Pine Stands as Bioindicators: Justification for Air Toxicity Monitoring in an Industrial Metropolis




Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment for the Permanent Sample Plots in the Pine Stands
2.2. Acute and Chronic Impacts on Pine Stands
2.3. Phytopathological Control of Needles in the SPs
2.4. The Stem Phloem Defense Response (Stem Lesion Test)
2.5. Preparation of the Fungal Extract
3. Results and Discussion
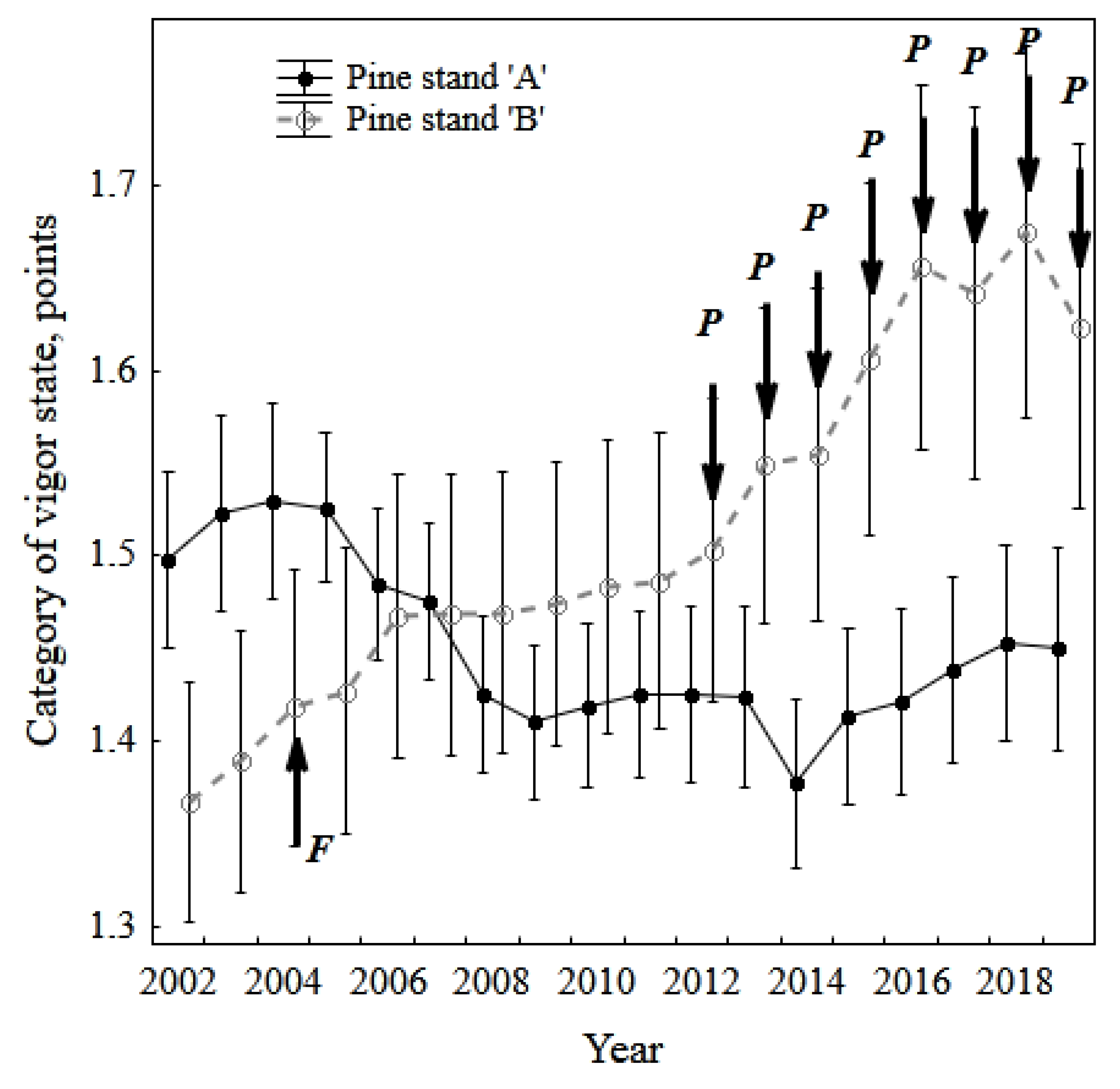
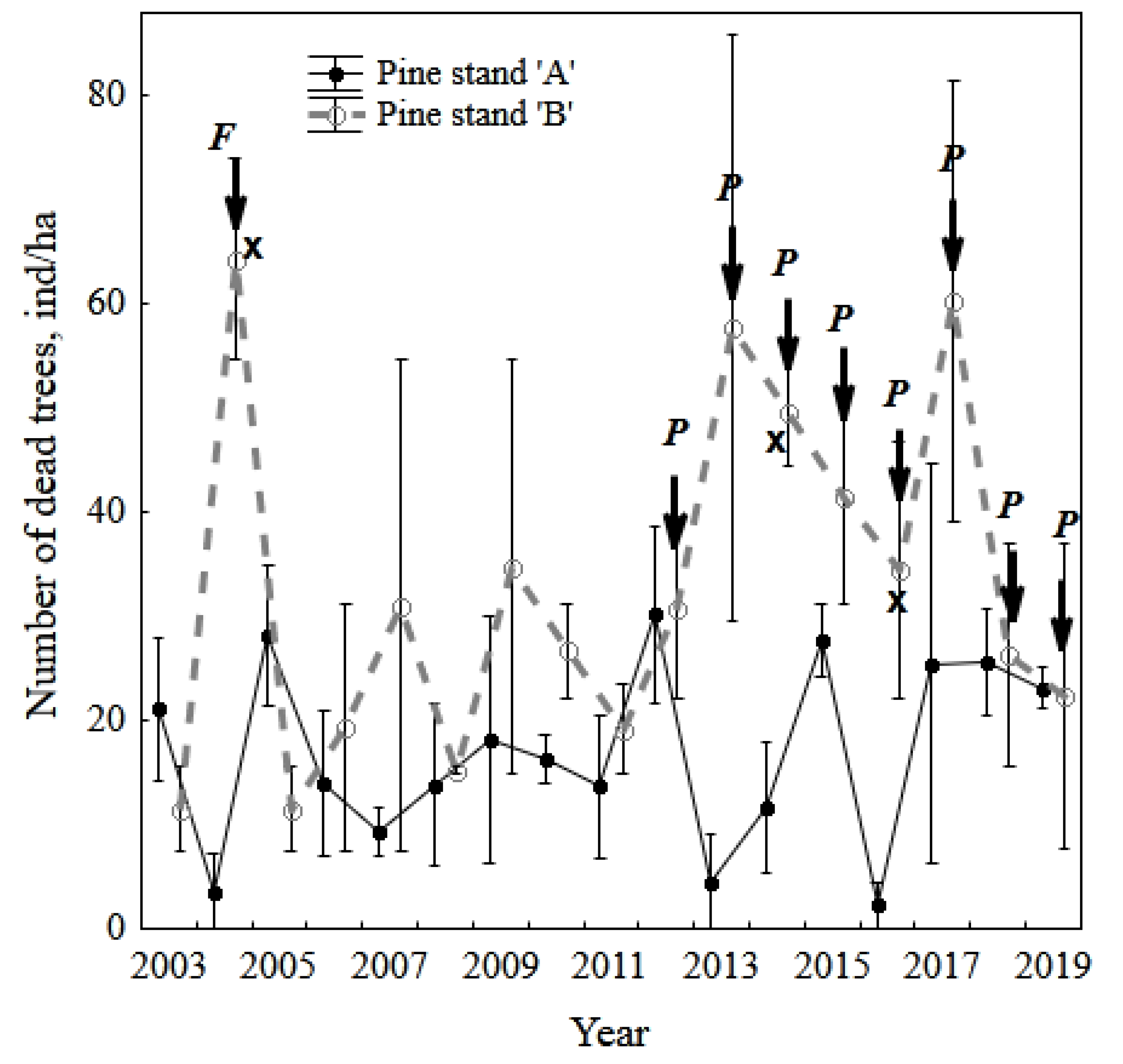
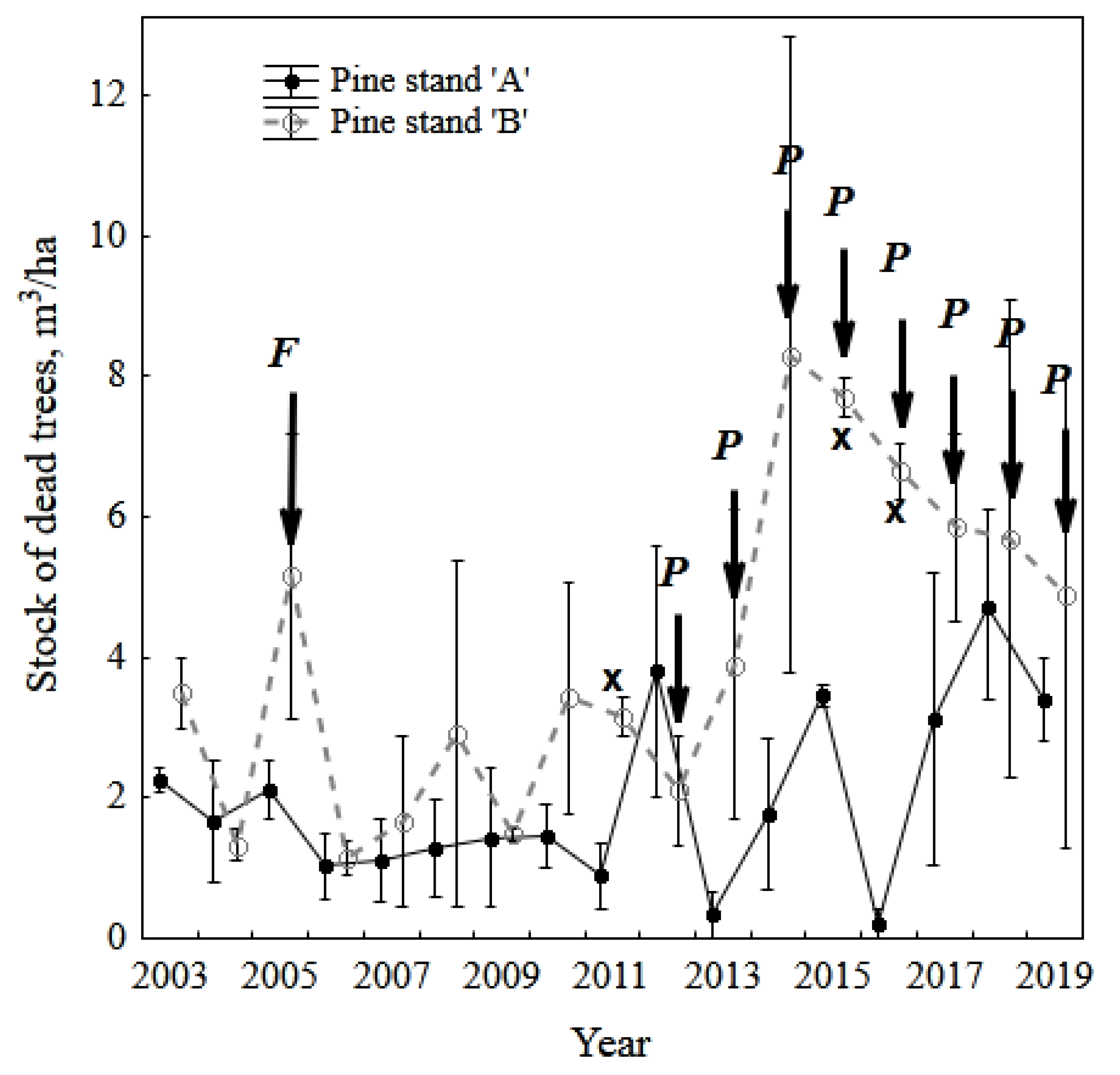
3.1. Vigor Rank of Pine Stands, Needle Necrosis, and Fall (Death) of Trees
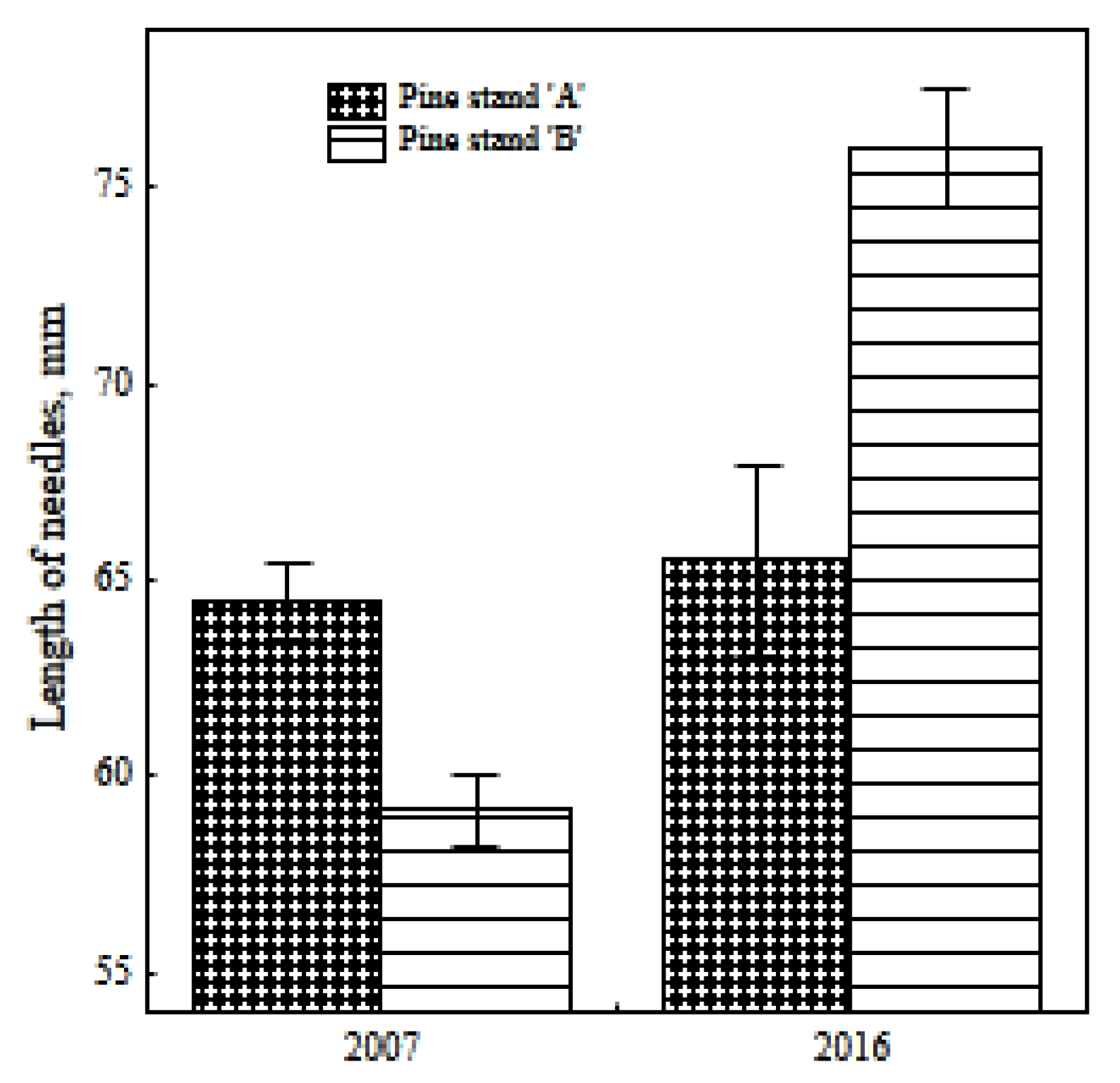
3.2. Needle Length and Selye’s Adaptation Syndrome (Hormesis)
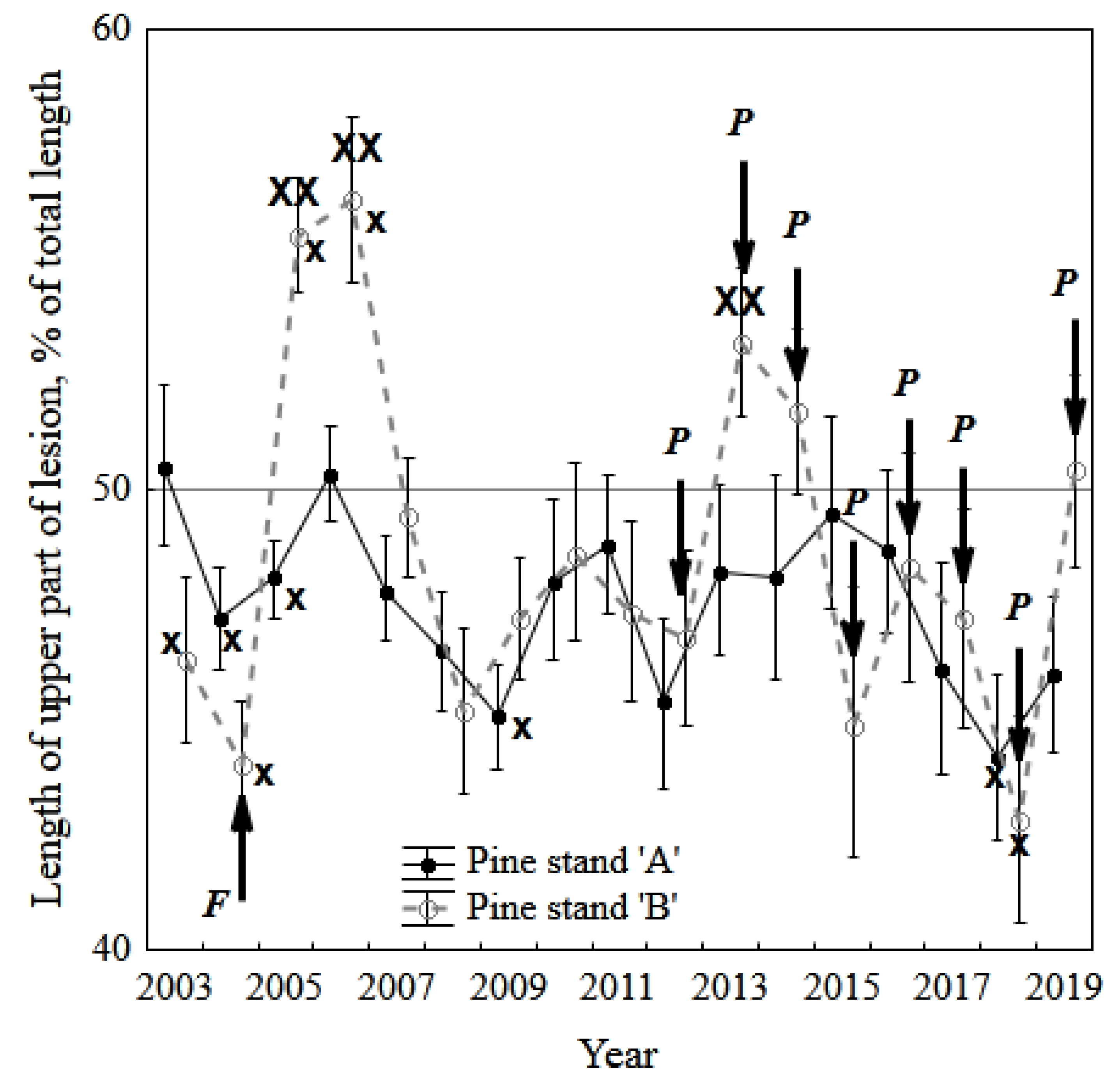
3.3. The Stem Lesion Test (Inoculation of the Trunk with the Fungal Extract and Registration of Stem Phloem Necrosis)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huo, D.; Wu, J.; Kong, Q.; Zhang, G.B.; Wang, Y.Y.; Yang, H.Y. Macromolecular toxins secreted by botrytis cinerea induce programmed cell death in arabidopsis leaves. Russ. J. Plant Physiol. 2018, 65, 579–587. [Google Scholar] [CrossRef]
- Kudryasheva, N.S.; Kovel, E. Monitoring of low-intensity exposures via luminescent bioassays of different complexity: Cells, enzyme reactions, and fluorescent proteins. Int. J. Mol. Sci. 2019, 20, 4451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewska, M.; Klimek, B. Trace element concentrations in tree leaves and lichen collected along a metal pollution gradient near olkusz (Southern Poland). Bull. Environ. Contam. Toxicol. 2017, 100, 245–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, P.; Fox, S.; Mickler, R.A. Impacts of Air Pollutants on Southern Pine Forests. J. Appl. Ecol. 1997, 34, 264. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Kandziora-Ciupa, M.; Ciepał, R.; Barczyk, G. Robinia pseudoacacia and Melandrium album in trace elements biomonitoring and air pollution tolerance index study. Int. J. Environ. Sci. Technol. 2016, 13, 1741–1752. [Google Scholar] [CrossRef] [Green Version]
- Mengel, K.; Breininger, M.T.; Lutz, H.J. Effect of acidic mist on nutrient leaching, carbohydrate status and damage symptoms of picea abies. In Air Pollution and Ecosystems; Springer Science and Business Media LLC: Berlin, Germany, 1988; pp. 312–320. [Google Scholar]
- Danek, M. The influence of industry on scots pine stands in the south-eastern part of the silesia-krakow upland (Poland) on the basis of dendrochronological analysis. Water Air Soil Pollut. 2007, 185, 265–277. [Google Scholar] [CrossRef]
- Polyakova, G.G.; Pashenova, N.V.; Polyakov, V.I.; Zrazhevskaya, G.K. Induction of conifer immune responses by phytopathogenic fungus metabolites. Russ. J. Plant Physiol. 2008, 55, 496–502. [Google Scholar] [CrossRef]
- Polyakova, G.G.; Stasova, V.V.; Pashenova, N.V. Defense response of pine stem phloem to wounding and treatment with mycelial extracts from Ceratocystis laricicola. Russ. J. Plant Physiol. 2011, 58, 819–827. [Google Scholar] [CrossRef]
- Polyakova, G.G.; Pashenova, N.V.; Senashova, V.A. Influence of chronical partial defoliation of larch by casebearer on stem physiological parameters. Biol. Bull. 2020, 1, 44–50. [Google Scholar]
- Sanderson, E.G.; Farant, J.-P. Indoor and outdoor polycyclic aromatic hydrocarbons in residences surrounding a söderberg aluminum smelter in Canada. Environ. Sci. Technol. 2004, 38, 5350–5356. [Google Scholar] [CrossRef]
- Otnyukova, T.N. Medicinal kupena (Poligonatum odoratum)—An indicator of atmospheric pollution by fluorine. Vestnik KrasGAU 2013, 4, 111–117. (In Russian) [Google Scholar]
- Ranking of World Air Quality Index (AQI) and PM2.5 Air Pollution. Available online: https://www.ncbi.nlm.nih.gov/pubmed/15543736 (accessed on 17 February 2020).
- Directions for Forest Management of the Russian Forest Resources; Voenizdat: Moscow, Russia, 1995. (In Russian)
- Forest Inventory Regulations of the Russian Federation; Ministry of Natural Resources: Moscow, Russia, 2008. (In Russian)
- Polyakov, V.I.; Polyakova, G.G.; Stasova, V.V. Application of fungal metabolites for evaluation of pine stans vigor in conditions of industrial pollution. Biol. Bull. 2005, 32, 419–424. [Google Scholar] [CrossRef]
- Cherednikova, Y.S.; Molokova, N.I.; Krasnoshchekov, Y.N.; Perevoznikova, V.D. Zoning and typological diversity of forests in Krasnoyarsk green belt. Geogr. Nat. Resour. 1999, 3, 84–91. (In Russian) [Google Scholar]
- Tret’yakov, N.V.; Gorskii, P.V.; Samoilovich, G.G. Spravochnik Taksatora (The Inventory’s Handbook); Goslesbumizdat: Leningrad, Russia, 1952. (In Russian) [Google Scholar]
- Sanitary Regulations in Forests of the Russian Federation (Forest Regulation of the Russian Federation. Collection of Regulatory Documents); PAIMS: Moscow, Russia, 1998; pp. 310–329. (In Russian)
- Nemich, V.N. Section area sums of normal pine stands in Priangarie. For. Invent. Manag. 1996, 1, 110–114. (In Russian) [Google Scholar]
- Maksimov, S.V. Potential Phytomass Productivity of Scots Pine Cultures and Its Geography (On the Example of North Eurasia). Ph.D. Thesis, Ekaterinburg University, Ekaterinburg, Russia, 2000. (In Russian). [Google Scholar]
- Semenkova, I.G.; Sokolova, E.S. Forest Phytopathology; Ekologiya Publishers: Moscow, Russia, 1992. (In Russian) [Google Scholar]
- Reid, R.W.; Whitney, H.S.; Watson, J.A. Reactions of lodgepole pine to attack by dendroctonus ponderosae Hopkins and blue stain fungi. Can. J. Bot. 1967, 45, 1115–1126. [Google Scholar] [CrossRef]
- Vetrova, V.P.; Polyakova, G.G.; Matrenina, R.M.; Pashenova, N.V. Accumulation of condensed tannins and lignin in fir and larh phloem induced by blye-stain fungi and their elicitors. Bark Beetles, Blue-stain Fungi, and Conifer Defence Systems. In Proceedings of the North American Forest Insect Work Conference, Ankara, Turkey, 31 July–2 August 1995; pp. 18–19. [Google Scholar]
- Solheim, H. Ecological aspects of fungi associated with the spruce bark beetle. In Norway Ceratocystis and Ophiostoma, 2nd ed.; Wingfild, M.J., Seifert, K.A., Webber, J.F., Eds.; APS Press: St. Paul, MN, USA, 1999; pp. 235–242. [Google Scholar]
- Goodsman, D.W.; Lusebrink, I.; Landhäusser, S.M.; Erbilgin, N.; Lieffers, V. Variation in carbon availability, defense chemistry and susceptibility to fungal invasion along the stems of mature trees. New Phytol. 2012, 197, 586–594. [Google Scholar] [CrossRef]
- Kutaf’eva, N.P. Morhpology of Mushrooms; Sib. Univ. Publ. House: Novosibirsk, Russia, 2003. (In Russian) [Google Scholar]
- International Union of Forest Research Organizations. Internationally Dangerous Forest Tree Diseases; International Union of Forest Research Organizations: Vienna, Austria, 1963. [Google Scholar]
- Selye, H. Changing distress into eustress: Hans Selye voices theories on stress. Tex. Med. 1980, 76, 78–100. [Google Scholar]
- Pakhomova, V.M. The basic principles of modern theory of stress and the nonspecific adaptive syndrome of plants. Russ. J. Cytol. 1995, 37, 66–91. (In Russian) [Google Scholar]
- Burlakova, E.B.; Konradov, A.A.; Maltseva, E.L. Effects of extremely weak chemical and physical stimuli on biological systems. Biophysics (Moscow) 2004, 49, 522–534. [Google Scholar]
- Wang, C.-R.; Tian, Y.; Wang, X.; Yu, H.-X.; Lu, X.-W.; Wang, C.; Wang, H. Hormesis effects and implicative application in assessment of lead-contaminated soils in roots of Vicia faba seedlings. Chemosphere 2010, 80, 965–971. [Google Scholar] [CrossRef]
- Kudryasheva, N.; Rozhko, T. Effect of low-dose ionizing radiation on luminous marine bacteria: Radiation hormesis and toxicity. J. Environ. Radioact. 2015, 142, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Shi, J.; Huber, M.; Wang, T.; Dali, W.; Yin, C.-S. Progress in the studies on hormesis of low-dose pollutants. Environ. Dis. 2016, 1, 58. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis: Path and progression to significance. Int. J. Mol. Sci. 2018, 19, 2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eschrich, W. Bidirectional Transport. In Encyclopedia of Plant Physiology; Zimmermann, M.H., Milburn, J.A., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2018; Volume 3, pp. 245–255. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SP No. | Year | Stand Composition | Degree of Density 1 | Principal Forest Element | Mean | Stock, m3/ha | Depth 3, ind/ha | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Age, Years | Height, m | Dia- Meter, cm 2 | Green Timber | Dead- Standing Trees | Green Timber | Dead- Standing Trees | |||||
| 1 | 2002 | 10PII+PI 4 | 1.4 | PII | 57 | 18.1 | 18.5 | 396 | 6 | 1671 | 56 |
| 1 | 2019 | 10PII+PI | 1.6 | » | 74 | 21.7 | 22.6 | 550 | 17 | 1360 | 119 |
| 2 | 2002 | 9PII+PI rB | 1.5 | PII | 58 | 21.3 | 21.9 | 489 | 10 | 1286 | 36 |
| 2 | 2019 | 10PII+PI | 1.6 | » | 75 | 25.4 | 26.3 | 658 | 16 | 1043 | 86 |
| 3 | 2002 | 9P1B r.L | 1.5 | P | 62 | 21.1 | 20.4 | 472 | 8 | 1430 | 81 |
| 3 | 2019 | 9P1B r.L | 1.6 | » | 80 | 25.2 | 25.2 | 616 | 29 | 1067 | 163 |
| 4 | 2002 | 9P1L+B | 1.5 | P | 62 | 20.9 | 19.6 | 503 | 9 | 1664 | 172 |
| 4 | 2019 | 9P1L+B | 1.6 | » | 81 | 25.5 | 25.5 | 628 | 32 | 1055 | 227 |
| 5 | 2005 | 9PII1PI | 1.5 | PII | 59 | 16.3 | 16.4 | 339 | 3 | 1980 | 41 |
| 5 | 2019 | 9PII1PI | 1.6 | » | 73 | 18.7 | 19.0 | 433 | 10 | 1682 | 88 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polyakova, G.; Pashenova, N.; Senashova, V.; Podolyak, N.; Kudryasheva, N. Pine Stands as Bioindicators: Justification for Air Toxicity Monitoring in an Industrial Metropolis. Environments 2020, 7, 28. https://doi.org/10.3390/environments7040028
Polyakova G, Pashenova N, Senashova V, Podolyak N, Kudryasheva N. Pine Stands as Bioindicators: Justification for Air Toxicity Monitoring in an Industrial Metropolis. Environments. 2020; 7(4):28. https://doi.org/10.3390/environments7040028
Chicago/Turabian StylePolyakova, Galina, Natalia Pashenova, Vera Senashova, Natalia Podolyak, and Nadezhda Kudryasheva. 2020. "Pine Stands as Bioindicators: Justification for Air Toxicity Monitoring in an Industrial Metropolis" Environments 7, no. 4: 28. https://doi.org/10.3390/environments7040028
APA StylePolyakova, G., Pashenova, N., Senashova, V., Podolyak, N., & Kudryasheva, N. (2020). Pine Stands as Bioindicators: Justification for Air Toxicity Monitoring in an Industrial Metropolis. Environments, 7(4), 28. https://doi.org/10.3390/environments7040028