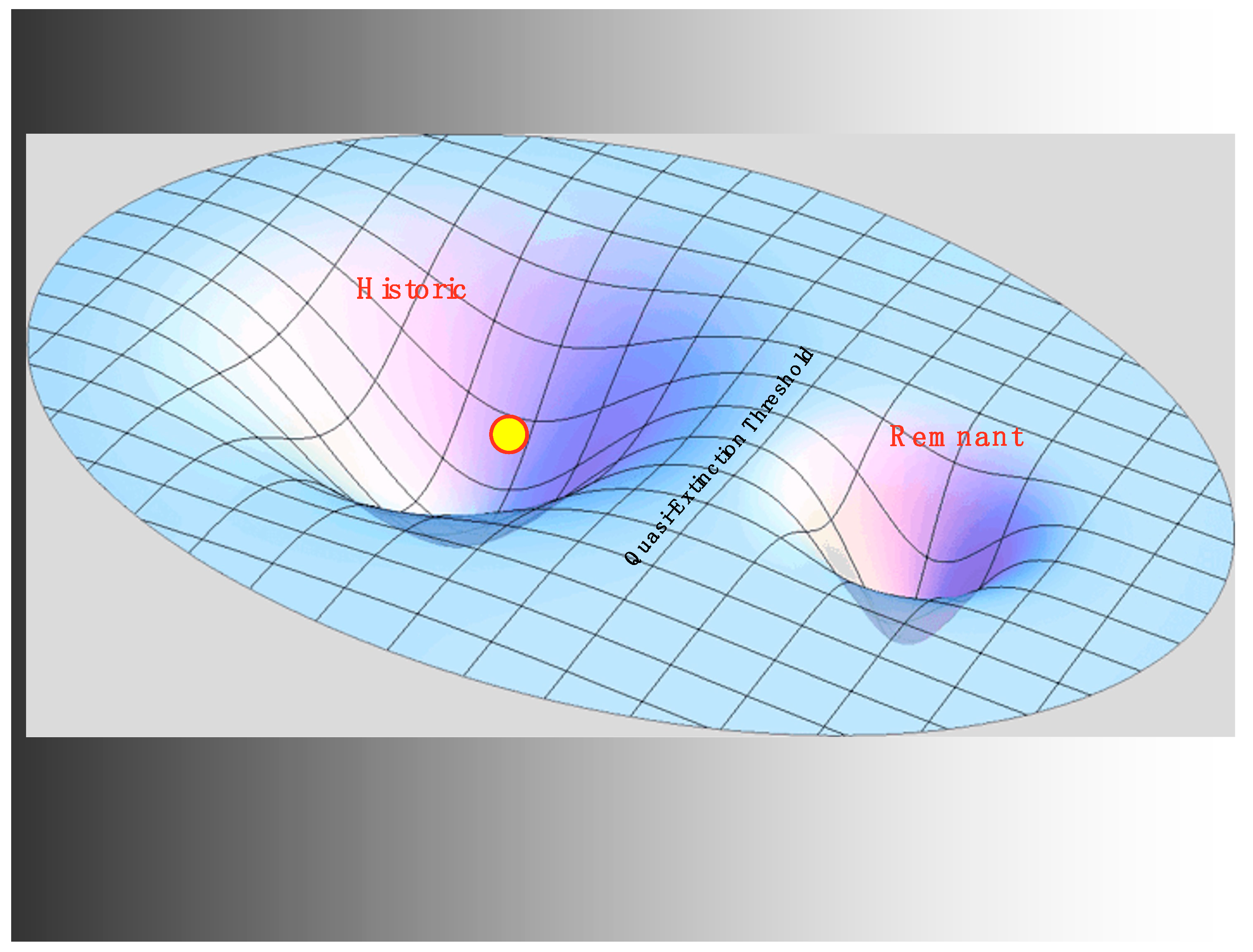
3. Regimes and Critical Transitions
3.1. The Historic Regime
“Life history forms, therefore, are genetically predisposed to strategies that optimize survival under the restraints of local stream environments. When referring to life history strategy, we are describing a behavioral repertoire that has proven successful for a particular population of the species under given environmental constraints. For example, fall Chinook that utilize a stream for incubation and early rearing that is unsuitable later in the summer or winter will acquire a short-term pattern of residence and disperse downstream before unsuitable conditions arise…”
“We approached the stratum risk criteria from the perspective of the historical template. The approach is based on the principle that the historical population structure of the strata produced a relatively low risk of extinction, and the closer the population structure is to that historical structure, the lower its extinction risk. Conversely a population structure that deviates greatly from the historical structure would be considered at high risk.”
“About 100,000 years ago, a large portion of the volcano’s north flank and summit collapsed. The resulting debris avalanche transformed into a lahar that swept down the Hood River valley. At the river’s mouth, where the town of Hood River now stands, the lahar was 400 feet deep. The lahar crossed the Columbia River and surged up the White Salmon River valley on the Washington side. Since that time lava has filled in the scar left by the debris avalanche”.[50]
“Indians were seasoned rational fishers. They developed a culture and economy that meshed well with nature. Respect, propitiation, utility, and territoriality reinforced a coherent, moderate strategy of exploitation. The synergistic result was a culturally specific form of conservation. No single mechanism entirely mitigated the potential for destruction, but in combination they worked wonders. Fishing techniques could wreak havoc on salmon runs, and cultural routes to prestige encouraged heavy consumption; but cultural forms of respect moderated harvest, and the technical limitations of food preservation capped storage capacity.”
“The recent ages of some of the geologic events that formed these features show quite emphatically that the pre-1805 landscape—including the land, people, and ecosystems—was not static but one of drastic and dramatic change… The large volcanic eruptions of Mount Hood in about 500 and again in the 1780s, as well as the large Mount St. Helens eruptions of about 1480, almost certainly had large effects on the Columbia River and the ecosystems and occupants that depended on it… Large sediment pulses into the river from volcanic eruptions are analogous to, but far larger than, sedimentation caused by land-use practices. The huge Missoula Floods of the last ice age reshaped and locally dredged the Columbia River channel…Yet, the tremendous natural resources sustaining the Native American populations at the time of Lewis and Clark attest to the resilience of the Columbia ecosystem in the face of such huge disturbances, even ones of just a few decades or centuries before.”
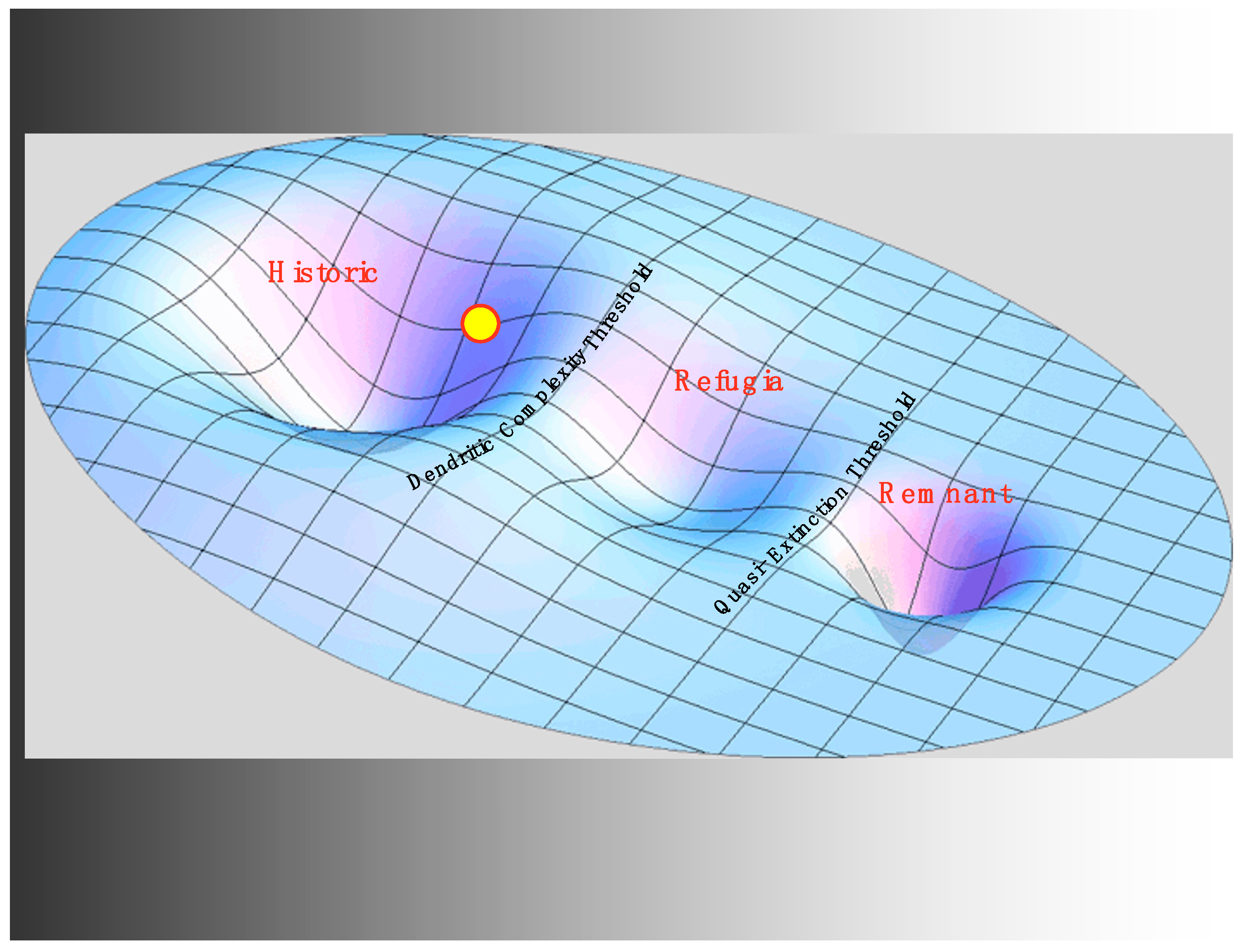
3.2. Habitat, Harvest, and a Preview of the Refugia Regime
“Severe scouring from splash damming was one of the earliest reported forms of widespread anthropogenic disturbance in streams of the Pacific Northwest, USA. Splash damming was a common method of log transport in western Oregon from the 1880s through the 1950s. Before being released in large freshets to downstream lumber mills, water and logs were stored in reservoirs behind splash dams. Further protocol called for dynamiting downstream obstacles such as large boulders and natural logjams… [resulting in] significantly more bedrock and fewer deep pools in splashed reaches… three times fewer pieces of key large wood were found in splashed reaches… Many of the in-channel variables that demonstrated significant differences are regarded as indicators of salmon habitat quality… splash-dam legacy effects still persist on evaluated stream reaches 50–130 years after the practice ceased…”[59]
“Concern for the fishing industry prompted sporadic and ineffectual conservation measures. As early as 1877, the legislature of Washington Territory declared a closed season—and the next year Oregon responded with similar measures. State governments also curtailed the use of certain types of fishing gear on the Columbia River. During the late 19th century Washington and Oregon prohibited fish traps, weirs, seines, and nets placed two-thirds of the way across freshwater streams, creeks, or lakes, if they prevented the passage of fish. Purse seines, which consisted of a long curtain of webbing drawn by a boat, were prohibited on the Columbia River in 1917 and in the coastal waters of both Washington and Oregon in 1922. Fishwheels, as noted, were banned in 1926 and 1934. However, many of these restraints were rarely enforced. Although Oregon created a Board of Fish Commissioners in 1887 and Washington established a Fish Commission in 1890, neither organization possessed sufficient funds to police the river and catch offenders. Hence, early restrictions on fishing did little to stop the decline of the salmon runs”.[69]
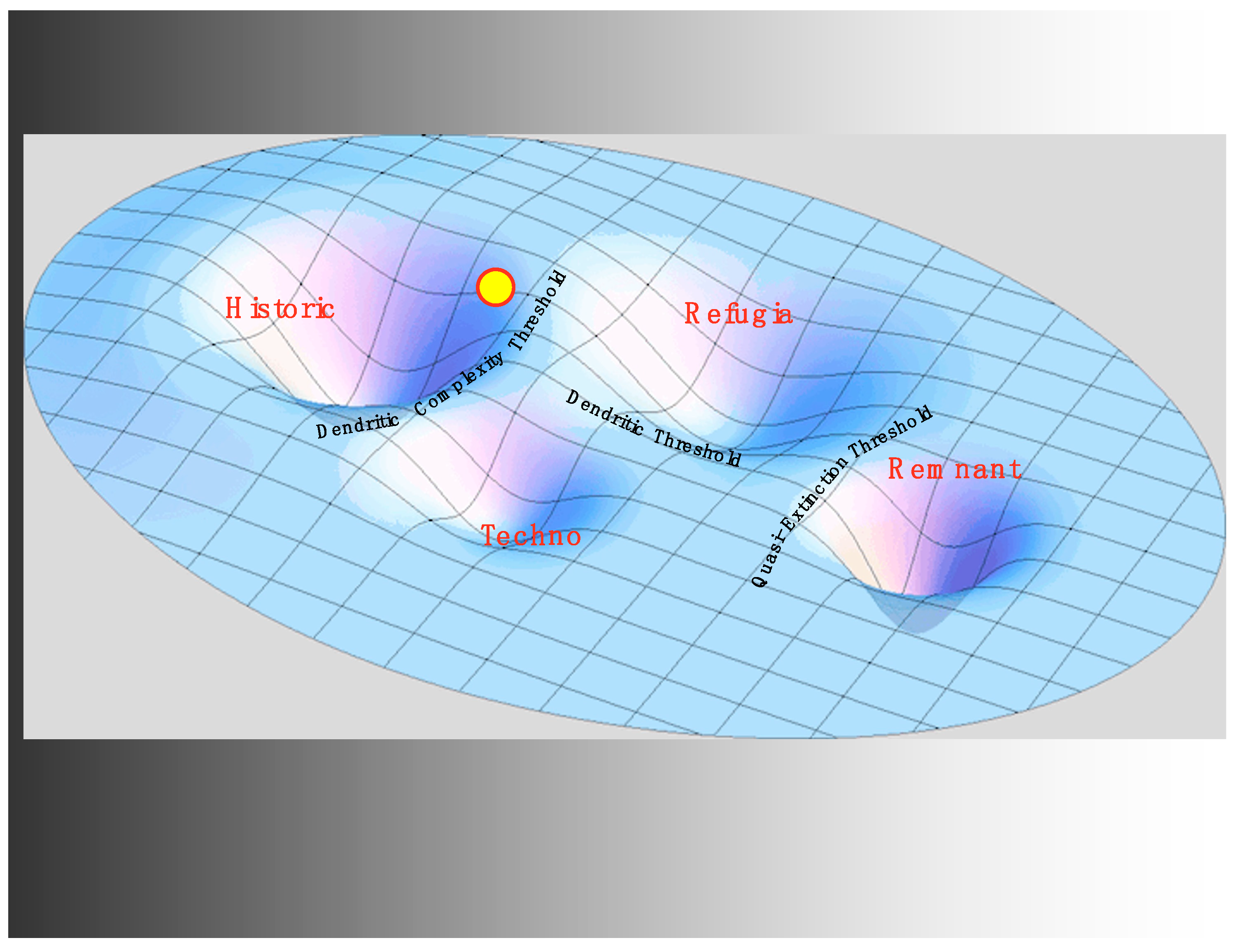
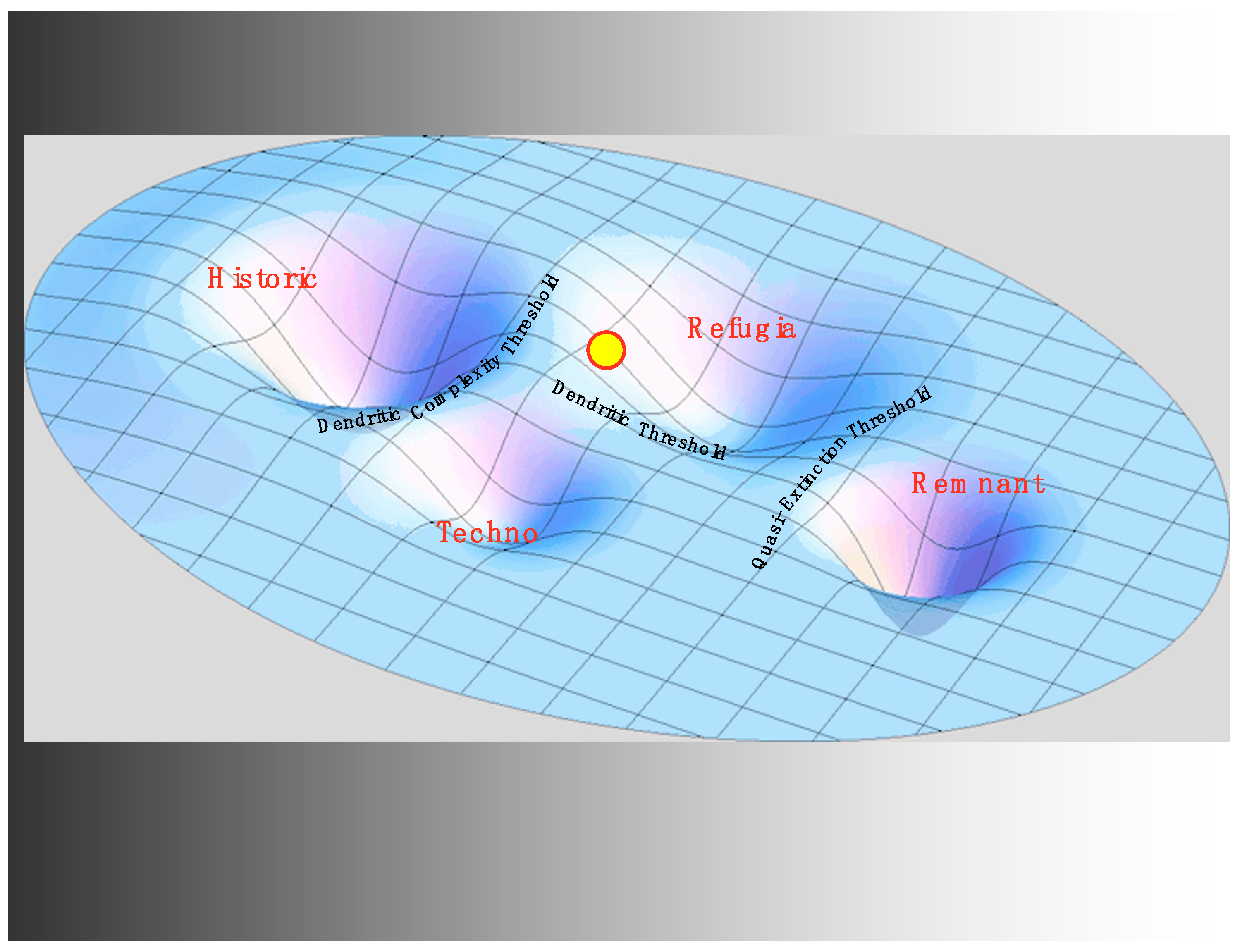
3.3. Hatcheries and the Techno Regime
“Hatcheries sought to wed technology and biology, to merge factorylike production with natural production. The canners, many fishermen, and many experts on the fisheries came to regard nature as inefficient. H. A. Jones, the federal official who first studied the decline of the Columbia fisheries, produced a rather typical piece of nineteenth-century arithmetic. He calculated that a $10,000 investment in a single hatchery and operating expenses of $3000 a year would produce 5 million mature fish a year, which, when canned, would yield $5 million. This was more than twice the fish and twice the income then being produced on the Columbia. “These figures border on the marvelous”, Jones admitted, but encapsulated in such marvelous numbers was a hope that an inefficient nature, with a helping managerial hand, could solve both a biological and a social crisis”.[71]
3.4. Hydro and the Critical Transition to the Refugia Regime
“For Roosevelt, the huge public works projects that would put people to work and raise America from the Depression were projects of opportunity…Puget Sound Power and Light Company completed Rock Island Dam in 1933… It was the first dam on the mainstem Columbia River, and it was not a multiple-purpose dam. Its sole purpose was to generate electricity. Construction began at Grand Coulee and Bonneville, both multiple-purpose dams, in 1933; Bonneville was completed in 1938 and Grand Coulee in 1941. The last major dams … were Lower Granite on the Snake River and Libby on the Kootenai, both in 1975. There are 14 dams on the mainstem Columbia from Mica to Bonneville, and five on the lower Snake from Hells Canyon Dam to Ice Harbor Dam… Collectively, Columbia River Basin dams have a total nameplate capacity of 36,682.2 megawatts and produce, on average, 16,604.42 megawatts of electricity…”
“Knowing further that each race [stock] is self-propagating, it becomes perfectly apparent that all parts of the salmon run in the Columbia River must be given adequate protection if the run as a whole is to be maintained. The protection of only one or two portions of the run will not be sufficient, inasmuch as certain races will be left entirely unprotected.”
“The transformation of the Columbia River began years before 1949… In the spring of 1933… President Franklin D. Roosevelt… directed federal monies at two huge public works projects’ Bonneville Dam near Portland and Grand Coulee Dam in north-central Washington. Roosevelt used public works to address fundamental economic issues by creating employment and stimulating regional development in three of the nation’s river basins, the Tennessee, the Missouri, and the Columbia. What the New Deal began on these rivers changed them dramatically and refashioned the regions they drained. Grand Coulee and Bonneville dams blockaded the Columbia, the second largest river by volume of flow in the United States and the river with the greatest hydroelectric potential on the face of the globe. Roosevelt’s action put the river on a new course, literally and figuratively.”
“Fish passage ends at Chief Joseph Dam at River Mile 545 on the Columbia. Before that dam was completed, fish passage ended at Grand Coulee Dam, 51 miles upriver. In the Snake River system, fish passage ends at Hells Canyon Dam, at River Mile 247, and, on the North Fork Clearwater River, at Dworshak Dam, which is about three miles from the confluence of the North Fork with the mainstem of the Clearwater”.[79]
“We combined existing inventories of barriers to adult fish passage in the Willamette and Lower Columbia River basins and identified 1491 anthropogenic barriers to fish passage blocking 14,931 km of streams. We quantified and compared the stream quality, land cover, and physical characteristics of lost versus currently accessible habitat by watershed, assessed the effect of barriers on the variability of accessible habitats, and investigated potential impacts of habitat reduction on endangered or threatened salmonid populations. The majority of the study watersheds have lost more than 40% of total fish stream habitat. Overall, 40% of the streams with spawning gradients suitable for steelhead (anadromous rainbow trout Oncorhynchus mykiss), 60% of streams with riparian habitat in good condition, and 30% of streams draining watersheds with all coniferous land cover are no longer accessible to anadromous fish… The differences between lost and accessible streams suggest that high-quality habitat has been disproportionately lost in many watersheds.”
“Interestingly, engineers viewed dams as mimicking nature since some of the largest glacial dams in the world existed in the same area during the Pleistocene epoch; in their view, they were returning it to a previous natural state… Nonetheless, increased damming harmed local wildlife, prevented fish from moving to spawning grounds, overstressed the landscape, and allowed for another and more harmful industry”.[91]
“[The Organic Machine’s] thesis is that the best way to understand the river is as an entity that has been in constant flux. Gradually human beings have modified it. They have created the illusion of conquering the river, of turning it, as the common phrase is in the Pacific Northwest, into a series of slackwater lakes. We apply social language to the river. We have raped it or killed it; but such language is deceptive. We have changed the Columbia to the detriment of some species and the benefit of others. Where once the Columbia said salmon, it now says shad and squawfish. The Columbia is not dead. The dams depend on larger natural rhythms of snowfall and snowmelt, of rain and gravity and seasons, but we have created a system where what is natural and what is human becomes harder and harder to distinguish. Each intrudes on and influences the other. The river has become an organic machine”.[96]
“Many of the region’s environments have been permanently altered in ways that do not favor salmon. The Columbia Basin, for example, is now dominated by a series of mainstem and tributary reservoirs. Land use in much of the watershed has also changed in ways that no longer favor salmon…As dramatic as the environmental changes are, some fishes, especially exotics, are thriving (e.g., walleye, American shad… smallmouth bass… northern pike… and brook trout…). These exotic species are well adapted to the new environment. It would be difficult—some argue impossible—to re-create the Pacific Northwest habitats that once existed and were ideal for wild salmon. A simpler, cheaper policy option would be to manage for those fishes, typically exotics, best suited to current habitat. Such an approach, while relatively easy and cheap to accomplish, would be an explicit decision to terminate many stocks of wild salmon.”
4. Discussion
- Restore anadromous fishes to the rivers and streams that support the historical, cultural, and economic practices of the tribes. (These are generally areas above Bonneville Dam.)
- Emphasize strategies that rely on natural production and healthy river systems to achieve this goal.
- Protect tribal sovereignty and treaty rights.
- Reclaim the anadromous fish resource and the environment on which it depends for future generations.
“When the Creator was preparing to bring humans onto the earth, He called a grand council of all the animal people, plant people, and everything else. In those days, the animals and plants were more like people because they could talk. He asked each one to give a gift to the humans—a gift to help them survive, since humans were pitiful and would die without help. The first to come forward was Salmon. He gave the humans his body for food. The second to give a gift was Water. She promised to be the home to the salmon. After that, everyone else gave the humans a gift, but it was special that the first to give their gifts were Salmon and Water. When the humans finally arrived, the Creator took away the animals’ power of speech and gave it to the humans. He told the humans that since the animals could no longer speak for themselves, it was a human responsibility to speak for the animals. To this day, Salmon and Water are always served first at tribal feasts to remember the story and honor the First Foods.”
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, R.N. Return to the River: Restoring Salmon to the Columbia River; Elsevier Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2006; pp. 1–720. [Google Scholar]
- Folke, C. Resilience (Republished). Ecol. Soc. 2016, 21, 44. [Google Scholar] [CrossRef]
- Gustafson, R.G.; Waples, R.S.; Myers, J.M.; Weitkamp, L.A.; Bryant, G.J.; Johnson, O.W.; Hard, J.J. Pacific salmon extinctions: Quantifying lost and remaining diversity. Conserv. Biol. 2007, 21, 1009–1020. [Google Scholar] [CrossRef]
- Dittmer, K. Changing streamflow on Columbia basin tribal lands—Climate change and salmon. Clim. Chang. 2013, 120, 627–641. [Google Scholar] [CrossRef]
- Wolf, E.C. Salmon Nation: People, Fish, and Our Common Home, 2nd ed.; Oregon State Univeristy Press: Corvallis, OR, USA, 2003; pp. 1–92. [Google Scholar]
- Folke, F.; Carpenter, C.; Walker, W.; Scheffer, M.; Chapin, T.; Rockström, J. Resilience thinking: Integrating resilience, adaptability and transformability. Ecol. Soc. 2010, 15, 20. [Google Scholar] [CrossRef]
- Baird, S. Salmon fisheries of Oregon. The Oregonian, 3 March 1875. [Google Scholar]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A. Resilience, adaptability and transformability in social–ecological systems. Ecol. Soc. 2004, 9, 5. Available online: http://www.ecologyandsociety.org/vol9/iss2/art5/ (accessed on 27 February 2023). [CrossRef]
- Westley, F.R.; Tjornbo, O.; Schultz, L.; Olsson, P.; Folke, C.; Crona, B.; Bodin, Ö. A theory of transformative agency in linked social-ecological systems. Ecol. Soc. 2013, 18, 27. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Folke, C.; Kofinas, G.P. (Eds.) A framework for understanding change. In Principles of Ecosystem Stewardship; Springer: New York, NY, USA, 2009; pp. 3–28. [Google Scholar]
- Sendzimir, S.; Magnuszewski, M.; Flachner, F.; Balogh, B.; Molnar, G.; Sarvari, A.; Nagy, Z. Assessing the resilience of a river management regime: Informal learning in a shadow network in the Tisza River Basin. Ecol. Soc. 2008, 13, 11. Available online: http://www.ecologyandsociety.org/vol13/iss1/art11/ (accessed on 27 February 2023). [CrossRef]
- Ludwig, D.; Jones, D.; Holling, S. Qualitative Analysis of Insect Outbreak Systems: The Spruce Budworm and Forest. J. Anim. Ecol. 1978, 47, 315–332. [Google Scholar] [CrossRef]
- Carpenter, C.; Walker, W.; Anderies, A.; Abel, A. From metaphor to measurement: Resilience of what to what? Ecosystems 2001, 4, 765–781. [Google Scholar] [CrossRef]
- Rinaldi, S.; Scheffer, M. Geometric analysis of ecological models with slow and fast processes. Ecosystems 2000, 3, 507–521. [Google Scholar] [CrossRef]
- Smith, D.M.S.; Abel, N.; Walker, B.; Chapin, F.S., III. Drylands: Coping with Uncertainty, Thresholds, and Changes in State. In Principles of Ecosystem Stewardship; Chapin, F.S., III, Folke, C., Kofinas, G.P., Eds.; Springer: New York, NY, USA, 2009; pp. 171–195. [Google Scholar]
- Folke, F.; Carpenter, C.; Elmqvist, E.; Gunderson, G.; Holling, H.; Walker, W. Resilience and sustainable development: Building adaptive capacity in a world of transformations. Ambio 2002, 31, 437–440. [Google Scholar] [CrossRef]
- Neville, H.M.; Isaak, D.J.; Dunham, J.B.; Thurow, F.R.; Rieman, B.E. Fine-scale natal homing and localized movement as shaped by sex and spawning habitat in Chinook salmon: Insights from spatial autocorrelation analysis of individual genotypes. Mol. Ecol. 2006, 15, 4589–4602. [Google Scholar] [CrossRef]
- Webb, W.; Bodin, B. A Network Perspective on Modularity and Control of Flow in Robust Systems. In Complexity Theory for a Sustainable Future; Norburg, N., Cumming, C., Eds.; Columbia Press: New York, NY, USA, 2008; pp. 85–118. [Google Scholar]
- Levin, S.A.; Lubchenco, J. Resilience, robustness, and marine ecosystem-based management. BioScience 2008, 58, 27–32. [Google Scholar] [CrossRef]
- Walker, B.; Salt, D. Resilience Thinking: Sustaining Ecosystems and People in a Changing World; Island Press: Washington, DC, USA, 2006; 192p. [Google Scholar]
- Walker, B.; Salt, D. Resilience Practice: Building Capacity to Absorb Disturbance and Maintain Function; Island Press: Washington, DC, USA, 2012; 248p. [Google Scholar]
- Biggs, R.; Schlüter, M.; Biggs, D.; Bohensky, E.L.; BurnSilver, S.; Cundill, G.; Dakos, V.; Daw, T.M.; Evans, L.S.; Kotschy, K.; et al. Toward principles for enhancing the resilience of ecosystem services. Annu. Rev. Environ. Resour. 2012, 37, 421–448. [Google Scholar] [CrossRef]
- Kharrazi, A.; Yu, Y.; Jacob, A.; Vora, N.; Fath, B.D. Redundancy, Diversity, and Modularity in Network Resilience: Applications for International Trade and Implications for Public Policy. Curr. Res. Environ. Sustain. 2020, 2, 100006. Available online: https://www.sciencedirect.com/science/article/pii/S2666049020300049?via%3Dihub (accessed on 9 March 2023). [CrossRef]
- Lichatowitch, J. Salmon Without Rivers: A History of the Pacific Salmon Crisis; Island Press: Washington, DC, USA, 2001; 336p. [Google Scholar]
- Wright, S. Evolution in Mendelian populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef]
- Wright, S. The roles of mutation, inbreeding, crossbreeding and selection in evolution. In Proceedings of the 6th International Congress of Genetics, Ithaca, NY, USA, 24–31 August 1932; pp. 356–366. Available online: http://www.esp.org/books/6th-congress/facsimile/contents/6th-cong-p356-wright.pdf (accessed on 27 February 2023).
- Wright, S.W. The shifting balance theory and macroevolution. Annu. Rev. Genet. 1982, 16, 1–19. [Google Scholar] [CrossRef]
- Kinzig, A.P.; Ryan, P.; Etienne, M.; Allison, H.; Elmqvist, T.; Walker, B.H. Resilience and regime shifts: Assessing cascading effects. Ecol. Soc. 2006, 11, 20. Available online: http://www.ecologyandsociety.org/vol11/iss1/art20/ (accessed on 27 February 2023). [CrossRef]
- Rietkerk, M.; Dekker, S.C.; de Ruiter, P.C.; van de Koppel, J. Self-organized patchiness and catastrophic shifts in ecosystems. Science 2004, 305, 1926–1929. [Google Scholar] [CrossRef]
- Erős, E.; Olden, O.; Schick, S.; Schmera, S.; Fortin, F. Characterizing connectivity relationships in freshwaters using patch-based graphs. Landsc. Ecol. 2012, 27, 303–317. [Google Scholar] [CrossRef]
- Campbell, G.C. Structural complexity, movement bias, and metapopulation extinction risk in dendritic ecological networks. J. N. Am. Benthol. Soc. 2011, 30, 252–258. [Google Scholar] [CrossRef]
- Yeakel, J.D.; Moore, J.W.; Guimarães, P.R.; Aguiar, M.A.M. Synchronisation and stability in river metapopulation networks. Ecol. Lett. 2014, 17, 273–283. [Google Scholar] [CrossRef]
- Mari, L.; Casagrandi, R.; Bertuzzo, E.; Rinaldo, A.; Gatto, M. Metapopulation persistence and species spread in river networks. Ecol. Lett. 2014, 17, 426–434. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/ele.12242 (accessed on 27 February 2023). [CrossRef]
- Ulanowicz, R.E.; Holt, R.D.; Barfield, M. Limits on ecosystem trophic complexity: Insights from ecological network analysis. Ecol. Lett. 2014, 17, 127–136. [Google Scholar] [CrossRef]
- Waples, R.S.; Pess, G.R.; Beechie, T. Evolutionary history of Pacific salmon in dynamic environments. Evol. Appl. 2008, 1, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Fagan, F. Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Letcher, B.H.; Schueller, P.; Bassar, R.D.; Nislow, K.H.; Coombs, J.A.; Sakrejda, K.; Morrissey, M.; Sigourney, D.B.; Whiteley, A.R.; O’Donnell, M.J.; et al. Robust estimates of environmental effects on population vital rates: An integrated capture–recapture model of seasonal brook trout growth, survival and movement in a stream network. J. Anim. Ecol. 2015, 84, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Gende, S.M.; Edwards, R.T.; Willson, M.F.; Wipfli, M.S. Pacific Salmon in Aquatic and Terrestrial Ecosystems. BioScience 2002, 52, 917–928. [Google Scholar] [CrossRef]
- Hocking, M.D.; Reimchen, T.E. Salmon-derived nitrogen in terrestrial invertebrates from coniferous forests of the Pacific Northwest. BMC Ecol. 2002, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Cederholm, C.J.; Kunze, M.D.; Murota, T.; Sibatani, A. Pacific salmon carcasses: Essential contributions of nutrients and energy for aquatic and terrestrial ecosystems. Fisheries 1999, 24, 6–15. [Google Scholar] [CrossRef]
- Gresh, T.; Lichatowich, J.; Schoonmaker, P. An estimation of historic and current levels of salmon production in the Northeast Pacific ecosystem: Evidence of a nutrient deficit in the freshwater systems of the Pacific Northwest. Fisheries 2000, 25, 15–21. [Google Scholar] [CrossRef]
- Willson, M.F.; Halupka, K.C. Anadromous fish as keystone species in vertebrate communities. Conserv. Biol. 1995, 9, 489–497. [Google Scholar] [CrossRef]
- Hocking, M.D.; Reynolds, J.D. Impacts of Salmon on Riparian Plant Diversity. Science 2011, 331, 1609–1612. [Google Scholar] [CrossRef]
- Spencer, C.N.; McClelland, B.R.; Stanford, J.A. Shrimp stocking, salmon collapse, and eagle displacement. BioScience 1991, 41, 14–21. [Google Scholar] [CrossRef]
- Brannon, E.; Powell, M.; Quinn, T.; Talbot, A. Population Structure of Columbia River Basin Chinook Salmon and Steelhead Trout; Project No. 1998-00403, BPA Report DOE/BP-08319-1; Bonneville Power Administration: Portland, OR, USA, 2002; pp. 1–181. [Google Scholar]
- Richter, A.; Kolmes, S.A. Maximum temperature limits for Chinook, coho, and chum salmon, and steelhead trout in the Pacific Northwest. Rev. Fish. Sci. 2005, 13, 23–49. [Google Scholar] [CrossRef]
- Wasowski, R.; Alexander, D.; Kolmes, S.; Banet, N.; Card, A.; Haines, M.; Kolmes, E.; Northcutt, K.; Roemer, D.; Webber, S. A multi-year longitudinal study of water quality parameters in four salmon-bearing and recreational streams on mount hood, Oregon. PLoS ONE 2013, 8, e70453. [Google Scholar] [CrossRef] [PubMed]
- McElhaney, P.; Backman, T.; Busak, C.; Heppell, S.; Kolmes, S.; Maule, A.; Myers, J.; Rawding, D.; Shively, D.; Steel, A.; et al. Interim Report on Viability Criteria for Willamette and Lower Columbia Basin Salmonids; National Marine Fisheries Service: Woods Hole, MA, USA; Northwest Fisheries Science Center: Seattle, WA, USA, 2003; pp. 1–331. [Google Scholar]
- Myers, J.; Busack, C.; Rawding, D.; Marshall, A.; Teel, D.; Van Doornik, D.M.; Maher, M.T. Historical Population Structure of Pacific Salmonids in the Willamette River and Lower Columbia River Basins; NMFS-NWFSC-73; U.S. Department of Commerce, NOAA Technical Memorandum: Washington, DC, USA, 2006; 311p. [Google Scholar]
- Gardner, C.A.; Scott, W.E.; Major, J.J.; Pierson, T.C. Mount Hood—History and Hazards of Oregon’s Most Recently Active Volcano; U.S. Geological Survey Fact Sheet, 2000. 060-00 Online Version 1.0. Available online: http://geopubs.wr.usgs.gov/fact-sheet/fs060-00/ (accessed on 27 February 2023).
- Waples, R.; Beechie, T.; Pess, G.R. Evolutionary History, Habitat Disturbance Regimes, and Anthropogenic Changes: What Do These Mean for Resilience of Pacific Salmon Populations? Ecol. Soc. 2009, 14, 3. Available online: http://www.ecologyandsociety.org/vol14/iss1/art3/ (accessed on 16 March 2023). [CrossRef]
- Butler, V.; O’Connor, J. 9000 years of salmon fishing on the Columbia River, North America. Quat. Res. 2004, 62, 1–8. [Google Scholar] [CrossRef]
- Hamlet, A.F.; Lettenmaier, D.P. Columbia River streamflow forecasting based on ENSO and PDO climate signals. J. Water Resour. Plan. Manag. 1999, 125, 333–341. [Google Scholar] [CrossRef]
- Taylor, J.E. Making Salmon: An Environmental History of the Northwest Fisheries Crisis; University of Washington Press: Seattle, WA, USA, 2001; pp. 1–488. [Google Scholar]
- O’Connor, O. The evolving landscape of the Columbia River Gorge: Lewis and Clark and cataclysms on the Columbia. Or. Hist. Soc. 2004, 105, 390–421. [Google Scholar] [CrossRef]
- Kramer, A.M.; Berec, L.; Drake, J.M. Allee effects in ecology and evolution. J. Anim. Ecol. 2018, 87, 7–10. Available online: https://besjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2656.12777 (accessed on 8 March 2023). [CrossRef] [PubMed]
- Greene, G.; Hall, H.; Guilbault, G.; Quinn, Q. Improved viability of populations with diverse life-history portfolios. Biol. Lett. 2010, 6, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Lichatowitch, J. Salmon, People, and Place: A Biologist’s Search for Salmon Recovery; Oregon State University Press: Corvallis, OR, USA, 2013; pp. 1–272. [Google Scholar]
- Miller, R.R. Is the Past Present? Historical Splash-Dam Mapping and Stream Disturbance Detection in the Oregon Coastal Province. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2010; pp. 1–110. Available online: https://ir.library.oregonstate.edu/concern/parent/4m90f001t/file_sets/dj52w8373 (accessed on 2 March 2023).
- Atlas, W.I.; Ban, N.C.; Moore, J.W.; Tuohy, A.M.; Greening, S.; Reid, A.J.; Morven, N.; White, E.; Housty, W.G.; Housty, J.A.; et al. Indigenous Systems of Management for Culturally and Ecologically Resilient Pacific Salmon (Oncorhynchus spp.) Fisheries. BioScience 2021, 71, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Cosens, B.; Fremier, A. Assessing System Resilience and Ecosystem Services in Large River Basins: A Case Study of the Columbia River Basin. Ida. Law Rev. 2014, 51, 91–125. [Google Scholar]
- Middleton, B.A. (Ed.) The flood pulse concept in wetland restoration. In Flood Pulsing in Wetlands: Restoring the Natural Hydrological Balance; John Wiley & Sons: New York, NY, USA, 2002; pp. 1–10. [Google Scholar]
- Hulse, D.; Gregory, S.; Baker, J. Willamette River Basin Atlas; Oregon State University Press: Corvallis, OR, USA, 2002; pp. 1–178. Available online: https://oregonexplorer.info/content/willamette-river-basin-atlas?topic&ptopic (accessed on 3 March 2023).
- Rohe, R. The Chinese and Hydraulic Mining in the Far West. Min. Hist. Assoc. Annu. 1994, 1, 73–91. Available online: https://www.mininghistoryassociation.org/Journal/MHJ-v1-1994-Rohe.pdf (accessed on 8 March 2023).
- Kareiva, P.; Marvier, M.; Silliman, B. (Eds.) Uncomfortable questions and inconvenient data in conservation science. In Effective Conservation Science: Data Not Dogma; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Tierney, K.B.; Sampson, J.L.; Ross, P.S.; Sekela, M.A.; Kennedy, C.J. Salmon Olfaction is Impaired by an Environmentally Realistic Pesticide Mixture. Environ. Sci. Technol. 2008, 42, 4996–5001. [Google Scholar] [CrossRef]
- Parman, D.L. Inconstant Advocacy: The Erosion of Indian Fishing Rights in the Pacific Northwest, 1933–1956. Pac. Hist. Rev. 1984, 53, 163–189. [Google Scholar] [CrossRef]
- University of Washington Libraries Digital Collection. Salmon in the Pacific Northwest and Alaska Collection, 1890–1961; University of Washington Libraries Digital Collection: Seattle, WA, USA, 2001; Available online: https://content.lib.washington.edu/salmonweb/index.html (accessed on 7 March 2023).
- Mighetto, L.; Ebel, W.J. Saving the Salmon: A History of the U.S. Army Corps of Engineers’ Efforts to Protect Anadromous Fish on the Columbia and Snake Rivers; Historical Research Associates, The Division: Missoula, MT, USA, 1994; pp. 1–253. Available online: https://usace.contentdm.oclc.org/digital/collection/p16021coll4/id/96/ (accessed on 7 March 2023).
- Report of the Commissioner—United States Commission of Fish and Fisheries . 1895, pp. 1–1979. Available online: https://archive.org/stream/reportcommissio06statgoog/reportcommissio06statgoog_djvu.txt (accessed on 8 March 2023).
- White, R. The Organic Machine: The Remaking of the Columbia River; Hill and Wang Critical Issues/MacMillan: New York, NY, USA, 1995; pp. 1–130. [Google Scholar]
- Terui, A.; Urabe, H.; Senzaki, H.; Nishizawa, B. Intentional release of native species undermines ecological stability. Proc. Natl. Acad. Sci. USA 2023, 120, e2218044120. [Google Scholar] [CrossRef] [PubMed]
- Kolmes, S.A. Salmon Farms and Hatcheries. Environ. Sci. Policy Sustain. Dev. 2004, 46, 40–43. [Google Scholar]
- Northwest Power and Conservation Council. Dams: History and Purpose. Available online: https://www.nwcouncil.org/reports/columbia-river-history/damshistory/ (accessed on 8 March 2023).
- Fish Commission of the State of Oregon. Biennial Report of the Fish Commission of the State of Oregon. 1931, pp. 1–33. Available online: https://ir.library.oregonstate.edu/concern/technical_reports/r207tt794 (accessed on 8 March 2023).
- Lang, W.T. 1949: Year of Decision on the Columbia River. Columbia Mag. 2005, 19, 8–15. Available online: https://www.washingtonhistory.org/wp-content/uploads/2020/04/1949-1.pdf (accessed on 8 March 2023).
- Caudill, C.C.; Keefer, M.L.; Clabough, T.S.; Naughton, G.P.; Burke, B.J.; Peery, C.A. Indirect Effects of Impoundment on Migrating Fish: Temperature Gradients in Fish Ladders Slow Dam Passage by Adult Chinook Salmon and Steelhead. PLoS ONE 2013, 8, e85586. [Google Scholar] [CrossRef] [PubMed]
- Whitney, W.; Coutant, C.; Mundy, M.; Williams, W. Mitigation of salmon losses due to hydroelectric development. In Return to the River, Restoring Salmon to the Columbia River; Williams, R.N., Ed.; Elsevier Academic Press: New York, NY, USA, 2006; pp. 325–416. [Google Scholar]
- Northwest Power and Conservation Council. Fish Passage at Dams. Available online: https://www.nwcouncil.org/reports/columbia-river-history/fishpassage/ (accessed on 8 March 2023).
- Sheer, M.B.; Steel, E.A. Lost Watersheds: Barriers, Aquatic Habitat Connectivity, and Salmon Persistence in the Willamette and Lower Columbia River Basins. Trans. Am. Fish. Soc. 2006, 135, 1654–1669. Available online: https://www.fs.usda.gov/pnw/lwm/aem/docs/steel/2006_sheer_and_steel_lost_watersheds.pdf (accessed on 8 March 2023). [CrossRef]
- Washington Department of Fisheries. The Salmon Crisis. 1951, pp. 1–14. Available online: https://digitalcollections.lib.washington.edu/digital/collection/salmon/id/421 (accessed on 9 March 2023).
- Tomczak, M.T.; Heymans, J.J.; Yletyinen, J.; Niiranen, S.; Otto, S.A.; Blenckner, T. Ecological network indicators of ecosystem status and change in the Baltic Sea. PLoS ONE 2013, 8, e75439. [Google Scholar] [CrossRef] [PubMed]
- Brock, W.A.; Carpenter, S.R. Early Warnings of Regime Shift When the Ecosystem Structure Is Unknown. PLoS ONE 2012, 7, e45586. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Bascompte, J.; Brock, W.A.; Brovkin, V.; Carpenter, S.R.; Dakos, V.; Held, H.; van Nes, E.H.; Rietkerk, M.; Sugihara, G. Early-warning signals for critical transitions. Nature 2009, 461, 53–59. [Google Scholar] [CrossRef]
- Dakos, V.; Carpenter, S.R.; Brock, W.A.; Ellison, A.M.; Guttal, V.; Ives, A.R.; Kéfi, S.; Livina, V.; Seekell, D.A.; van Nes, E.H.; et al. Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data. PLoS ONE 2012, 7, e41010. [Google Scholar] [CrossRef]
- Biggs, R.; Carpenter, S.R.; Brock, W.A. Turning back from the brink: Detecting an impending regime shift in time to avert it. Proc. Natl. Acad. Sci. USA 2009, 106, 826–831. [Google Scholar] [CrossRef]
- Goodwell, A.; Campbell, N. An information theory-based approach to characterize drivers of upstream salmon migration. PLoS ONE 2022, 17, e0269193. [Google Scholar] [CrossRef]
- Lackey, R.T.; Lach, D.H.; Duncan, S.L. (Eds.) Chapter 3. Wild Salmon in Western North America: Forecasting the Most Likely Status in 2100. In Salmon 2100: The Future of Wild Pacific Salmon; American Fisheries Society: Bethesda, MD, USA, 2006; pp. 57–70. [Google Scholar]
- Lackey, R.T. Saving wild salmon: A 165 year policy conundrum. In Proceedings of the Dubach Workshop: Science and Scientists in the Contemporary Policy Process, Oregon State University, Portland, OR, USA, 3–4 October 2013; Available online: http://forestpolicypub.com/wp-content/uploads/2014/01/165-Year-Salmon-Policy-Conundrum-R-T-Lackey.pdf (accessed on 16 March 2022).
- Biggs, R.; Peterson, G.D.; Rocha, J.C. The Regime Shifts Database: A framework for analyzing regime shifts in social-ecological systems. Ecol. Soc. 2018, 23, 9. [Google Scholar] [CrossRef]
- Newberry, D. The Organic Machine: A Discussion of Human Relationships with Nature. Built Environment Plus. 2017. Available online: https://builtenvironmentplus.org/organic-machine-discussion-human-relationships-nature/ (accessed on 7 March 2023).
- Geiling, N. How Oregon’s Second Largest City Vanished in a Day. Smithsonian Magazine. 2015. Available online: https://www.smithsonianmag.com/history/vanport-oregon-how-countrys-largest-housing-project-vanished-day-180954040/ (accessed on 7 March 2023).
- Blumm, M.C.; Thorson, E.J.; Smith, J.D. Practiced at the Art of Deception: The Failure of Columbia Basin Salmon Recovery Under the Endangered Species Act. Environ. Law 2006, 36, 709–810. [Google Scholar]
- Blumm, M.C.; Erickson, A. Dam Removal in the Pacific Northwest: Lessons for the Nation. Environ. Law 2012, 42, 1043–1100. [Google Scholar] [CrossRef]
- Federal Energy Regulatory Commission. Order Modifying and Approving Surrender of License and Rrmoval of Project Facilities, 181 FERC 61,122. 2022. Available online: https://www.ferc.gov/media/h-1-p-2082-063 (accessed on 7 March 2023).
- Cohen, B.R. Escaping the false binary of nature and culture through connection: Richard White’s The Organic Machine: The Remaking of the Columbia River. Organ. Environ. 2005, 18, 445–457. [Google Scholar] [CrossRef]
- Lackey, R.T.; Lach, D.H.; Duncan, S.L. (Eds.) The Challenge of Restring Wild Salmon. In Salmon 2100, The Future of Wild Pacific Salmon; American Fisheries Society: Bethesda, MD, USA, 2006; pp. 1–11. [Google Scholar]
- Mantua, N.J.; Hare, S.R. The Pacific Decadal Oscillation. J. Oceanogr. 2002, 58, 35–44. [Google Scholar] [CrossRef]
- Di Lorenzo, E.; Schneider, N.; Cobb, K.M.; Franks PJ, S.; Chhak, K.; Miller, A.J.; McWilliams, J.C.; Bograd, S.J.; Arango, H.; Curchitser, E.; et al. North Pacific Gyre Oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 2008, 35, L08607. [Google Scholar] [CrossRef]
- Tootle, G.A.; Piechota, T.C.; Singh, A. Coupled oceanic-atmospheric vatiability and US streamflow. Water Resour. Res. 2005, 41, 1–11. [Google Scholar] [CrossRef]
- Bottom, D.L.; Jones, K.K.; Simenstad, C.A.; Smith, C.L. Reconnecting social and ecological resilience in salmon ecosystems. Ecol. Soc. 2009, 14, 5. Available online: http://www.ecologyandsociety.org/vol14/iss1/art5/ (accessed on 7 March 2023). [CrossRef]
- Ford, P. Declining Salmon Counts on Salmon River Flashing Red Light Warning Extinction. The Columbia Basin Bulletin, Fish & Wildlife News. 2022. Available online: https://cbbulletin.com/guest-column-declining-salmon-counts-on-salmon-river-flashing-red-light-warning-extinction/ (accessed on 7 March 2023).
- Herbold, B.; Carlson, S.M.; Henery, R.; Johnson, R.C.; Mantua, N.; McClure, M.; Moyle, P.; Sommer, T. Managing for Salmon Resilience in California’s Variable and Changing Climate. San Fr. Estuary Watershed Sci. 2018, 16, 3. [Google Scholar] [CrossRef]
- Mason, J.G.; Eurich, J.G.; Lau, J.D.; Battista, W.; Free, C.M.; Mills, K.E.; Tokunaga, K.; Zhao, L.Z.; Dickey-Collas, M.; Valle, M.; et al. Attributes of climate resilience in fisheries: From theory to practice. Fish Fish. 2022, 23, 522–544. [Google Scholar] [CrossRef]
- Hartman, G.F.; Groot, C.; Northcote, T.G. Science and Management in Sustainable Salmonid Fisheries: The Ball is Not in Our Court. In Sustainable Fisheries Management, Pacific Salmon; Knudsen, E.E., McDonald, D., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 1–752. [Google Scholar]
- Lackey, R.T. The great Pacific Northwest Salmon conundrum. Water Resour. IMPACT (Am. Water Resour. Assoc.) 2022, 24, 5–8. [Google Scholar]
- Northwest Power and Conservation Council. Guide for Subbasin Planners. 2001. Council Document Number: 2001-20. Available online: https://www.nwcouncil.org/reports/technical-guide-subbasin-planners/ (accessed on 7 March 2023).
- Hill, G.; Kolmes, S.A.; Humphreys, M.; McLain, R.; Jones, E.T. Using decision support tools in multi-stakeholder environmental planning: Restorative justice and subbasin planning in the Columbia River Basin. J. Environ. Stud. Sci. 2019, 9, 170–186. [Google Scholar] [CrossRef]
- Hill, G.; Monticino, M.; Jones, E.T.; Kolmes, S.; McLain, R. Aspirational Goals and Incremental Tools: Does forecasting exclude other frameworks for Strategic planning? In Proceedings of the UC Berkeley: Computer Professionals for Social Responsibility, Directions and Implications of Advanced Computing Conference Proceedings, Berkeley, CA, USA, 26–29 June 2008; Available online: http://www.publicsphereproject.org/events/diac08/proceedings/index.html (accessed on 7 March 2023).
- Rahr, R.; Augerot, A. A proactive sanctuary strategy to anchor and restore high-priority wild salmon ecosystems. In Salmon 2100, The Future of Wild Pacific Salmon; Lackey, R.T., Lach, D.H., Duncan, S.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 2006; pp. 465–489. [Google Scholar]
- Schloss, C.A.; Lawler, J.J.; Larson, E.R.; Papendick, H.L.; Case, M.J.; Evans, D.M.; DeLap, J.H.; Langdon, J.G.R.; Hall, S.A.; McRae, B.H. Systematic Conservation Planning in the Face of Climate Change: Bet-Hedging on the Columbia Plateau. PLoS ONE 2011, 6, e28788. [Google Scholar] [CrossRef] [PubMed]
- Senate Hearing 111-833, S. 817. The Pacific Salmon Stronghold Conservation Act. The U.S. Government Publishing Office. pp. 1–94. Available online: https://www.govinfo.gov/content/pkg/CHRG-111shrg64140/html/CHRG-111shrg64140.htm (accessed on 7 March 2023).
- “Salmon Focused Investments in Sustainable Habitats Act” Is Available at Congressman Huffman’s Website. Available online: https://huffman.house.gov/imo/media/doc/Huffman%20Salmon%20FISH%20Act%20117th%20FINAL.pdf (accessed on 7 March 2023).
- Columbia River Inter-Tribal Fish Commission. Spirit of the Salmon: Wy-Kan-Ush-Mi Wa-Kish-Wit. Available online: https://plan.critfc.org/ (accessed on 14 March 2013).
- Columbia Basin Partnership Task Force (CBPTF). A Vision for Salmon and Steelhead: Goals to Restore Thriving Salmon and Steelhead to the Columbia River Basin: Phase 2 Report of the Columbia Basin Partnership Task Force of the Marine Fisheries Advisory Committee. October 2020. Available online: https://www.fisheries.noaa.gov/vision-salmon-and-steelhead-goals-restore-thriving-salmon-and-steelhead-columbia-river-basin (accessed on 16 March 2023).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, G.M.; Kolmes, S.A. A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies. Environments 2023, 10, 76. https://doi.org/10.3390/environments10050076
Hill GM, Kolmes SA. A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies. Environments. 2023; 10(5):76. https://doi.org/10.3390/environments10050076
Chicago/Turabian StyleHill, Gregory M., and Steven A. Kolmes. 2023. "A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies" Environments 10, no. 5: 76. https://doi.org/10.3390/environments10050076
APA StyleHill, G. M., & Kolmes, S. A. (2023). A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies. Environments, 10(5), 76. https://doi.org/10.3390/environments10050076